
生物多样性 ›› 2017, Vol. 25 ›› Issue (9): 981-989. DOI: 10.17520/biods.2017052 cstr: 32101.14.biods.2017052
李宇航1, 陈万东2, 蔡厚才2, 孙忠民1,4, 徐奎栋1,3,4,*( )
)
收稿日期:2017-02-21
接受日期:2017-07-26
出版日期:2017-09-20
发布日期:2017-10-04
通讯作者:
徐奎栋
作者简介:# 共同第一作者
基金资助:
Yuhang Li1, Wandong Chen2, Houcai Cai2, Zhongmin Sun1,4, Kuidong Xu1,3,4,*()
Received:2017-02-21
Accepted:2017-07-26
Online:2017-09-20
Published:2017-10-04
Contact:
Xu Kuidong
About author:# Co-first authors
摘要:
为了解南麂列岛国家级海洋自然保护区砂质潮间带底栖硅藻多样性与群落结构的现状及近20多年来的变化, 于2013年11月至2014年8月, 对南麂列岛火焜岙砂质潮间带的底栖硅藻进行了4个季节的采样和研究, 并与1981-1993年有关的历史资料进行了比较分析。本研究共鉴定底栖硅藻49属120种, 海岸曲解藻(Fallacia litoricola)、史氏双壁藻(Diploneis smithii)、稀疏双壁藻(D. parca)等17种为目前的优势种。Shannon多样性指数在2.388-3.445之间, 以春季最高, 秋季最低; 在空间分布上依次为: 中潮区 > 低潮区 > 高潮区。相似性分析(analysis of similarities, ANOSIM)表明底栖硅藻群落在不同潮区间差异显著, 而季节差异不显著。BIOENV分析显示盐度与底栖硅藻群落结构的相关性最高。本研究结果表明, 南麂列岛砂质潮间带的底栖硅藻群落结构近几十年来已发生了明显变化。与1981-1993年南麂列岛的3次调查数据相比, 目级阶元减少了2个, 科级阶元增加了7个, 而属级和种级阶元较过去的29属55种有了显著增加, 这可能是分类研究强度增加所致。但分类学多样性降低, 平均分类差异指数Δ+由过去的79.79降至71.41; 且过去记录的大个体固着类群被现今的小个体固着类群和间隙运动类群(epipelon)所取代, 这可能是火焜岙过去人类活动频繁、有机质过量排放的长期效应所致。
李宇航, 陈万东, 蔡厚才, 孙忠民, 徐奎栋 (2017) 南麂列岛砂质潮间带底栖硅藻多样性与群落结构的时空变化. 生物多样性, 25, 981-989. DOI: 10.17520/biods.2017052.
Yuhang Li, Wandong Chen, Houcai Cai, Zhongmin Sun, Kuidong Xu (2017) Spatio-temporal variation of benthic diatom diversity and community structure in a sandy intertidal zone of the Nanji Islands, China. Biodiversity Science, 25, 981-989. DOI: 10.17520/biods.2017052.

图1 南麂列岛火焜岙潮间带底栖硅藻采样站位。H、M、L分别表示高、中、低潮区。
Fig. 1 Sampling sites of benthic diatoms from the intertidal zone of Huokun’ao in the Nanji Islands. H, M, and L represent high tide zone, middle tide zone and low tide zone, respectively.
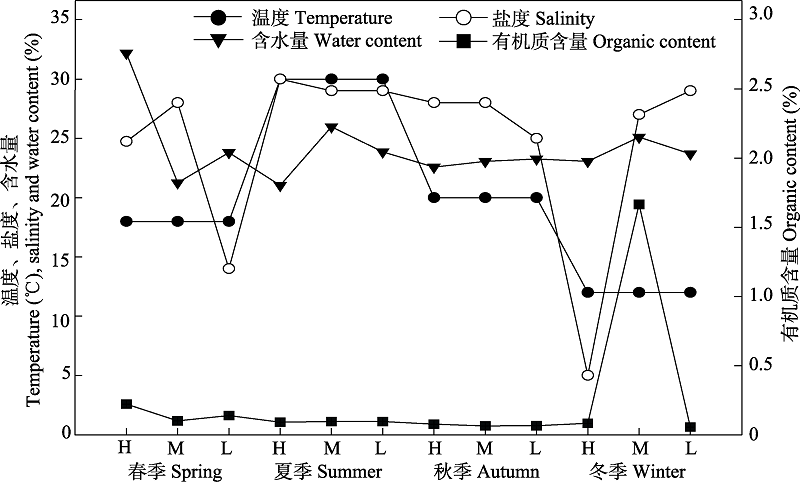
图2 南麂列岛火焜岙潮间带环境因子的季节变化。H、M、L分别表示高、中、低潮区。
Fig. 2 Seasonal changes in environmental factors in the intertidal zone of Huokun’ao, Nanji Islands. H, M, and L represent high tide zone, middle tide zone and low tide zone, respectively.
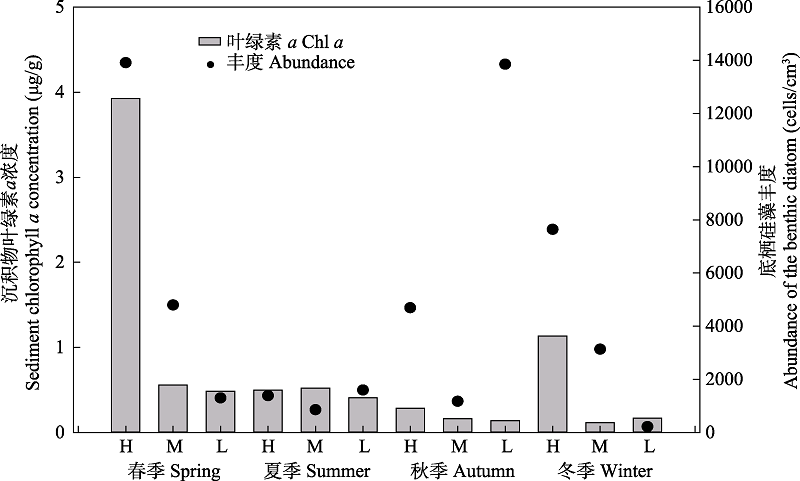
图3 南麂列岛火焜岙潮间带底栖硅藻丰度和沉积物中叶绿素a浓度。H、M、L分别表示高、中、低潮区。
Fig. 3 Diatom abundance and sediments chlorophyll a concentration in intertidal zone of Huokun’ao, Nanji Islands. H, M, and L represent high tide zone, middle tide zone and low tide zone, respectively.
| 物种 Species | 优势度 Dominant index (Y) |
|---|---|
| 海岸曲解藻 Fallacia litoricola | 0.1942 |
| 史氏双壁藻 Diploneis smithii | 0.1780 |
| 稀疏双壁藻 Diploneis parca | 0.1585 |
| 似叶状舟形藻 Navicula phylleptosoma | 0.0718 |
| 匙形菱形藻 Nitzschia spathulata | 0.0550 |
| 钳状曲解藻 Fallacia forcipata | 0.0522 |
| 沙生舟形藻 Navicula arenaria | 0.0520 |
| 琴状福氏藻 Fogedia lyra | 0.0509 |
| 矩形羽纹藻Pinnularia rectangulata | 0.0447 |
| 方格舟形藻 Navicula cancellata | 0.0397 |
| 线性蹄状藻 Hippodonta linearis | 0.0274 |
| 格氏双眉藻 Amphora graeffeana | 0.0273 |
| 直菱板藻加拉变种 Hantzschia virgata var. kariana | 0.0264 |
| 海岸曲解藻长圆变种 Diploneis litoralis var. clathrata | 0.0254 |
| 新西兰美壁藻 Diploneis novaeseelandiae | 0.0243 |
| 蹄状藻一种 Hippodonta sp. | 0.0234 |
| 海洋菱板藻 Hantzschia marina | 0.0223 |
表1 南麂列岛火焜岙潮间带底栖硅藻优势种
Table 1 Dominant species of benthic diatoms in the intertidal zone of Huokun’ao, Nanji Islands
| 物种 Species | 优势度 Dominant index (Y) |
|---|---|
| 海岸曲解藻 Fallacia litoricola | 0.1942 |
| 史氏双壁藻 Diploneis smithii | 0.1780 |
| 稀疏双壁藻 Diploneis parca | 0.1585 |
| 似叶状舟形藻 Navicula phylleptosoma | 0.0718 |
| 匙形菱形藻 Nitzschia spathulata | 0.0550 |
| 钳状曲解藻 Fallacia forcipata | 0.0522 |
| 沙生舟形藻 Navicula arenaria | 0.0520 |
| 琴状福氏藻 Fogedia lyra | 0.0509 |
| 矩形羽纹藻Pinnularia rectangulata | 0.0447 |
| 方格舟形藻 Navicula cancellata | 0.0397 |
| 线性蹄状藻 Hippodonta linearis | 0.0274 |
| 格氏双眉藻 Amphora graeffeana | 0.0273 |
| 直菱板藻加拉变种 Hantzschia virgata var. kariana | 0.0264 |
| 海岸曲解藻长圆变种 Diploneis litoralis var. clathrata | 0.0254 |
| 新西兰美壁藻 Diploneis novaeseelandiae | 0.0243 |
| 蹄状藻一种 Hippodonta sp. | 0.0234 |
| 海洋菱板藻 Hantzschia marina | 0.0223 |
| 春季 Spring | 夏季 Summer | 秋季 Autumn | 冬季 Winter | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| H | M | L | H | M | L | H | M | L | H | M | L | |
| J° | 0.7861 | 0.8761 | 0.7394 | 0.7785 | 0.8053 | 0.8284 | 0.6955 | 0.8034 | 0.8681 | 0.8644 | 0.8301 | 0.8108 |
| H' | 2.817 | 3.445 | 2.629 | 2.648 | 2.525 | 2.991 | 2.388 | 2.648 | 2.722 | 2.968 | 2.902 | 2.702 |
| 1-λ | 0.9112 | 0.9549 | 0.8511 | 0.8766 | 0.8856 | 0.9304 | 0.839 | 0.8974 | 0.9103 | 0.9302 | 0.9199 | 0.9041 |
表2 南麂列岛火焜岙潮间带不同季节底栖硅藻的Shannon多样性(H° )、均匀度(J° )和优势度(1-λ)。H、M、L分别表示高、中、低潮区。
Table 2 Diversity (H° ), evenness (J° ) and dominance index (1-λ) of benthic diatoms in the intertidal zone of Huokun’ao, Nanji Islands. H, M, and L represent high tide zone, middle tide zone, and low tide zone, respectively.
| 春季 Spring | 夏季 Summer | 秋季 Autumn | 冬季 Winter | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| H | M | L | H | M | L | H | M | L | H | M | L | |
| J° | 0.7861 | 0.8761 | 0.7394 | 0.7785 | 0.8053 | 0.8284 | 0.6955 | 0.8034 | 0.8681 | 0.8644 | 0.8301 | 0.8108 |
| H' | 2.817 | 3.445 | 2.629 | 2.648 | 2.525 | 2.991 | 2.388 | 2.648 | 2.722 | 2.968 | 2.902 | 2.702 |
| 1-λ | 0.9112 | 0.9549 | 0.8511 | 0.8766 | 0.8856 | 0.9304 | 0.839 | 0.8974 | 0.9103 | 0.9302 | 0.9199 | 0.9041 |
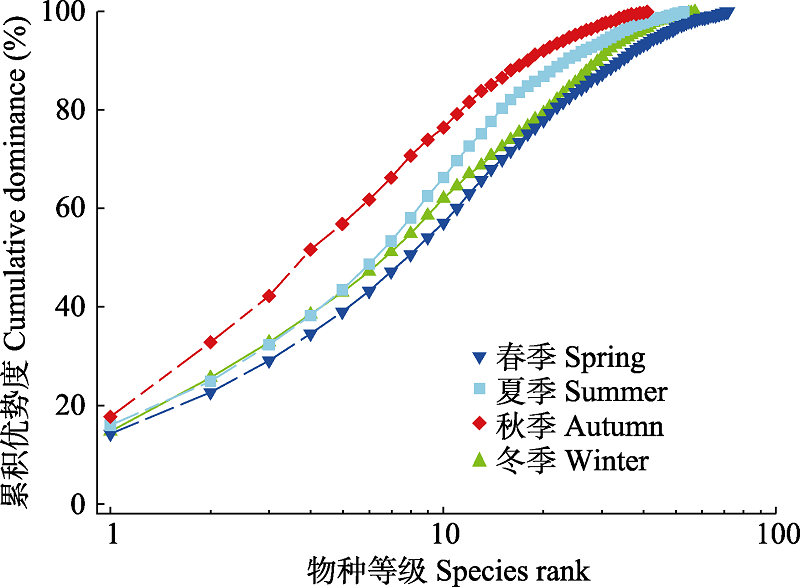
图4 南麂列岛火焜岙潮间带不同季节底栖硅藻群落的K优势曲线
Fig. 4 K-dominance curve of the benthic diatom community in the intertidal zone of Huokun’ao, Nanji Islands
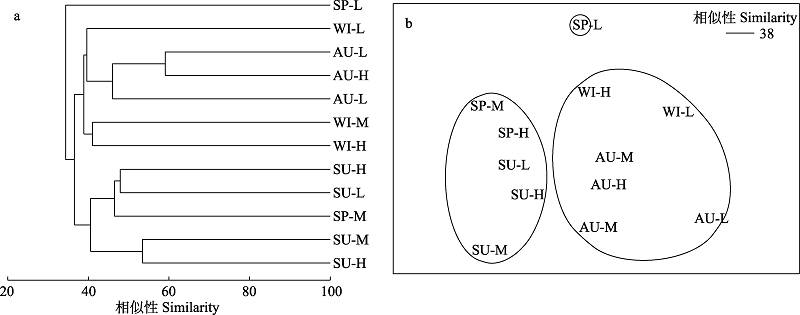
图5 南麂列岛火焜岙潮间带底栖硅藻的等级聚类和非参数多维标度排序。SP、SU、AU、WI分别代表春季、夏季、秋季、冬季; H、M、L分别表示高、中、低潮区。
Fig. 5 Clustering and Non-metric Multidimensional scaling (MDS) of benthic diatom community in the intertidal zone of Huokun’ao, Nanji Islands. SP, SU, AU, and WI represent spring, summer, autumn and winter, respectively. H, M, and L represent high tide zone, middle tide zone, and low tide zone, respectively.
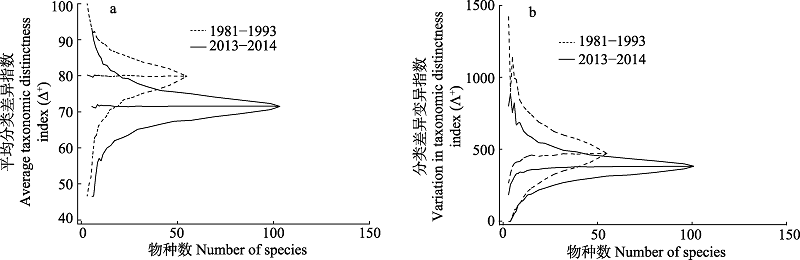
图6 南麂列岛火焜岙潮间带底栖硅藻分类差异性指数(Δ+)和分类差异性变异指数(Λ+)与历史资料(朱根海等1998a, b)比较的95%置信区间漏斗图, 分类阶元间路径长度为1。
Fig. 6 95% confidence intervals of average taxonomic distinctness index (Δ+) and variation in taxonomic distinctness index (Λ+), comparing with benthic diatoms data between this study and 1981-1993 (Zhu et al, 1998a, b). The distance between each taxon was set to 1.
| 属 Genus | 1981-1993* | 2013-2014# | 属 Genus | 1981-1993* | 2013-2014# |
|---|---|---|---|---|---|
| 曲壳藻属 Achnanthes | + | + | 蹄状藻属 Hippodonta | + | |
| 长曲壳藻属 Achnanthidium | + | 泥生藻属 Luticola | + | ||
| 辐裥藻属 Actinoptychus | + | 胸隔藻属 Mastogloia | + | ||
| 双眉藻属 Amphora | + | + | 直链藻属 Melosira | + | + |
| 蛛网藻属 Arachnoidiscus | + | 栖沙藻属 Moreneis | + | ||
| 沟盘藻属 Aulacodiscus | + | 舟形藻属 Navicula | + | + | |
| 棍形藻属 Bacillaria | + | 菱形藻属 Nitzschia | + | + | |
| 中鼓藻属 Bellerochea | + | 齿状藻属 Odontella | + | + | |
| 伯克力藻属 Berkeleya | + | + | 书形藻属 Parlibellus | + | |
| 盒形藻属 Biddulphia | + | 石舟藻属 Petroneis | + | + | |
| 对纹藻属 Biremis | + | 羽纹藻属 Pinnularia | + | ||
| 美壁藻属 Caloneis | + | 斜脊藻属 Plagiotropis | + | ||
| 脊弯藻属 Carinasigma | + | 平面藻属 Planothidium | + | ||
| 链形藻属 Catenula | + | 斜纹藻属 Pleurosigma | + | + | |
| 梯舟藻属 Climaconeis | + | 柄链藻属 Podosira | + | ||
| 卵形藻属 Cocconeis | + | 普氏藻属 Proschkinia | + | ||
| 圆筛藻属 Coscinodiscus | + | + | 沙网藻属 Psammodictyon | + | |
| 筒柱藻属 Cylindrotheca | + | 杆线藻属 Rhabdonema | + | ||
| 桥弯藻属 Cymbella | + | 洛氏藻属 Roperia | + | ||
| 迪氏藻属 Dickieia | + | 半舟藻属 Seminavis | + | ||
| 双壁藻属 Diploneis | + | + | 骨条藻属 Skeletonema | + | |
| 唐氏藻属 Donkina | + | 辐节藻属 Stauroneis | + | + | |
| 内茧藻属 Entomoneis | + | + | 班盘藻属 Sticotodiscus | + | |
| 曲解藻属 Fallacia | + | + | 条纹藻属 Striatella | + | |
| 福氏藻属 Fogedia | + | 双菱藻属 Surirella | + | ||
| 斑条藻属 Grammatophora | + | + | 针杆藻属 Synedra | + | |
| 布纹藻属 Gyrosigma | + | + | 平片藻属 Tabularia | + | |
| 海生双眉藻属 Halamphora | + | 海链藻属 Thalassiosira | + | ||
| 菱板藻属 Hantzschia | + | + | 粗纹藻属 Trachyneis | + | + |
| 海氏藻属 Haslea | + | 盘杆藻属 Tryblionella | + |
表3 南麂列岛火焜岙潮间带底栖硅藻(属)的变化
Table 3 Changes of benthic diatom genera from the intertidal sandy sediments of Huokun’ao, Nanji Islands
| 属 Genus | 1981-1993* | 2013-2014# | 属 Genus | 1981-1993* | 2013-2014# |
|---|---|---|---|---|---|
| 曲壳藻属 Achnanthes | + | + | 蹄状藻属 Hippodonta | + | |
| 长曲壳藻属 Achnanthidium | + | 泥生藻属 Luticola | + | ||
| 辐裥藻属 Actinoptychus | + | 胸隔藻属 Mastogloia | + | ||
| 双眉藻属 Amphora | + | + | 直链藻属 Melosira | + | + |
| 蛛网藻属 Arachnoidiscus | + | 栖沙藻属 Moreneis | + | ||
| 沟盘藻属 Aulacodiscus | + | 舟形藻属 Navicula | + | + | |
| 棍形藻属 Bacillaria | + | 菱形藻属 Nitzschia | + | + | |
| 中鼓藻属 Bellerochea | + | 齿状藻属 Odontella | + | + | |
| 伯克力藻属 Berkeleya | + | + | 书形藻属 Parlibellus | + | |
| 盒形藻属 Biddulphia | + | 石舟藻属 Petroneis | + | + | |
| 对纹藻属 Biremis | + | 羽纹藻属 Pinnularia | + | ||
| 美壁藻属 Caloneis | + | 斜脊藻属 Plagiotropis | + | ||
| 脊弯藻属 Carinasigma | + | 平面藻属 Planothidium | + | ||
| 链形藻属 Catenula | + | 斜纹藻属 Pleurosigma | + | + | |
| 梯舟藻属 Climaconeis | + | 柄链藻属 Podosira | + | ||
| 卵形藻属 Cocconeis | + | 普氏藻属 Proschkinia | + | ||
| 圆筛藻属 Coscinodiscus | + | + | 沙网藻属 Psammodictyon | + | |
| 筒柱藻属 Cylindrotheca | + | 杆线藻属 Rhabdonema | + | ||
| 桥弯藻属 Cymbella | + | 洛氏藻属 Roperia | + | ||
| 迪氏藻属 Dickieia | + | 半舟藻属 Seminavis | + | ||
| 双壁藻属 Diploneis | + | + | 骨条藻属 Skeletonema | + | |
| 唐氏藻属 Donkina | + | 辐节藻属 Stauroneis | + | + | |
| 内茧藻属 Entomoneis | + | + | 班盘藻属 Sticotodiscus | + | |
| 曲解藻属 Fallacia | + | + | 条纹藻属 Striatella | + | |
| 福氏藻属 Fogedia | + | 双菱藻属 Surirella | + | ||
| 斑条藻属 Grammatophora | + | + | 针杆藻属 Synedra | + | |
| 布纹藻属 Gyrosigma | + | + | 平片藻属 Tabularia | + | |
| 海生双眉藻属 Halamphora | + | 海链藻属 Thalassiosira | + | ||
| 菱板藻属 Hantzschia | + | + | 粗纹藻属 Trachyneis | + | + |
| 海氏藻属 Haslea | + | 盘杆藻属 Tryblionella | + |
| [1] | Cai HC, Peng X (2001) Welcome to the Kingdom of Shellfish and Algae. Shanghai People’s Fine Arts Publishing House, Shanghai. (in Chinese) |
| [蔡厚才, 彭欣 (2011) 走进贝藻王国. 上海人民美术出版社, 上海.] | |
| [2] | Clarke KR, Gorley RN (2006) PRIMER v6: User Manual/ Tutorial (Plymouth Routines in Multivariate Ecological research). Primer-E Ltd., Plymouth. |
| [3] | Du GY, Chung IK (2007) Spatio-termporal variation of intertidal microphytobenthos in the Nakdong estuary. The Sea, 12, 186-190. |
| [4] |
Du GY, Son M, Yun M, An S, Chung IK (2009) Microphytobenthic biomass and species composition in intertidal flats of the Nakdong River estuary, Korea. Estuarine, Coastal and Shelf Science, 82, 663-672.
DOI URL |
| [5] |
Field CB, Behrenfeld MJ, Randerson JT, Falkowski P (1998) Primary production of the biosphere: integrating terrestrial and oceanic components. Science, 281, 237-240.
DOI URL |
| [6] |
Hendey NI (1964) An Introductory Account of the Smaller Algae of British Coastal Water. V. Bacillariophyceae (Diatoms). H. M. Stationary Office, London.
DOI URL |
| [7] | Jean-Marc G, Laurent C, Jennifer CG (2008) Can the intertidal benthic microalgal primary production account for the “Missing Carbon Sink”? Journal of Oceanography, Research and Data, 1, 13-19. |
| [8] |
Mann DG, Vanormelingen P (2013) An inordinate fondness? The number, distributions, and origins of diatom species. Journal of Eukaryotic Microbiology, 60, 414-420.
DOI URL PMID |
| [9] |
Medlin LK, Kaczmarska I (2004) Evolution of the diatoms. V. Morphological and cytological support for the major clades and a taxonomic revision. Phycologia, 43, 245-270.
DOI URL |
| [10] | Pielou EC (1975) Ecological Diversity. John Wiley & Sons Inc., New York. |
| [11] | Pielou EC (1984) The Interpretation of Ecological Data: A Primer on Classification and Ordination. John Wiley & Sons Inc., New York. |
| [12] |
Ribeiro L (2010) Intertidal Benthic Diatoms of the Tagus Estuary: Taxonomic Compositoin and Spatial-temporal Variation. PhD dissertation, University of Lisbon, Lisbon.
DOI URL |
| [13] |
Ribeiro L, Brotas V, Rincé Y, Jesus B (2013) Structure and diversity of intertidal benthic diatom assemblages in contrasting shores: a case study from the Tagus estuary. Journal of Phycology, 49, 258-270.
DOI URL PMID |
| [14] |
Sabbe K (1993) Short-term fluctuations in benthic diatom number on an intertidal sandflat in the Westerschelde estuary (Zeeland, The Netherlands). Hydrobiologia, 269/270, 275-284.
DOI URL |
| [15] | Sabbe K (1997) Systematics and Ecology of Intertidal Benthic Diatoms of the Westerschelde Estuary (The Netherlands). PhD dissertation, Ghent University, Ghent. |
| [16] |
Schindler DW (1999) Carbon cycling: the mysterious missing sink. Nature, 198, 105-107.
DOI URL |
| [17] |
Shannon CE (1948) A mathematical theory of communication. Bell System Technical Journal, 27, 379-423.
DOI URL |
| [18] |
Simpson EH (1949) Measurement of diversity. Nature, 163, 688.
DOI URL |
| [19] | Underwood GJC, Kromkamp J (1999) Primary production by phytoplankton and microphytobenthos in estuaries. In: Advances in Ecological Research (eds Nedwell DB, Raffaelli DG), pp. 93-153. Academic Press, London. |
| [20] | Yu YY (2011) Practices and Lessons from Island-based Conservation of Biodiversity in Nanji Islands, China. Ocean Press, Beijing. (in Chinese) |
| [俞永跃 (2011) 基于海岛管理的南麂列岛生物多样性保护实践与经验. 海洋出版社, 北京.] | |
| [21] | Zhu GH, Wang X, Wang CS, Gao AG (1998a) Ecological studies on nanoalgae and microalgae in Nanji Island National Marine Nature Conservation Area. I. Species composition and ecological characteristics. Journal of Marine Sciences, 16(2), 1-21. (in Chinese with English abstract) |
| [朱根海, 王旭, 王春生, 高爱根 (1998a) 南麂列岛国家海洋自然保护区微、小型藻类生态研究. I. 种类组成与生态特点. 东海海洋, 16(2), 1-21.] | |
| [22] | Zhu GH, Wang X, Wang CS, Gao AG (1998b) Ecological studies on nanoalgae and microalgae in Nanji Islands National Marine Natural Conservation Area. II. Quantitative distribution. Journal of Marine Sciences, 16(2), 22-28. (in Chinese with English abstract) |
| [朱根海, 王旭, 王春生, 高爱根 (1998b) 南麂列岛国家海洋自然保护区微、小型藻类生态研究. II. 数量分布. 东海海洋, 16(2), 23-28.] |
| [1] | 吴晓晴 张美惠 葛苏婷 李漫淑 宋坤 沈国春 达良俊 张健. 上海近自然林重建过程中木本植物物种多样性与地上生物量的时空动态——以闵行区生态岛为例[J]. 生物多样性, 2025, 33(5): 24444-. |
| [2] | 干靓 刘巷序 鲁雪茗 岳星. 全球生物多样性热点地区大城市的保护政策与优化方向[J]. 生物多样性, 2025, 33(5): 24529-. |
| [3] | 曾子轩 杨锐 黄越 陈路遥. 清华大学校园鸟类多样性特征与环境关联[J]. 生物多样性, 2025, 33(5): 24373-. |
| [4] | 周昊, 王茗毅, 张楚格, 肖治术, 欧阳芳. 昆虫旅馆在独栖蜂多样性保护中的现状与挑战[J]. 生物多样性, 2025, 33(5): 24472-. |
| [5] | 臧明月, 刘立, 马月, 徐徐, 胡飞龙, 卢晓强, 李佳琦, 于赐刚, 刘燕. 《昆明-蒙特利尔全球生物多样性框架》下的中国城市生物多样性保护[J]. 生物多样性, 2025, 33(5): 24482-. |
| [6] | 祝晓雨, 王晨灏, 王忠君, 张玉钧. 城市绿地生物多样性研究进展与展望[J]. 生物多样性, 2025, 33(5): 25027-. |
| [7] | 袁琳, 王思琦, 侯静轩. 大都市地区的自然留野:趋势与展望[J]. 生物多样性, 2025, 33(5): 24481-. |
| [8] | 胡敏, 李彬彬, Coraline Goron. 只绿是不够的: 一个生物多样性友好的城市公园管理框架[J]. 生物多样性, 2025, 33(5): 24483-. |
| [9] | 王欣, 鲍风宇. 基于鸟类多样性提升的南滇池国家湿地公园生态修复效果分析[J]. 生物多样性, 2025, 33(5): 24531-. |
| [10] | 明玥, 郝培尧, 谭铃千, 郑曦. 基于城市绿色高质量发展理念的中国城市生物多样性保护与提升研究[J]. 生物多样性, 2025, 33(5): 24524-. |
| [11] | 徐欢, 辛凤飞, 施宏亮, 袁琳, 薄顺奇, 赵欣怡, 邓帅涛, 潘婷婷, 余婧, 孙赛赛, 薛程. 生态修复技术集成应用对长江口北支生境与鸟类多样性提升效果评估[J]. 生物多样性, 2025, 33(5): 24478-. |
| [12] | 谢淦, 宣晶, 付其迪, 魏泽, 薛凯, 雒海瑞, 高吉喜, 李敏. 草地植物多样性无人机调查的物种智能识别模型构建[J]. 生物多样性, 2025, 33(4): 24236-. |
| [13] | 王太, 宋福俊, 张永胜, 娄忠玉, 张艳萍, 杜岩岩. 河西走廊内陆河水系鱼类多样性及资源现状[J]. 生物多样性, 2025, 33(4): 24387-. |
| [14] | 褚晓琳, 张全国. 演化速率假说的实验验证研究进展[J]. 生物多样性, 2025, 33(4): 25019-. |
| [15] | 张浩斌, 肖路, 刘艳杰. 夜间灯光对外来入侵植物和本地植物群落多样性和生长的影响[J]. 生物多样性, 2025, 33(4): 24553-. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
备案号:京ICP备16067583号-7
Copyright © 2022 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn