
生物多样性 ›› 2019, Vol. 27 ›› Issue (12): 1298-1308. DOI: 10.17520/biods.2019157 cstr: 32101.14.biods.2019157
刘硕然1,2,3,杨道德1,*( ),李先福2,3,4,谭路4,孙军5,和晓阳5,杨文书5,任国鹏2,3,Davide Fornacca2,3,蔡庆华4,肖文2,3,*()
),李先福2,3,4,谭路4,孙军5,和晓阳5,杨文书5,任国鹏2,3,Davide Fornacca2,3,蔡庆华4,肖文2,3,*()
收稿日期:2019-05-08
接受日期:2019-09-26
出版日期:2019-12-20
发布日期:2020-02-22
通讯作者:
杨道德,肖文
基金资助:
Shuoran Liu1,2,3,Daode Yang1,*(),Xianfu Li2,3,4,Lu Tan4,Jun Sun5,Xiaoyang He5,Wenshu Yang5,Guopeng Ren2,3,Davide Fornacca2,3,Qinghua Cai4,Wen Xiao2,3,*()
Received:2019-05-08
Accepted:2019-09-26
Online:2019-12-20
Published:2020-02-22
Contact:
Yang Daode,Xiao Wen
摘要:
高山微水体由于面积微小且通过地表径流形成串联结构常常被认为与高山溪流具有类似的生境, 然而由于这两类生境中环境因子与底栖动物多样性存在差异, 它们在生态系统中的作用可能完全不同。滇西北地区是全球生物多样性热点区域之一, 境内高山微水体和高山溪流分布密集, 在区域底栖生物多样性维持方面具有重要的功能, 然而目前对这两类高山淡水生态系统的研究较少。为了比较这两类生境环境因子的异同及其对底栖动物多样性的维持作用, 2015年6月, 作者在云南省怒江州贡山县的高山峡谷内, 对27个高山微水体和同区域分布的1条高山溪流(海拔高差500 m范围)的底栖动物多样性和水环境因子进行了实地调查。结果表明: (1)高山微水体和高山溪流底栖动物群落中优势分类单元种群数量均比较庞大, 而稀有分类单元数量较多且种群较小; (2)两种生境在环境因子、物种多样性、功能多样性和群落结构方面的差异明显, 高山溪流有较高的物种丰富度、物种多样性和功能多样性; (3)高山微水体底栖动物多样性的分布与水环境因子无关, 而高山溪流底栖动物多样性与群落结构的形成受到与流速关联的水环境因子和海拔的影响。因此, 高山微水体与高山溪流不能简单地视为类似的生境类型, 它们对区域底栖动物多样性和生态功能维持可能具有不同的作用。
刘硕然, 杨道德, 李先福, 谭路, 孙军, 和晓阳, 杨文书, 任国鹏, Davide Fornacca, 蔡庆华, 肖文 (2019) 滇西北高山微水体与溪流生境底栖动物多样性和环境特征. 生物多样性, 27, 1298-1308. DOI: 10.17520/biods.2019157.
Shuoran Liu, Daode Yang, Xianfu Li, Lu Tan, Jun Sun, Xiaoyang He, Wenshu Yang, Guopeng Ren, Davide Fornacca, Qinghua Cai, Wen Xiao (2019) Diversity in benthic and environmental characteristics on alpine micro-waterbodies and stream ecosystems in northwest Yunnan. Biodiversity Science, 27, 1298-1308. DOI: 10.17520/biods.2019157.
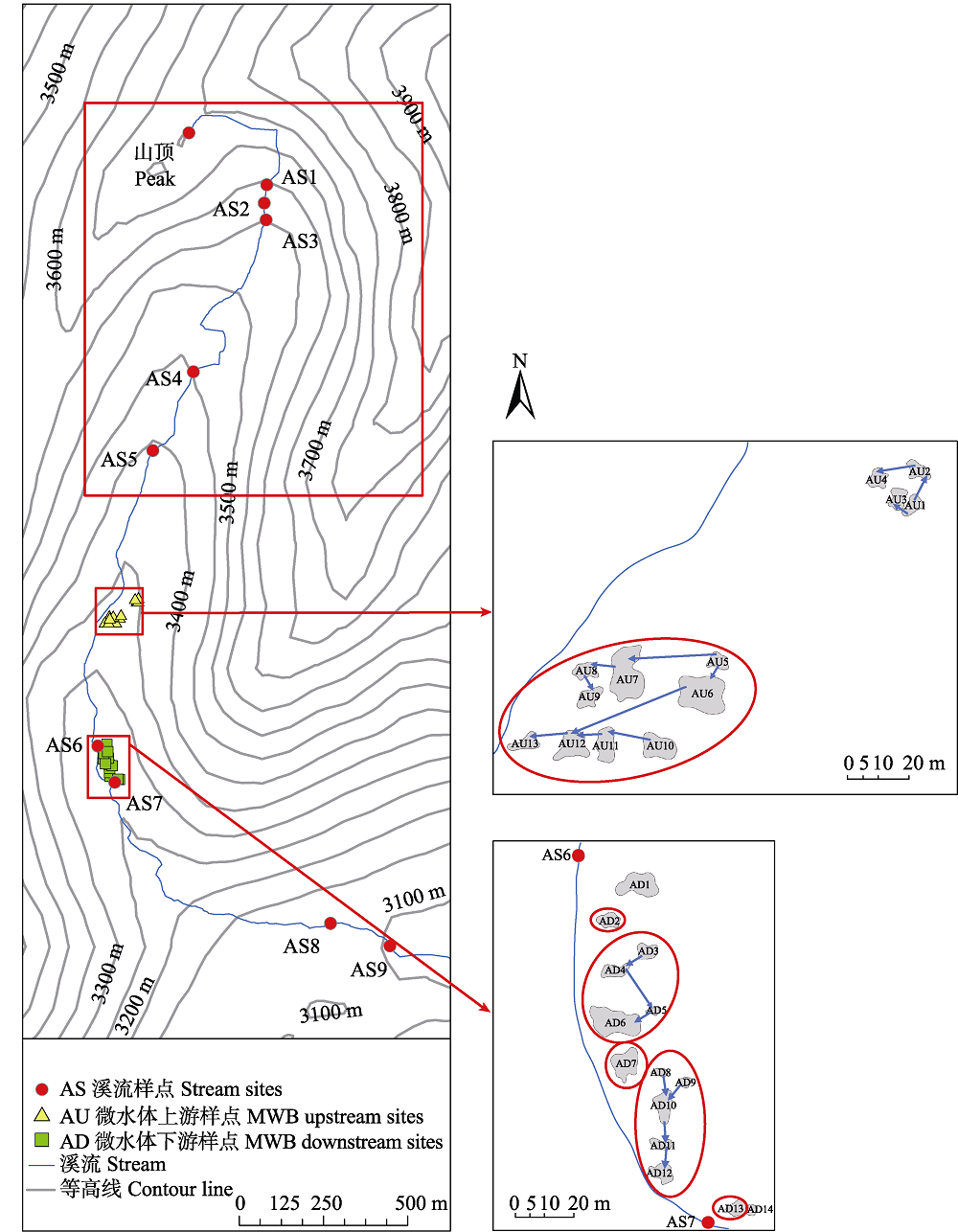
图1 研究区域及样点图示(微水体串联图示中圈内的微水体串最终与溪流连通)
Fig. 1 Research area and sampling sites. The micro-waterbodies (MWB) which are circled indicate that the water flow from the micro-waterbody cascades is running into the stream finally.
| 环境因子 Environmental variables | 微水体 Micro-waterbody | 溪流 Stream | ||||
|---|---|---|---|---|---|---|
| 最小值 Min. | 最大值 Max. | 平均值 ± 标准差 Mean ± SD | 最小值 Min. | 最大值 Max. | 平均值 ± 标准差 Mean ± SD | |
| 海拔 Alt (m) ns2 | 3,266.00 | 3,332.00 | 3,299.70 ± 27.34 | 3,087.00 | 3,586.00 | 3,364.90 ± 188.82 |
| 电导率 Cond (μs/cm) **2 | 2.94 | 10.45 | 5.12 ± 1.33 | 10.82 | 34.50 | 17.54 ± 8.84 |
| 溶解氧含量 DO (mg/L) **1 | 3.13 | 7.51 | 5.68 ± 0.92 | 6.22 | 7.18 | 6.72 ± 0.27 |
| pH值 pH **1 | 5.48 | 6.20 | 5.84 ± 0.24 | 5.80 | 6.55 | 6.23 ± 0.26 |
| 面积 Area (m2) none | 3.00 | 160.00 | 26.15 ± 33.65 | / | / | / |
| 水深 Depth (cm) *1 | 8.00 | 43.00 | 27.70 ± 10.09 | 6.00 | 26.00 | 18.33 ± 6.48 |
| 底泥深度 BSD (cm) none | 9.50 | 58.00 | 26.09 ± 13.02 | / | / | / |
| 浊度 Turb (NTU) ns1 | 0.00 | 3.70 | 1.42 ± 1.08 | 0.00 | 2.40 | 0.98 ± 1.06 |
| 总氮 TN (mg/L) ns1 | 0.096 | 0.729 | 0.300 ± 0.170 | 0.115 | 0.445 | 0.280 ± 0.110 |
| 总磷 TP (mg/L) ns1 | 0.024 | 0.104 | 0.046 ± 0.016 | 0.036 | 0.068 | 0.050 ± 0.010 |
| 化学需氧量 COD (mg/L) *1 | 0.160 | 4.730 | 2.150 ± 1.290 | 0.160 | 5.056 | 3.320 ± 1.420 |
| 总有机碳含量 TOC (mg/L) ns1 | 0.924 | 7.590 | 4.270 ± 1.640 | 3.043 | 6.963 | 4.800 ± 1.070 |
| 溪流宽度 Width (m) none | / | / | / | 0.50 | 5.30 | 2.53 ±1.78 |
| 溪流流速 FV (m/s) none | / | / | / | 0.16 | 0.93 | 0.58 ± 0.31 |
表1 溪流与微水体各环境因子描述及差异性分析
Table 1 List of the descriptive statistics and difference in environmental variables between stream and micro-waterbody
| 环境因子 Environmental variables | 微水体 Micro-waterbody | 溪流 Stream | ||||
|---|---|---|---|---|---|---|
| 最小值 Min. | 最大值 Max. | 平均值 ± 标准差 Mean ± SD | 最小值 Min. | 最大值 Max. | 平均值 ± 标准差 Mean ± SD | |
| 海拔 Alt (m) ns2 | 3,266.00 | 3,332.00 | 3,299.70 ± 27.34 | 3,087.00 | 3,586.00 | 3,364.90 ± 188.82 |
| 电导率 Cond (μs/cm) **2 | 2.94 | 10.45 | 5.12 ± 1.33 | 10.82 | 34.50 | 17.54 ± 8.84 |
| 溶解氧含量 DO (mg/L) **1 | 3.13 | 7.51 | 5.68 ± 0.92 | 6.22 | 7.18 | 6.72 ± 0.27 |
| pH值 pH **1 | 5.48 | 6.20 | 5.84 ± 0.24 | 5.80 | 6.55 | 6.23 ± 0.26 |
| 面积 Area (m2) none | 3.00 | 160.00 | 26.15 ± 33.65 | / | / | / |
| 水深 Depth (cm) *1 | 8.00 | 43.00 | 27.70 ± 10.09 | 6.00 | 26.00 | 18.33 ± 6.48 |
| 底泥深度 BSD (cm) none | 9.50 | 58.00 | 26.09 ± 13.02 | / | / | / |
| 浊度 Turb (NTU) ns1 | 0.00 | 3.70 | 1.42 ± 1.08 | 0.00 | 2.40 | 0.98 ± 1.06 |
| 总氮 TN (mg/L) ns1 | 0.096 | 0.729 | 0.300 ± 0.170 | 0.115 | 0.445 | 0.280 ± 0.110 |
| 总磷 TP (mg/L) ns1 | 0.024 | 0.104 | 0.046 ± 0.016 | 0.036 | 0.068 | 0.050 ± 0.010 |
| 化学需氧量 COD (mg/L) *1 | 0.160 | 4.730 | 2.150 ± 1.290 | 0.160 | 5.056 | 3.320 ± 1.420 |
| 总有机碳含量 TOC (mg/L) ns1 | 0.924 | 7.590 | 4.270 ± 1.640 | 3.043 | 6.963 | 4.800 ± 1.070 |
| 溪流宽度 Width (m) none | / | / | / | 0.50 | 5.30 | 2.53 ±1.78 |
| 溪流流速 FV (m/s) none | / | / | / | 0.16 | 0.93 | 0.58 ± 0.31 |
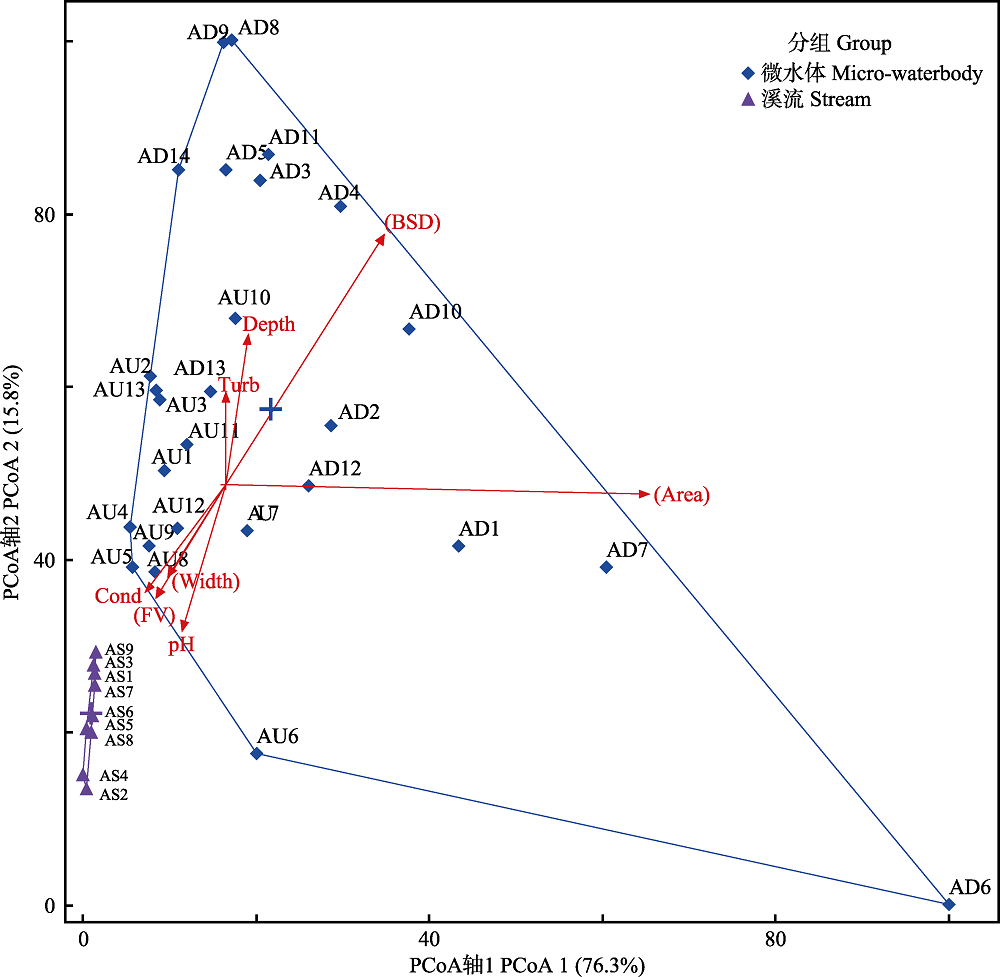
图2 基于环境因子的PCoA双标图。AS为溪流样点, AU、AD为微水体样点。Depth: 水深; Area: 面积; Width: 溪流宽度; FV:流速; Cond: 电导率; Turb: 浊度。
Fig. 2 Biplot of PCoA calculated based on environmental factors. AS indicates stream sites, AU and AD indicate micro-waterbody sites. Depth, Water depth; Area, Waterbody surface area; Width, Stream width; FV, Flow velocity; Cond, Conductivity; Turb, Turbidity.
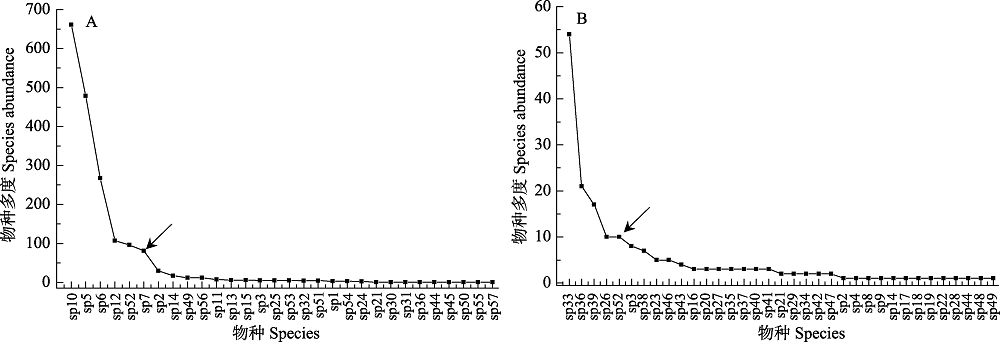
图3 高山微水体(A)及溪流(B)分类单元多度排序曲线(箭头所示为曲线拐点, 作为优势分类单元与稀有分类单元分界), 横坐标为物种排列顺序,对应附录1。
Fig. 3 Species abundance ranking curves for alpine micro-waterbody (A) and stream (B). Arrows indicate the inflection point of the curve between the dominant taxa and rare taxa. X axis refers to the species order in Appendix 1.
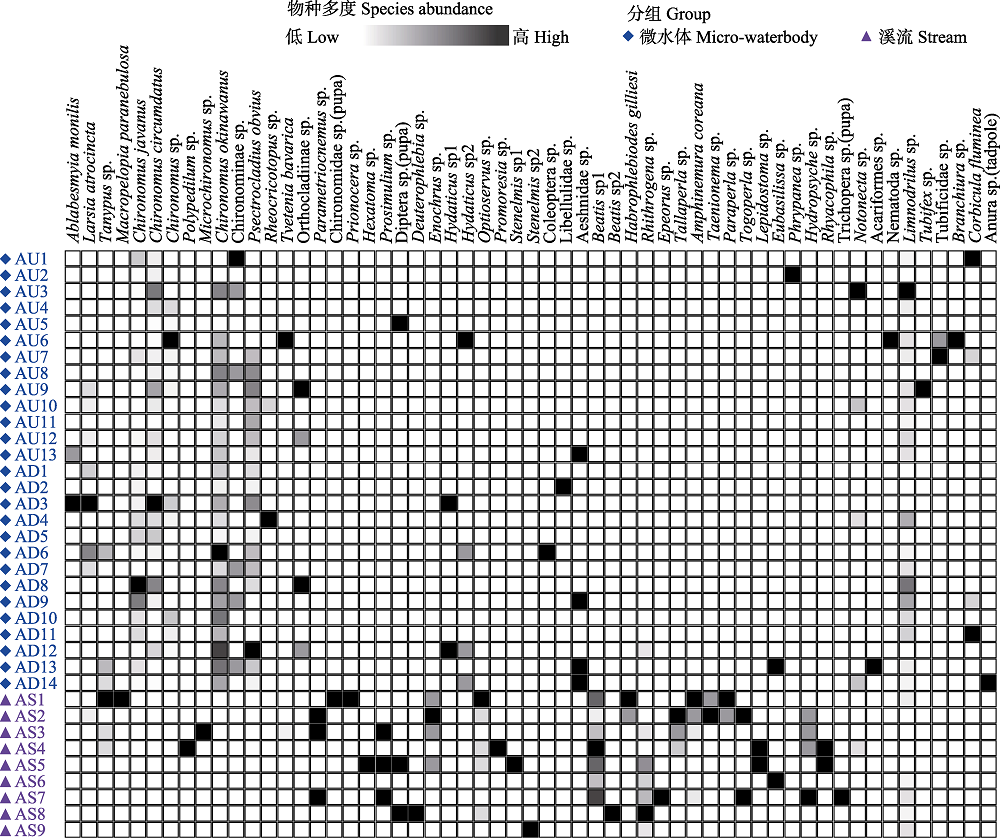
图4 底栖动物物种在样点间分布热图(分类单元物种多度以颜色深浅区分)
Fig. 4 Heatmap of the benthic species distribution among the collection sites. The species abundance for each taxon marked by the color shade
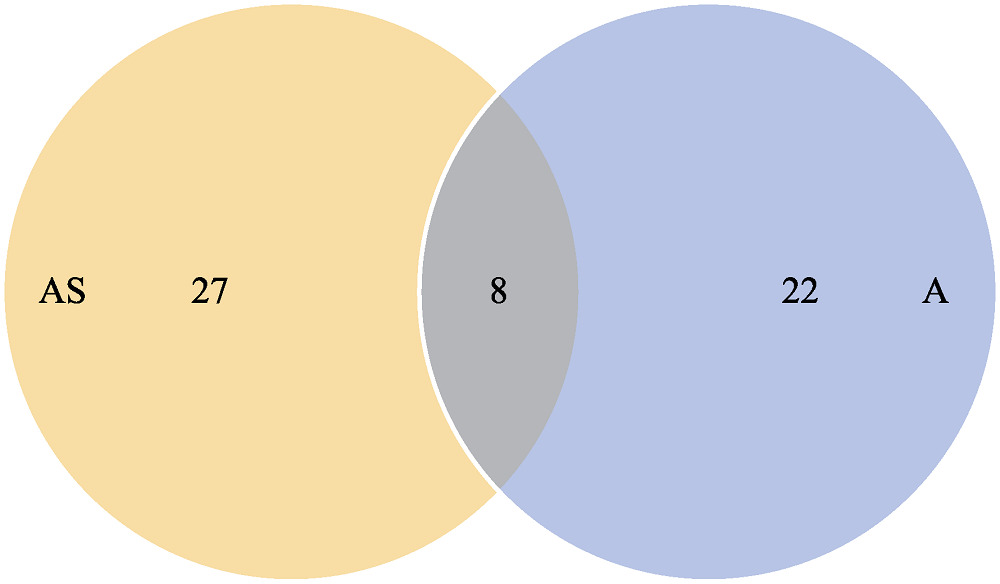
图5 溪流与微水体分类单元丰富度维恩图
Fig. 5 The Venn diagram between the micro-waterbody and stream by taxon richness
| 微水体 Micro-waterbody | 溪流 Stream | |||||
|---|---|---|---|---|---|---|
| 最小值 Min. | 最大值 Max. | 平均值 ± 标准差 Mean ± SD | 最小 值 Min. | 最大 值 Max. | 平均值 ± 标准差 Mean ± SD | |
| S ns1 | 3.00 | 10.00 | 6.26 ± 2.07 | 3.00 | 12.00 | 8.67 ± 3.43 |
| H' *1 | 0.882 | 1.649 | 1.240 ± 0.250 | 1.011 | 2.265 | 1.720 ± 0.460 |
| FDis **1 | 0.000 | 0.469 | 0.220 ± 0.130 | 0.544 | 0.968 | 0.820 ± 0.130 |
表2 溪流与微水体底栖动物群落的物种丰富度(S)、物种多样性(H')和功能多样性(FDis)
Table 2 List of the species richness, species diversity and functional diversity between stream and micro-waterbody
| 微水体 Micro-waterbody | 溪流 Stream | |||||
|---|---|---|---|---|---|---|
| 最小值 Min. | 最大值 Max. | 平均值 ± 标准差 Mean ± SD | 最小 值 Min. | 最大 值 Max. | 平均值 ± 标准差 Mean ± SD | |
| S ns1 | 3.00 | 10.00 | 6.26 ± 2.07 | 3.00 | 12.00 | 8.67 ± 3.43 |
| H' *1 | 0.882 | 1.649 | 1.240 ± 0.250 | 1.011 | 2.265 | 1.720 ± 0.460 |
| FDis **1 | 0.000 | 0.469 | 0.220 ± 0.130 | 0.544 | 0.968 | 0.820 ± 0.130 |
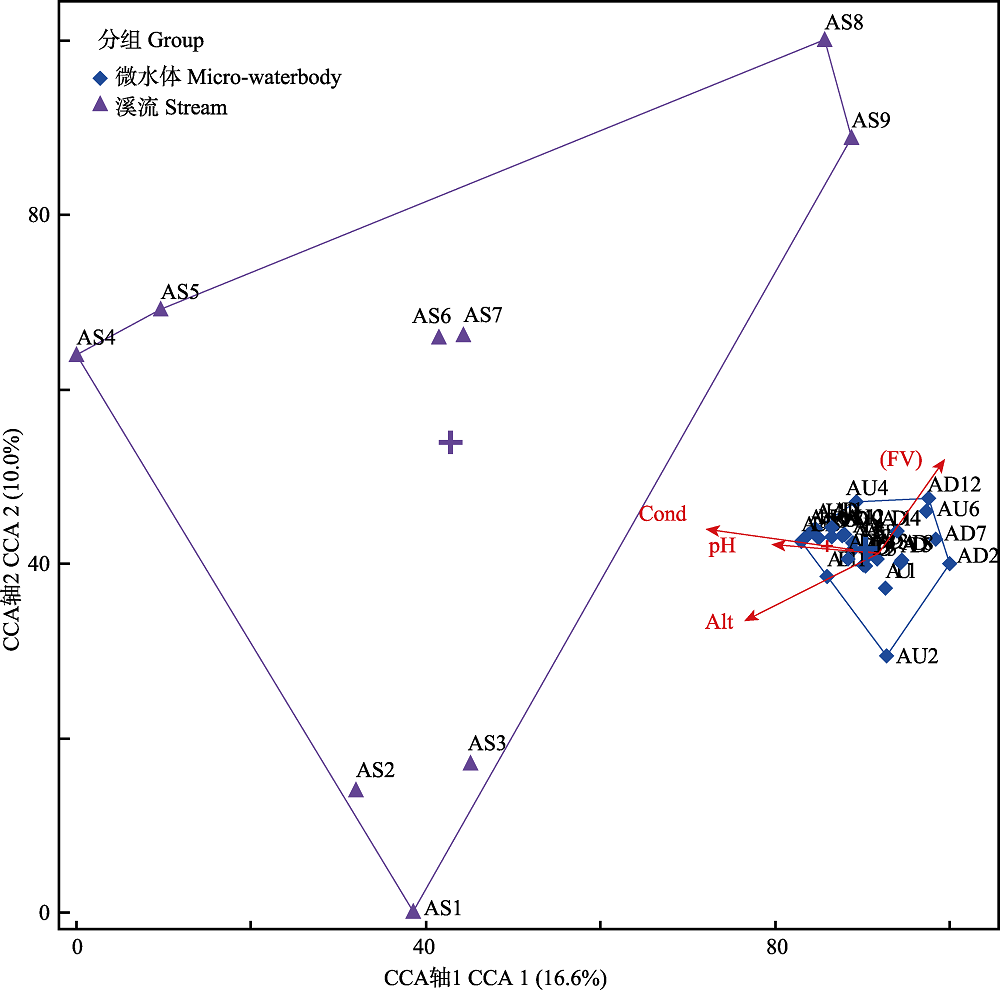
图6 基于底栖动物群落数据和环境因子的CCA双标图。AS为溪流样点, AU、AD为微水体样点。Alt: 海拔; FV: 流速; Cond: 电导率。
Fig. 6 Biplot of CCA which calculated based on the benthic community data and environmental factors. hydrogen ion concentration (pH), flow velocity (FV) and conductivity (Cond). AS indicates stream sites, AU and AD indicate micro-waterbody sites. Alt, Altitude; FV, Flow velocity; Cond, Conductivity.
| [1] | Altermatt F ( 2013) Diversity in riverine metacommunities: A network perspective. Aquatic Ecology, 47, 365-377. |
| [2] | Bazzanti M, Bella VD, Seminara M ( 2003) Factors affecting macroinvertebrate communities in astatic ponds in central Italy. Journal of Freshwater Ecology, 18, 537-548. |
| [3] | Belletti B, Rinaldi M, Buijse AD, Gurnell AM, Mosselman E ( 2015) A review of assessment methods for river hydromorphology. Environmental Earth Sciences, 73, 2079-2100. |
| [4] | Belmar O, Velasco J, Gutierrezcanovas C, Melladodiaz A, Millan A, Wood PJ ( 2013) The influence of natural flow regimes on macroinvertebrate assemblages in a semiarid Mediterranean basin. Ecohydrology, 6, 363-379. |
| [5] | Biggs J, Williams P, Whitfield M, Nicolet P, Weatherby AJ ( 2005) 15 years of pond assessment in Britain: Results and lessons learned from the work of pond conservation. Aquatic Conservation-Marine and Freshwater Ecosystems, 15, 693-714. |
| [6] | Bogan MT, Boersma KS, Lytle DA ( 2013) Flow intermittency alters longitudinal patterns of invertebrate diversity and assemblage composition in an arid-land stream network. Freshwater Biology, 58, 1016-1028. |
| [7] | Bonada N, Rieradevall M, Prat N ( 2007) Macroinvertebrate community structure and biological traits related to flow permanence in a Mediterranean river network. Hydrobiologia, 589, 91-106. |
| [8] | Borcard D, Legendre P ( 2002) All-scale spatial analysis of ecological data by means of principal coordinates of neighbour matrices. Ecological Modelling, 153, 51-68. |
| [9] | Carrara F, Altermatt F, Rodrigueziturbe I, Rinaldo A ( 2012) Dendritic connectivity controls biodiversity patterns in experimental metacommunities. Proceedings of the National Academy of Sciences, USA, 109, 5761-5766. |
| [10] | Carrara F, Rinaldo A, Giometto A, Altermatt F ( 2014) Complex interaction of dendritic connectivity and hierarchical patch size on biodiversity in river-like landscapes. The American Naturalist, 183, 13-25. |
| [11] | Chakona A, Phiri C, Magadza CH, Brendonck L ( 2008) The influence of habitat structure and flow permanence on macroinvertebrate assemblages in temporary rivers in northwestern Zimbabwe. Hydrobiologia, 607, 199-209. |
| [12] | Dudgeon D, Arthington AH, Gessner MO, Kawabata ZI, Knowler D, Leveque C, Naiman RJ, Prieur-Richard AH, Soto D, Stiassny MLJ, Sullivan CA ( 2006) Freshwater biodiversity: Importance, threats, status and conservation challenges. Biological Reviews, 81, 163-182. |
| [13] | Henriques-Oliveira AL, Nessimian JL ( 2010) Aquatic macroinvertebrate diversity and composition in streams along an altitudinal gradient in southeastern Brazil. Biota Neotropica, 10, 115-128. |
| [14] | Hoffman M, Koenig K, Bunting G, Costanza J, Kristen JM ( 2016) Biodiversity Hotspots (version 2016.1). . (accessed on 2019-03-18) |
| [15] | Laliberte E, Legendre PA ( 2010) Distance-based framework for measuring functional diversity from multiple traits. Ecology, 91, 299-305. |
| [16] | Leibold MA, Holyoak M, Mouquet N, Amarasekare P, Chase JM, Hoopes MF, Holt RD, Shurin JB, Tilman D, Loreau M, Gonzalez A ( 2004) The metacommunity concept: A framework for multi-scale community ecology. Ecology Letters, 7, 601-613. |
| [17] | Liu SR, He XY, Yang WS, Ren GP, Li YP, Zhou J, Cai QH, Xiao W ( 2017) Spatial distribution and significance of high mountain micro-waterbodies in northwestern Yunnan, China. Journal of Hydroecology, 38, 18-23. (in Chinese with English abstract) |
| [ 刘硕然, 和晓阳, 杨文书, 任国鹏, 李延鹏, 周俊, 蔡庆华, 肖文 ( 2017) 滇西北高山微水体空间分布格局及研究意义初探. 水生态学杂志, 38, 18-23.] | |
| [18] | Liu SR, Li YP, Yan JG, Xiao W ( 2018) A Waterbody, Organisms and Sediment Sampling Equipment Used for Different Kinds of Water Environments. Chinese Patent. ZL201510837357.X. 2018-10-19. (in Chinese) |
| [ 刘硕然, 李延鹏, 闫家国, 肖文 ( 2018) 一种适用于不同水体环境的水体、生物、沉积物采集器. 中国专利: ZL201510837357. X. 2018-10-19.] | |
| [19] | Liu SR, Lu T, Yang DD, Ren GP, He XY, Yang WS, Cai QH, Xiao W ( 2018) Spatiotemporal environmental heterogeneity of alpine micro-waterbodies. Fresenius Environmental Bulletin, 27, 8088-8095. |
| [20] | McCune B, Mefford MJ ( 2016) PC-ORD Multivariate Analysis of Ecological Data, Version 7.04. MjM Software, Gleneden Beach, Oregon. |
| [21] | Ministry of Ecology and Environment of the People’s Republic of China ( 2017) HJ 828-2017. Water Quality Determination of Chemical Oxygen Demand: Dichromate Method . China Environmental Press, Beijing. (in Chinese with) |
| [ 中华人民共和国生态环境部 ( 2017) HJ 828-2017 水质化学需氧量的测定: 重铬酸盐法. 中国环境出版社, 北京.] | |
| [22] | Miserendino ML ( 2001) Macroinvertebrate assemblages in Andean Patagonian rivers and streams: Environmental relationships. Hydrobiologia, 444, 147-158. |
| [23] | Myers N ( 1988) Threatened biotas: Hot spots in tropical forests. The Environmentalist, 8, 187-208. |
| [24] | Myers N, Mittermeier RA, Mittermeier CG, Fonseca GA, Kent J ( 2000) Biodiversity hotspots for conservation priorities. Nature, 403, 853-858. |
| [25] | National Environmental Protection Administration ( 1990a) GB 11893-1989. Water Quality Determination of Total Phosphorus: Ammonium Molybdate Spectrophotometric Method. China Standard Press, Beijing. (in Chinese with) |
| [ 国家环境保护总局 ( 1990) GB 11893-1989 水质总磷的测定: 钼酸铵分光光度法. 中国标准出版社, 北京.] | |
| [26] | National Environmental Protection Administration ( 1990b) GB 11894-1989. Water Quality Determination of Total Nitrogen: Alkaline Potassium Persulfate Digestion UV Spectrophotometric Method. China Standard Press, Beijing. (in Chinese with) |
| [ 国家环境保护总局 ( 1990) GB 11894-1989 水质总氮的测定: 碱性过硫酸钾消解紫外分光光度法. 中国标准出版社, 北京.] | |
| [27] | Poff NL, Olden JD, Vieira NK, Finn DS, Simmons MP, Kondratieff BC ( 2006) Functional trait niches of North American lotic insects: Traits-based ecological applications in light of phylogenetic relationships. Journal of the North American Benthological Society, 25, 730-755. |
| [28] | Real M, Rieradevall M, Prat N ( 2000) Chironomus species (Diptera: Chironomidae) in the profundal benthos of Spanish reservoirs and lakes: Factors affecting distribution patterns. Freshwater Biology, 43, 1-18. |
| [29] | Sala OE, Chapin FS, Armesto JJ, Berlow EL, Bloomfield J, Dirzo R, Huber-Sanwald E, Huenneke LF, Jackson RB, Kinzig AP, Leemans R, Lodge DM, Mooney HA, Oesterheld M, Poff NL, Sykes MT, Walker BH, Walker M, Wall DH ( 2000) Global biodiversity scenarios for the year 2100. Science, 287, 1770-1774. |
| [30] | Scheffer M, Van Geest GJ, Zimmer KD, Jeppesen E, Sondergaard M, Butler MG, Hanson MA, Declerck S, De Meester L ( 2006) Small habitat size and isolation can promote species richness: Second-order effects on biodiversity in shallow lakes and ponds. Oikos, 112, 227-231. |
| [31] | Stokstad E ( 2014) EPA science report signals start of wetlands battle. Science, 343, 15. |
| [32] | Taft B, Koncelik JP ( 2006) Methods for Assessing Habitat in Flowing Waters: Using the Qualitative Habitat Evaluation Index (QHEI). Division of Surface Water, Ohio EPA. . (accessed on 2019-05-07) |
| [1] | 张明燡, 王晓梅, 郑言鑫, 吴楠, 李东浩, 樊恩源, 李娜, 单秀娟, 于涛, 赵春暖, 李波, 徐帅, 吴玉萍, 任利群. 黄河口典型牡蛎礁分布区资源状况和栖息地功能[J]. 生物多样性, 2025, 33(4): 24208-. |
| [2] | 仝淼, 王欢, 张文双, 王超, 宋建潇. 重金属污染土壤中细菌抗生素抗性基因分布特征[J]. 生物多样性, 2025, 33(3): 24101-. |
| [3] | 李艳朋, 陈洁, 卢春洋, 许涵. 海南尖峰岭热带山地雨林64 ha次生林动态监测样地群落结构特征[J]. 生物多样性, 2025, 33(2): 24445-. |
| [4] | 陈自宏, 张翼飞, 陈凯, 陈见影, 徐玲. 高黎贡山南段昆虫病原真菌物种多样性及影响因素[J]. 生物多样性, 2025, 33(1): 24228-. |
| [5] | 魏诗雨, 宋天骄, 罗佳宜, 张燕, 赵子萱, 茹靖雯, 易华, 林雁冰. 秦岭火地塘针叶林土壤细菌群落的海拔分布格局[J]. 生物多样性, 2024, 32(9): 24180-. |
| [6] | 时永强, 栾青杉, 单秀娟, 韦超, 赵永松, 孙策策, 金显仕. 长岛南部海域浮游动物多样性周年变化[J]. 生物多样性, 2024, 32(7): 23428-. |
| [7] | 倪艳梅, 陈莉, 董志远, 孙德斌, 李宝泉, 王绪敏, 陈琳琳. 黄河三角洲湿地生态修复区大型底栖动物群落结构与生态健康评价[J]. 生物多样性, 2024, 32(3): 23303-. |
| [8] | 陈瑶琪, 郭晶晶, 蔡国俊, 葛依立, 廖宇, 董正, 符辉. 近七十年(1954-2021)长江中下游湖泊沉水植物群落多样性演变特征[J]. 生物多样性, 2024, 32(3): 23319-. |
| [9] | 魏嘉欣, 姜治国, 杨林森, 熊欢欢, 金胶胶, 罗方林, 李杰华, 吴浩, 徐耀粘, 乔秀娟, 魏新增, 姚辉, 余辉亮, 杨敬元, 江明喜. 湖北神农架中亚热带山地落叶阔叶林25 ha动态监测样地群落物种组成与结构特征[J]. 生物多样性, 2024, 32(3): 23338-. |
| [10] | 刘啸林, 吴友贵, 张敏华, 陈小荣, 朱志成, 陈定云, 董舒, 李步杭, 丁炳扬, 刘宇. 浙江百山祖25 ha亚热带森林动态监测样地群落组成与结构特征[J]. 生物多样性, 2024, 32(2): 23294-. |
| [11] | 吴芳芳, 刘娜, 何春梅, 原作强, 郝占庆, 尹秋龙. 秦岭山地木本植物群落结构及多样性的海拔梯度格局[J]. 生物多样性, 2024, 32(12): 24239-. |
| [12] | 单航, 雷祖培, 郑方东, 韦博良, 仲磊, 于明坚. 2013-2023年浙江乌岩岭次生常绿阔叶林群落动态变化[J]. 生物多样性, 2024, 32(12): 24372-. |
| [13] | 冯嘉谊, 练琚愉, 冯瑜莙, 张东旭, 曹洪麟, 叶万辉. 鼎湖山南亚热带常绿阔叶林群落垂直分层对群落结构及功能的影响[J]. 生物多样性, 2024, 32(12): 24306-. |
| [14] | 王兴煜, 孟京辉, 任思远, 祝燕. 北京东灵山暖温带落叶阔叶林群落生物多样性与地上生物量的关系[J]. 生物多样性, 2024, 32(12): 24230-. |
| [15] | 杜晴晴, 任思远, Nicole Tsz Shun Yuan, 祝燕. 北京东灵山暖温带落叶阔叶林幼树及成树生产力的影响因素[J]. 生物多样性, 2024, 32(12): 24284-. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
备案号:京ICP备16067583号-7
Copyright © 2022 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn