
生物多样性 ›› 2016, Vol. 24 ›› Issue (1): 30-39. DOI: 10.17520/biods.2015207 cstr: 32101.14.biods.2015207
郭屹立1,2, 王斌1,2, 向悟生1,2, 丁涛1,2, 陆树华1,2, 黄甫昭1,2, 文淑均1,2, 李冬兴1,2, 何运林1,2, 李先琨1,2,*( )
)
收稿日期:2015-07-18
接受日期:2015-11-02
出版日期:2016-01-20
发布日期:2016-06-12
通讯作者:
李先琨
基金资助:
Yili Guo1,2, Bin Wang1,2, Wusheng Xiang1,2, Tao Ding1,2, Shuhua Lu1,2, Fuzhao Huang1,2, Shujun Wen1,2, Dongxing Li1,2, Yunlin He1,2, Xiankun Li1,2,*()
Received:2015-07-18
Accepted:2015-11-02
Online:2016-01-20
Published:2016-06-12
Contact:
Li Xiankun
摘要:
森林生态系统地上生物量的分布格局是物种分布格局的重要内容。局域尺度内因地形差异引起的生境异质性是地上生物量分布格局形成的环境基础。本文以弄岗北热带喀斯特季节性雨林15 ha动态监测样地2011年第一次普查数据中每木个体为研究对象, 尝试以每20 m × 20 m样方内所有个体及不同径级类群的胸高断面积之和为木本植物地上生物量的衡量指标, 利用广义可加模型对喀斯特季节性雨林中胸高断面积之和的空间分布格局进行研究, 定量分析了其对7个地形因子的响应。结果表明, 洼地、山坡和山脊3种生境类型中, 所有个体的胸高断面积之和山坡最高、山脊最低, 且山脊与山坡、洼地的差异均显著; 广义可加模型结果显示, 不同地形因子对胸高断面积之和的解释偏差差异明显, 其中海拔、坡向、凹凸度、岩石裸露率对胸高断面积之和的解释偏差依次降低, 而干旱度指数、坡度和地形湿润指数解释偏差相对较小。喀斯特季节性雨林木本植物胸高断面积之和空间分布的异质性及其与地形因子之间的关系, 反映了胸高断面积之和在地形因子对土壤、水分和光照等条件重分配影响下的多重响应机制及生长策略。
郭屹立, 王斌, 向悟生, 丁涛, 陆树华, 黄甫昭, 文淑均, 李冬兴, 何运林, 李先琨 (2016) 喀斯特季节性雨林木本植物胸高断面积分布格局及其对地形因子的响应. 生物多样性, 24, 30-39. DOI: 10.17520/biods.2015207.
Yili Guo, Bin Wang, Wusheng Xiang, Tao Ding, Shuhua Lu, Fuzhao Huang, Shujun Wen, Dongxing Li, Yunlin He, Xiankun Li (2016) Responses of spatial pattern of woody plants’ basal area to topographic factors in a tropical karst seasonal rainforest in Nonggang, Guangxi, southern China. Biodiversity Science, 24, 30-39. DOI: 10.17520/biods.2015207.
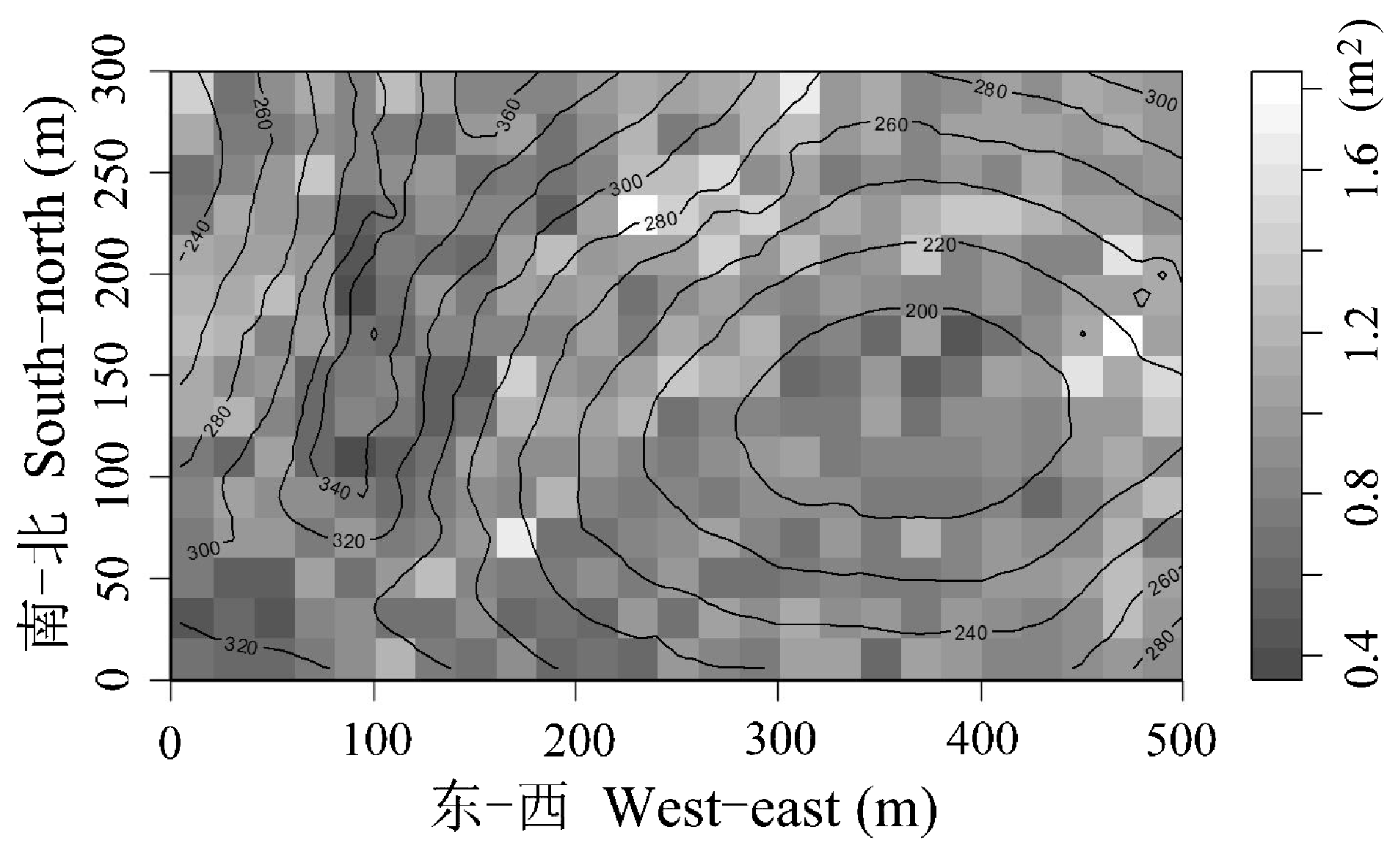
图1 弄岗喀斯特季节性雨林胸高断面积之和(m2)的分布格局
Fig. 1 Spatial patterns of total basal area of karst seasonal rainforest in Nonggang, Guangxi
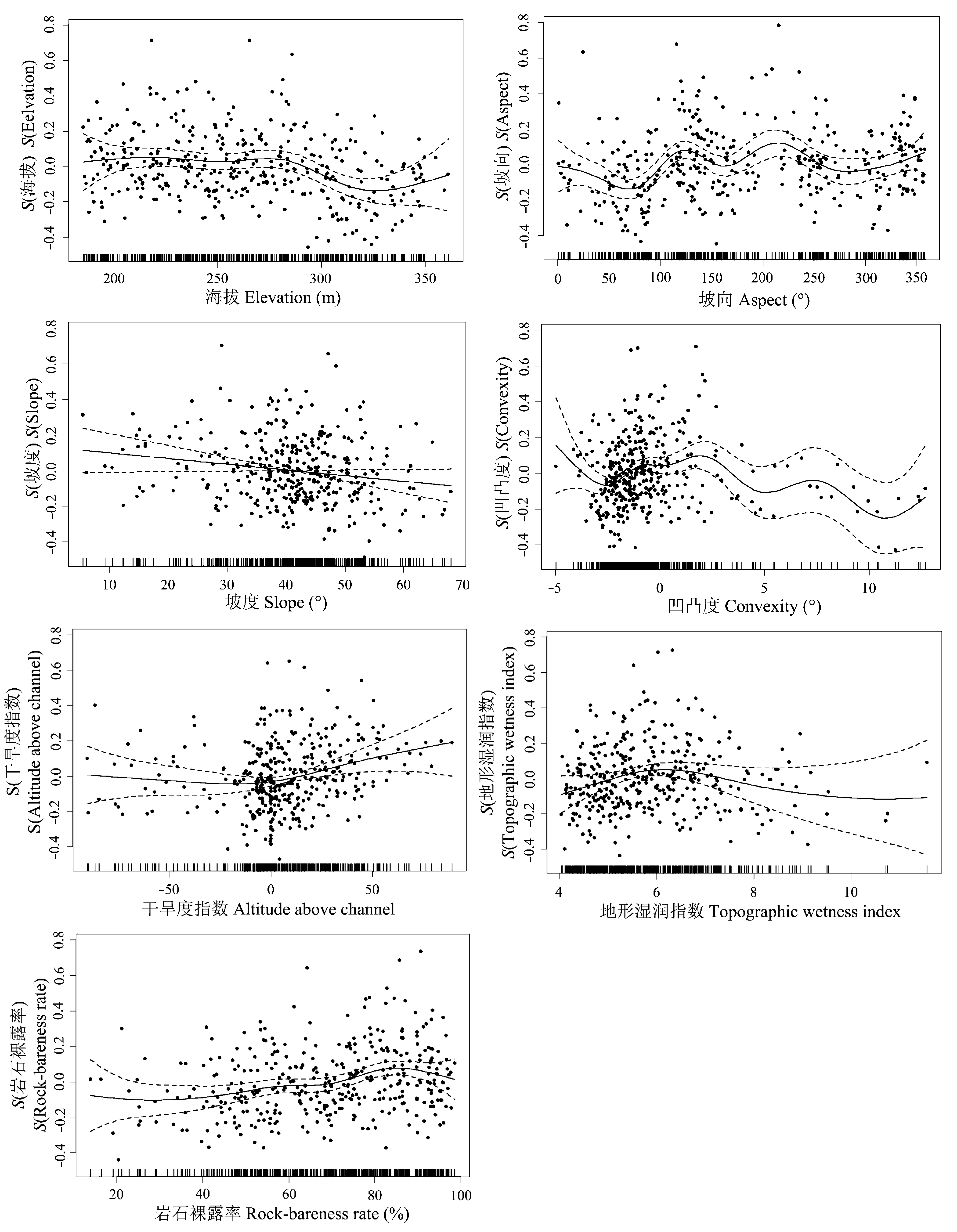
图2 广义可加模型(GAM)所揭示的不同地形因子对弄岗喀斯特季节性雨林胸高断面积之和分布格局的影响。S(地形因子)为光滑样条函数的拟合值, 表示其对胸高断面积之和的影响。实线表示胸高断面积之和的期望值, 上下两侧虚线表示方程的95%置信区间。
Fig. 2 Results of generalized additive models (GAM) regression between different topographic factors and total basal area of the karst seasonal rainforest in Nonggang, Guangxi. S(topographic factor) is the fitted value of smoothing spline functions, which represent their impacts on the total basal area. The solid lines represent the expected values of total basal area; the dotted lines represent the 95% confidence intervals of equations.
| 径级 DBH Class (cm) | 环境参数 Environmental parameters | r2adj | 累计解释偏差 Cumulative explained deviation (%) | 赤池信息准则Akaike Information Criterion (AIC) |
|---|---|---|---|---|
| 所有个体 All individuals | 海拔 Elevation | 0.148 | 16.1 | -88.071 |
| 坡向 Aspect | 0.256 | 28.4 | -130.984 | |
| 凹凸度 Convexity | 0.291 | 33.1 | -142.770 | |
| 岩石裸露率 Rock-bareness rate | 0.313 | 36.0 | -148.860 | |
| 地形湿润指数Topographic wetness index | 0.314 | 36.4 | -149.909 | |
| 坡度 Slope | 0.322 | 37.4 | -152.044 | |
| 干旱度指数 Altitude above channel | 0.336 | 39.4 | -156.461 | |
| DBH < 2.5 | 海拔 Elevation | 0.113 | 12.6 | -2,416.026 |
| 坡向 Aspect | 0.153 | 18.4 | -2,425.371 | |
| 凹凸度 Convexity | 0.193 | 22.8 | -2,441.038 | |
| 岩石裸露率 Rock-bareness rate | 0.266 | 29.7 | -2,477.055 | |
| 地形湿润指数Topographic wetness index | 0.303 | 34.0 | -2,492.105 | |
| 坡度 Slope | 0.311 | 34.9 | -2,495.940 | |
| 干旱度指数 Altitude above channel | 0.328 | 36.3 | -2,506.965 | |
| 2.5 ≤ DBH < 7.5 | 海拔 Elevation | 0.412 | 42.1 | -1,340.583 |
| 坡向 Aspect | 0.432 | 44.7 | -1,348.138 | |
| 凹凸度 Convexity | 0.462 | 48.9 | -1,360.737 | |
| 岩石裸露率 Rock-bareness rate | 0.481 | 51.1 | -1,371.831 | |
| 地形湿润指数Topographic wetness index | 0.505 | 53.1 | -1,390.343 | |
| 坡度 Slope | 0.540 | 57.6 | -1,407.893 | |
| 干旱度指数 Altitude above channel | 0.540 | 57.8 | -1,410.145 | |
| 7.5 ≤ DBH < 22.5 | 海拔 Elevation | 0.194 | 21.0 | -448.036 |
| 坡向 Aspect | 0.267 | 29.5 | -476.660 | |
| 凹凸度 Convexity | 0.305 | 34.4 | -490.249 | |
| 岩石裸露率 Rock-bareness rate | 0.316 | 36.2 | -492.008 | |
| 地形湿润指数Topographic wetness index | 0.334 | 38.0 | -498.367 | |
| 坡度 Slope | 0.339 | 38.5 | -507.223 | |
| 干旱度指数 Altitude above channel | 0.381 | 43.6 | -522.046 | |
| 22.5 ≤ DBH | 海拔 Elevation | 0.263 | 27.4 | -193.325 |
| 坡向 Aspect | 0.321 | 34.7 | -216.574 | |
| 凹凸度 Convexity | 0.326 | 35.4 | -216.982 | |
| 岩石裸露率 Rock-bareness rate | 0.335 | 36.5 | -220.844 | |
| 地形湿润指数Topographic wetness index | 0.345 | 37.8 | -225.257 | |
| 坡度 Slope | 0.392 | 42.8 | -250.109 | |
| 干旱度指数 Altitude above channel | 0.398 | 43.7 | -251.184 |
表1 喀斯特季节性雨林胸高断面积之和与地形因子的广义可加模型(GAM)检验
Table 1 Tests of generalized additive models (GAM) for modeling total basal area in the Nonggang karst seasonal rainforest and topographic factors
| 径级 DBH Class (cm) | 环境参数 Environmental parameters | r2adj | 累计解释偏差 Cumulative explained deviation (%) | 赤池信息准则Akaike Information Criterion (AIC) |
|---|---|---|---|---|
| 所有个体 All individuals | 海拔 Elevation | 0.148 | 16.1 | -88.071 |
| 坡向 Aspect | 0.256 | 28.4 | -130.984 | |
| 凹凸度 Convexity | 0.291 | 33.1 | -142.770 | |
| 岩石裸露率 Rock-bareness rate | 0.313 | 36.0 | -148.860 | |
| 地形湿润指数Topographic wetness index | 0.314 | 36.4 | -149.909 | |
| 坡度 Slope | 0.322 | 37.4 | -152.044 | |
| 干旱度指数 Altitude above channel | 0.336 | 39.4 | -156.461 | |
| DBH < 2.5 | 海拔 Elevation | 0.113 | 12.6 | -2,416.026 |
| 坡向 Aspect | 0.153 | 18.4 | -2,425.371 | |
| 凹凸度 Convexity | 0.193 | 22.8 | -2,441.038 | |
| 岩石裸露率 Rock-bareness rate | 0.266 | 29.7 | -2,477.055 | |
| 地形湿润指数Topographic wetness index | 0.303 | 34.0 | -2,492.105 | |
| 坡度 Slope | 0.311 | 34.9 | -2,495.940 | |
| 干旱度指数 Altitude above channel | 0.328 | 36.3 | -2,506.965 | |
| 2.5 ≤ DBH < 7.5 | 海拔 Elevation | 0.412 | 42.1 | -1,340.583 |
| 坡向 Aspect | 0.432 | 44.7 | -1,348.138 | |
| 凹凸度 Convexity | 0.462 | 48.9 | -1,360.737 | |
| 岩石裸露率 Rock-bareness rate | 0.481 | 51.1 | -1,371.831 | |
| 地形湿润指数Topographic wetness index | 0.505 | 53.1 | -1,390.343 | |
| 坡度 Slope | 0.540 | 57.6 | -1,407.893 | |
| 干旱度指数 Altitude above channel | 0.540 | 57.8 | -1,410.145 | |
| 7.5 ≤ DBH < 22.5 | 海拔 Elevation | 0.194 | 21.0 | -448.036 |
| 坡向 Aspect | 0.267 | 29.5 | -476.660 | |
| 凹凸度 Convexity | 0.305 | 34.4 | -490.249 | |
| 岩石裸露率 Rock-bareness rate | 0.316 | 36.2 | -492.008 | |
| 地形湿润指数Topographic wetness index | 0.334 | 38.0 | -498.367 | |
| 坡度 Slope | 0.339 | 38.5 | -507.223 | |
| 干旱度指数 Altitude above channel | 0.381 | 43.6 | -522.046 | |
| 22.5 ≤ DBH | 海拔 Elevation | 0.263 | 27.4 | -193.325 |
| 坡向 Aspect | 0.321 | 34.7 | -216.574 | |
| 凹凸度 Convexity | 0.326 | 35.4 | -216.982 | |
| 岩石裸露率 Rock-bareness rate | 0.335 | 36.5 | -220.844 | |
| 地形湿润指数Topographic wetness index | 0.345 | 37.8 | -225.257 | |
| 坡度 Slope | 0.392 | 42.8 | -250.109 | |
| 干旱度指数 Altitude above channel | 0.398 | 43.7 | -251.184 |
| 坡度 Slope (SLO) | 凹凸度 Convexity (CON) | 坡向 Aspect (ASP) | 地形湿润指数Topographic wetness index (TWI) | 干旱度指数 Altitude above channel (ACH) | 岩石裸露率 Rock-bareness rate (RBR) | 胸高断面积之和 Total basal area | |
|---|---|---|---|---|---|---|---|
| 海拔 Elevation (ELE) | 0.578** | 0.466** | 0.052 | -0.701** | 0.289** | 0.450** | -0.147* |
| SLO | 0.336** | 0.205* | -0.631** | 0.220** | 0.509** | -0.080 | |
| CON | 0.025 | -0.564** | 0.518** | 0.300** | 0.051 | ||
| ASP | -0.098 | -0.159* | 0.140* | 0.134* | |||
| TWI | -0.629** | -0.326** | 0.035 | ||||
| ACH | -0.016 | 0.083 | |||||
| RBR | 0.044 |
表2 不同地形因子间的Spearman秩相关性分析
Table 2 The Spearman correlations (rho values) between different topographic factors
| 坡度 Slope (SLO) | 凹凸度 Convexity (CON) | 坡向 Aspect (ASP) | 地形湿润指数Topographic wetness index (TWI) | 干旱度指数 Altitude above channel (ACH) | 岩石裸露率 Rock-bareness rate (RBR) | 胸高断面积之和 Total basal area | |
|---|---|---|---|---|---|---|---|
| 海拔 Elevation (ELE) | 0.578** | 0.466** | 0.052 | -0.701** | 0.289** | 0.450** | -0.147* |
| SLO | 0.336** | 0.205* | -0.631** | 0.220** | 0.509** | -0.080 | |
| CON | 0.025 | -0.564** | 0.518** | 0.300** | 0.051 | ||
| ASP | -0.098 | -0.159* | 0.140* | 0.134* | |||
| TWI | -0.629** | -0.326** | 0.035 | ||||
| ACH | -0.016 | 0.083 | |||||
| RBR | 0.044 |
| 1 | Alves LF, Vieira SA, Scaranello MA, Camargo PB, Santos FAM, Joly CA, Martinelli LA (2010) Forest structure and live aboveground biomass variation along an elevational gradient of tropical Atlantic moist forest (Brazil). Forest Ecology and Management, 260, 679-691. |
| 2 | Burnham KP, Anderson DR (2002) Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd edn. Springer, New York. |
| 3 | Cantón Y, Del Barrio G, Solé-Benet A, Lázaro R (2004) Topographic controls on the spatial distribution of ground cover in the Tabernas badlands of SE Spain. Catena, 55, 341-365. |
| 4 | Chauvel A, Lucas Y, Boulet R (1987) On the genesis of the soil mantle of the region of Manaus, Central Amazonia. Brazil Experientia, 43, 234-241. |
| 5 | Clements R, Sodhi N S, Schilthuizen M, Ng PK (2006) Limestone karsts of Southeast Asia: imperiled arks of biodiversity. BioScience, 56, 733-742. |
| 6 | Condit R (1998) Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and a Comparison with Other Plots. Springer, Berlin. |
| 7 | Deng JM, Qin BQ, Wang BW (2015) Quick implementing of generalized additive models using R and its application in bluegreen algal bloom forecasting. Chinese Journal of Ecology, 34, 835-842. (in Chinese with English abstract) |
| [邓建明, 秦伯强, 王博雯 (2015) 广义可加模型在R中的快捷实现及蓝藻水华预测分析中的应用. 生态学杂志, 34, 835-842.] | |
| 8 | Deng ZQ (1988) Report on the investigation of karst geology from Nonggang Natural Reserve. Guihaia, (S1), 1-16. (in Chinese) |
| [邓自强 (1988) 广西弄岗自然保护区综合考察报告. 广西植物, S1, 1-16.] | |
| 9 | Dong X, Bennion H, Maberly SC, Sayer CD, Simpson GL, Battarbee RW (2012) Nutrients exert a stronger control than climate on recent diatom communities in Esthwaite Water: evidence from monitoring and palaeolimnological records. Freshwater Biology, 57, 2044-2056. |
| 10 | Engelbrecht BMJ, Kursar TA, Tyree MT (2005) Drought effects on seedling survival in a tropical moist forest. Trees: Structure and Function, 19, 312-321. |
| 11 | Guisan A, Edwards Jr TC, Hastie T (2002) Generalized linear and generalized additive models in studies of species distributions: setting the scene. Ecological Modelling, 157, 89-100. |
| 12 | Guo FT, Hu HQ, Jin S, Ma ZH, Zhang Y (2010) Relationship between forest lighting fire occurrence and weather factors in Daxing’an Mountains based on negative binomial model and zero-inflated negative binomial models. Chinese Journal of Plant Ecology, 34, 571-577. (in Chinese with English abstract) |
| [郭福涛, 胡海清, 金森, 马志海, 张扬 (2010) 基于负二项和零膨胀负二项回归模型的大兴安岭地区雷击火与气象因素的关系. 植物生态学报, 34, 571-577.] | |
| 13 | Guo YL, Wang B, Xiang WS, Ding T, Lu SH, Huang FZ, Li DX, Wen SJ, He YL, Li XK (2015a) Sprouting characteristics of tree species in 15 ha northern tropical karst seasonal rain forest dynamics plot in Nonggang, Guangxi, southern China. Chinese Journal of Ecology, 34, 955-961. (in Chinese with English abstract) |
| [郭屹立, 王斌, 向悟生, 丁涛, 陆树华, 黄甫昭, 李冬兴, 文淑均, 何运林, 李先琨 (2015a) 弄岗北热带喀斯特季节性雨林15 ha样地萌生特征分析. 生态学杂志, 34, 955-961.] | |
| 14 | Guo YL, Wang B, Xiang WS, Ding T, Lu SH, Huang FZ, Li DX, Wen SJ, He YL, Li XK (2015b) Dynamics of density-dependent effects of tree species in a 15 ha seasonal rain forest plot in northern tropical karst in Nonggang, Guangxi, southern China. Chinese Science Bulltin, 60, 1602-1611. (in Chinese with English abstract) |
| [郭屹立, 王斌, 向悟生, 丁涛, 陆树华, 黄甫昭, 李冬兴, 文淑均, 何运林, 李先琨 (2015b) 弄岗喀斯特季节性雨林15 ha样地密度制约效应分析. 科学通报, 60, 1602-1611.] | |
| 15 | Harms KE, Condit R, Hubbell SP, Foster RB (2001) Habitat associations of trees and shrubs in a 50-ha neotropical forest plot. Journal of Ecology, 89, 947-959. |
| 16 | Hastie TJ, Tibshirani RJ (1990) Generalized Additive Models. CRC Press, Boca Raton. |
| 17 | Huang FZ, Wang B, Ding T, Xiang WS, Li XK, Zhou AP (2014) Numerical classification of associations in a northern tropical karst seasonal rain forest and the relationships of these associations with environmental factors. Biodiversity Science, 22, 157-166. (in Chinese with English abstract) |
| [黄甫昭, 王斌, 丁涛, 向悟生, 李先琨, 周爱萍 (2014) 弄岗北热带喀斯特季节性雨林群丛数量分类及与环境的关系. 生物多样性, 22, 157-166.] | |
| 18 | Jiang ZZ, Yuan DX (1999) Dynamics features of the epikarst zone and their significance in environment sand resources. Acta Geoscientica Sinica, 20, 302-308. (in Chinese with English abstract) |
| [蒋忠诚, 袁道先 (1999) 表层岩溶带的岩溶动力学特征及其环境和资源意义. 地球学报, 20, 302-308.] | |
| 19 | John R, Dalling JW, Harms KE, Yavitt JB, Stallard RF, Mirabello M, Hubbell SP, Valencia R, Navarrete H, Vallejo M, Foster RB (2007) Soil nutrients influence spatial distributions of tropical tree species. Proceedings of the National Academy of Sciences, USA, 104, 864-869. |
| 20 | Kanagaraj R, Wiegand T, Comita LS, Huth A (2011) Tropical tree species assemblages in topographical habitats change in time and with life stage. Journal of Ecology, 99, 1441-1452. |
| 21 | Lai JS, Mi XC, Ren HB, Ma KP (2009) Species-habitat associations change in a subtropical forest of China. Journal of Vegetation Science, 20, 415-423. |
| 22 | Li XK, Su ZM, Lü SH, Ou ZL, Xiang WS, Qu Z, Lu SH (2003) The spatial pattern of natural vegetation in the karst regions of Guangxi and the ecological signality for ecosystem rehabilitation and reconstruction. Journal of Mountain Science, 21, 129-139. (in Chinese with English abstract) |
| [李先琨, 苏宗明, 吕仕洪, 欧祖兰, 向悟生, 区智, 陆树华 (2003) 广西岩溶植被自然分布规律及对岩溶生态恢复重建的意义. 山地学报, 21, 129-139.] | |
| 23 | Li YB, Hou JJ, Xie DT (2002) The recent development of research on karst system of Southwest China. Scientia Geographica Sinica, 22, 365-370. (in Chinese with English abstract) |
| [李阳兵, 侯建筠, 谢德体 (2002) 中国西南岩溶生态研究进展. 地理科学, 22, 365-370.] | |
| 24 | Lin DM, Lai JS, Muller-Landau HC, Mi XC, Ma KP (2012) Topographic variation in aboveground biomass in a subtropical evergreen broad-leaved forest in China. PLoS ONE, 7, e48244. |
| 25 | Liu HF, Xue DY, Sang WG (2012) Effect of topographic factors on the relationship between species richness and aboveground biomass in a warm temperate forest. Ecology and Environmental Sciences, 21, 1403-1407. |
| [刘海丰, 薛达元, 桑卫国 (2012) 地形因子对暖温带森林群落物种丰富度-地上生物量关系的影响. 生态环境学报, 21, 1403-1407.] | |
| 26 | Liu XL, Shi ZM, Yang DS, Liu SR, Yang YP, Ma QY (2005) Advances in study on changes of biodiversity and productivity along elevational gradient in mountainous plant community. World Forestry Research, 18(4), 27-34. (in Chinese with English abstract) |
| [刘兴良, 史作民, 杨冬生, 刘世荣, 杨玉坡, 马钦彦 (2005) 山地植物群落生物多样性与生物生产力海拔梯度变化研究进展. 世界林业研究, 18(4), 27-34.] | |
| 27 | Luizão RCC, Luizão FJ, Paiva RQ, Monteiro TF, Sousa LS, Kruijt B (2004) Variation of carbon and nitrogen cycling processes along a topographic gradient in a central Amazonian forest. Global Change Biology, 10, 592-600. |
| 28 | Marshall AR, Willcock S, Platts PJ, Lovetta JC, Balmfordd A, Burgessd ND, Lathama JE, Munishih PKT, Saltera R, Shirimah DD, Lewisc SL (2012) Measuring and modelling aboveground carbon and tree allometry along a tropical elevation gradient. Biological Conservation, 154, 20-33. |
| 29 | Mascaro J, Asner G P, Muller-Landau H C, van Breugel M, Hall J, Dahlin K (2011) Controls over aboveground forest carbon density on Barro Colorado Island, Panama. Biogeosciences, 8, 1615-1629. |
| 30 | McEwan RW, Lin YC, Sun IF, Hsieh CF, Su SH, Chang LW, Song GZM, Wang HH, Hwong JL, Lin KC, Yang KC, Chiang JM (2011) Topographic and biotic regulation of aboveground carbon storage in subtropical broad-leaved forests of Taiwan. Forest Ecology and Management, 262, 1817-1825. |
| 31 | McEwan RW, Muller RN (2006) Spatial and temporal dynamics in canopy dominance of an old-growth central Appalachian forest. Canadian Journal of Forest Research, 36, 1536-1550. |
| 32 | Punchi-Manage R, Getzin S, Wiegand T, Kanagaraj R, Gunatilleke CVS, Gunatilleke IAUN, Wiegand K, Huth A (2013) Effects of topography on structuring local species assemblages in a Sri Lankan mixed dipterocarp forest. Journal of Ecology, 101, 149-160. |
| 33 | R Core Team (2014) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. |
| 34 | Sun HL (2005) Ecosystem of China. Science Press, Beijing. (in Chinese) |
| [孙鸿烈 (2005) 中国生态系统. 科学出版社, 北京.] | |
| 35 | Sutherland WJ, Aveling R, Bennun L, Chapman E, Clout M, Côté IM, Depledge MH, Dicks LV, Dobson AP, Fellman L, Fleishman E, Gibbons DW, Keim B, Lickorish F, Lindenmayer DB, Monk5 KA, Norris K, Peck LS, Prior SV, Scharlemann JPW, Spalding M, Watkinson AR (2012) A horizon scan of global conservation issues for 2012. Trends in Ecology and Evolution, 27, 12-18. |
| 36 | Swartzman G, Huang CH, Kaluzny S (1992) Spatial analysis of Bering Sea groundfish survey data using generalized additive models. Canadian Journal of Fisheries and Aquatic Sciences, 49, 1366-1378. |
| 37 | Takyu M, Aiba SI, Kitayama K (2003) Changes in biomass, productivity and decomposition along topographical gradients under different geological conditions in tropical lower montane forests on Mount Kinabalu, Borneo. Oecologia, 134, 397-404. |
| 38 | Tarboton DG (1997) A new method for the determination of flow directions and upslope areas in grid digital elevation models. Water Resources Research, 33, 309-319. |
| 39 | Valencia R, Foster RB, Villa G, Condit R, Svenning JC, Hernández C, Romoleroux K, Losos E, Magård E, Balslev H (2004) Tree species distributions and local habitat variation in the Amazon: large forest plot in eastern Ecuador. Journal of Ecology, 92, 214-229. |
| 40 | Wang B, Huang YS, Li XK, Xiang WS, Ding T, Huang FZ, Lu SH, Han WH, Wen SJ, He LJ (2014) Species composition and spatial distribution of a 15 ha northern tropical karst seasonal rain forest dynamics study plot in Nonggang, Guangxi, southern China. Biodiversity Science, 22, 141-156. (in Chinese with English abstract) |
| [王斌, 黄俞淞, 李先琨, 向悟生, 丁涛, 黄甫昭, 陆树华, 韩文衡, 文淑均, 何兰军 (2014) 弄岗北热带喀斯特季节性雨林15 ha监测样地的树种组成与空间分布. 生物多样性, 22, 141-156.] | |
| 41 | Wood SN (2001) mgcv: GAMs and generalized ridge regression for R. R News, 1, 20-25. |
| 42 | Wood SN (2004) Stable and efficient multiple smoothing parameter estimation for generalized additive models. Journal of the American Statistical Association, 99, 673-689. |
| 43 | Wood SN (2006a) Low rank scale invariant tensor product smooths for generalized additive mixed models. Biometrics, 62, 1025-1036. |
| 44 | Wood SN (2006b) Generalized Additive Models: An Introduction with R. Chapman and Hall/CRC Press, London. |
| [1] | 李艳朋, 陈洁, 卢春洋, 许涵. 海南尖峰岭热带山地雨林64 ha次生林动态监测样地群落结构特征[J]. 生物多样性, 2025, 33(2): 24445-. |
| [2] | 王鹏, 隋佳容, 丁欣瑶, 王伟中, 曹雪倩, 赵海鹏, 王彦平. 郑州城市公园鸟类群落嵌套分布格局及其影响因素[J]. 生物多样性, 2024, 32(3): 23359-. |
| [3] | 曹可欣, 王敬雯, 郑国, 武鹏峰, 李英滨, 崔淑艳. 降水格局改变及氮沉降对北方典型草原土壤线虫多样性的影响[J]. 生物多样性, 2024, 32(3): 23491-. |
| [4] | 林迪, 陈双林, 杜榷, 宋文龙, 饶固, 闫淑珍. 大别山黏菌的物种多样性调查[J]. 生物多样性, 2024, 32(2): 23242-. |
| [5] | 孟敬慈, 王国栋, 曹光兰, 胡楠林, 赵美玲, 赵延彤, 薛振山, 刘波, 朴文华, 姜明. 中国芦苇沼泽植物物种丰富度分布格局及其驱动因素[J]. 生物多样性, 2024, 32(2): 23194-. |
| [6] | 陈明苗, 张楚然, 邓云, 李生发, 李逢昌, 唐志忠, 魏兆喆, 张彩彩, 林露湘. 地形因子对亚热带半湿润常绿阔叶林木本植物萌生特征的影响[J]. 生物多样性, 2024, 32(12): 24282-. |
| [7] | 王丽媛, 胡慧建, 姜杰, 胡一鸣. 南岭哺乳类和鸟类物种丰富度空间分布格局及其影响因子[J]. 生物多样性, 2024, 32(1): 23026-. |
| [8] | 王明慧, 陈昭铨, 李帅锋, 黄小波, 郎学东, 胡子涵, 尚瑞广, 刘万德. 云南普洱季风常绿阔叶林不同种子扩散方式的优势种空间点格局分析[J]. 生物多样性, 2023, 31(9): 23147-. |
| [9] | 刘志发, 王新财, 龚粤宁, 陈道剑, 张强. 基于红外相机监测的广东南岭国家级自然保护区鸟兽多样性及其垂直分布特征[J]. 生物多样性, 2023, 31(8): 22689-. |
| [10] | 曹亚苏, 范敏, 彭羽, 辛嘉讯, 彭楠一. 景观格局动态对浑善达克沙地植物物种多样性和功能多样性的影响[J]. 生物多样性, 2023, 31(8): 23048-. |
| [11] | 杨俊毅, 关潇, 李俊生, 刘晶晶, 郝颢晶, 王槐睿. 乌江流域生物多样性与生态系统服务的空间格局及相互关系[J]. 生物多样性, 2023, 31(7): 23061-. |
| [12] | 杨胜娴, 杨清, 李晓东, 巢欣, 刘惠秋, 魏蓝若雪, 巴桑. 确定性过程主导高原典型河流浮游植物地理分布格局和群落构建[J]. 生物多样性, 2023, 31(7): 23092-. |
| [13] | 李世东. 中国和美国国家公园时空发展及驱动因素[J]. 生物多样性, 2023, 31(6): 23040-. |
| [14] | 陈宏, 冼晓青, 陈宜雪, 林娜, 王苗苗, 李志鹏, 赵健. 海岛型城市红火蚁发生程度空间格局及驱动因子——以福建海坛岛为例[J]. 生物多样性, 2023, 31(5): 22501-. |
| [15] | 杨清, 李晓东, 杨胜娴, 巢欣, 刘惠秋, 巴桑. 雅鲁藏布江中游丰水期原生动物群落多样性及其影响因子[J]. 生物多样性, 2023, 31(4): 22500-. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
备案号:京ICP备16067583号-7
Copyright © 2022 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn