
生物多样性 ›› 2018, Vol. 26 ›› Issue (7): 701-706. DOI: 10.17520/biods.2018144 cstr: 32101.14.biods.2018144
所属专题: 昆虫多样性与生态功能
张宇, 冯刚*( )
)
收稿日期:2018-05-15
接受日期:2018-07-20
出版日期:2018-07-20
发布日期:2018-09-11
通讯作者:
冯刚
作者简介:# 共同第一作者
基金资助:
Yu Zhang, Gang Feng*()
Received:2018-05-15
Accepted:2018-07-20
Online:2018-07-20
Published:2018-09-11
Contact:
Feng Gang
About author:# Co-first authors
摘要:
物种多样性的地理分布格局及其机制是宏生态学和生物地理学的核心问题之一。区域尺度与局域尺度的影响因素, 如温度、降水、海拔变化、生境过滤、捕食、竞争与互惠等, 共同影响昆虫物种多样性的分布格局。然而, 迄今为止少有研究同时讨论不同尺度驱动因子对昆虫多样性地理分布格局的影响。本文基于内蒙古自治区86个旗县的昆虫多样性数据, 结合各地年平均气温、年降水量、古气候变化、海拔变化及植物多样性, 探讨昆虫物种多样性分布格局及其主要驱动因子。结果发现内蒙古昆虫多样性主要受到植物多样性与海拔变化的影响, 而气候因子对昆虫物种多样性的影响并不大。这一结果表明种间关系(食物多样性)与生境异质性可能对内蒙古昆虫多样性的分布格局起着主导作用。
张宇, 冯刚 (2018) 内蒙古昆虫物种多样性分布格局及其机制. 生物多样性, 26, 701-706. DOI: 10.17520/biods.2018144.
Yu Zhang, Gang Feng (2018) Distribution pattern and mechanism of insect species diversity in Inner Mongolia. Biodiversity Science, 26, 701-706. DOI: 10.17520/biods.2018144.
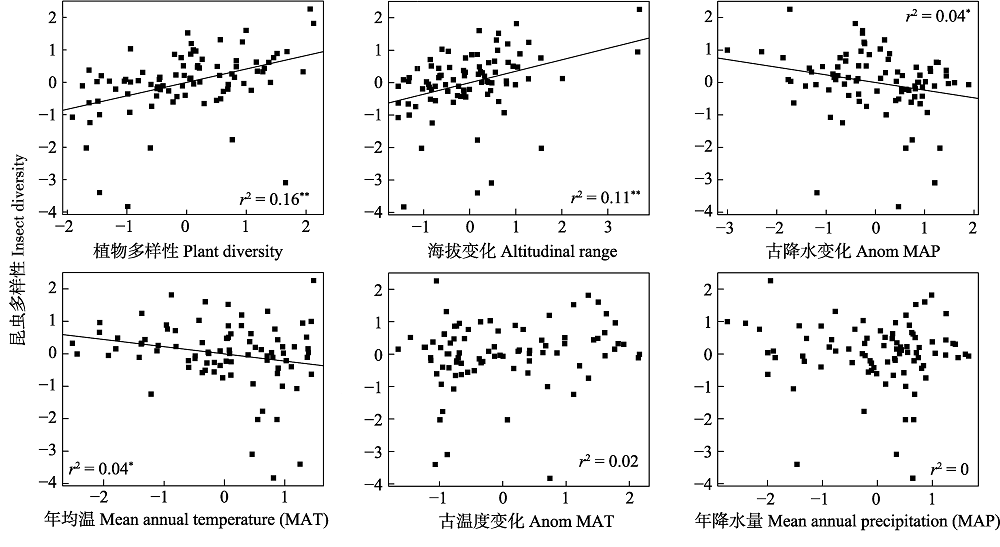
图1 昆虫多样性及其相关变量散点图。古降水变化与古温度变化是末次最大冰期到现代的降水和温度的变化。* P < 0.05, ** P < 0.01。
Fig. 1 Scatter plots of insect species diversity and the related variables. Anom MAP and Anom MAT are the change of MAP/MAT between LGM and contemporary time.
| coefOLS | r2OLS | coefSAR | r2SAR | |
|---|---|---|---|---|
| 年平均气温 Mean annual temperature (MAT) | -0.22 | 0.04* | -0.24 | 0.08** |
| 年降水量 Mean annual precipitation (MAP) | -0.06 | 0 | -0.05 | 0 |
| 古温度变化 Anom MAT | 0.17 | 0.02 | 0.2 | 0.06** |
| 古降水变化 Anom MAP | -0.24 | 0.04* | -0.23 | 0.07* |
| 植物多样性 Plant diversity | 0.35 | 0.16** | 0.36 | 0.13** |
| 海拔变化 Altitudinal range | 0.42 | 0.11** | 0.41 | 0.18** |
表1 最小二乘法(OLS)和空间自回归(SAR)模型单变量分析结果
Table 1 Results of single-variable analysis by ordinary least squares (OLS) and simultaneous autoregressive (SAR) models
| coefOLS | r2OLS | coefSAR | r2SAR | |
|---|---|---|---|---|
| 年平均气温 Mean annual temperature (MAT) | -0.22 | 0.04* | -0.24 | 0.08** |
| 年降水量 Mean annual precipitation (MAP) | -0.06 | 0 | -0.05 | 0 |
| 古温度变化 Anom MAT | 0.17 | 0.02 | 0.2 | 0.06** |
| 古降水变化 Anom MAP | -0.24 | 0.04* | -0.23 | 0.07* |
| 植物多样性 Plant diversity | 0.35 | 0.16** | 0.36 | 0.13** |
| 海拔变化 Altitudinal range | 0.42 | 0.11** | 0.41 | 0.18** |
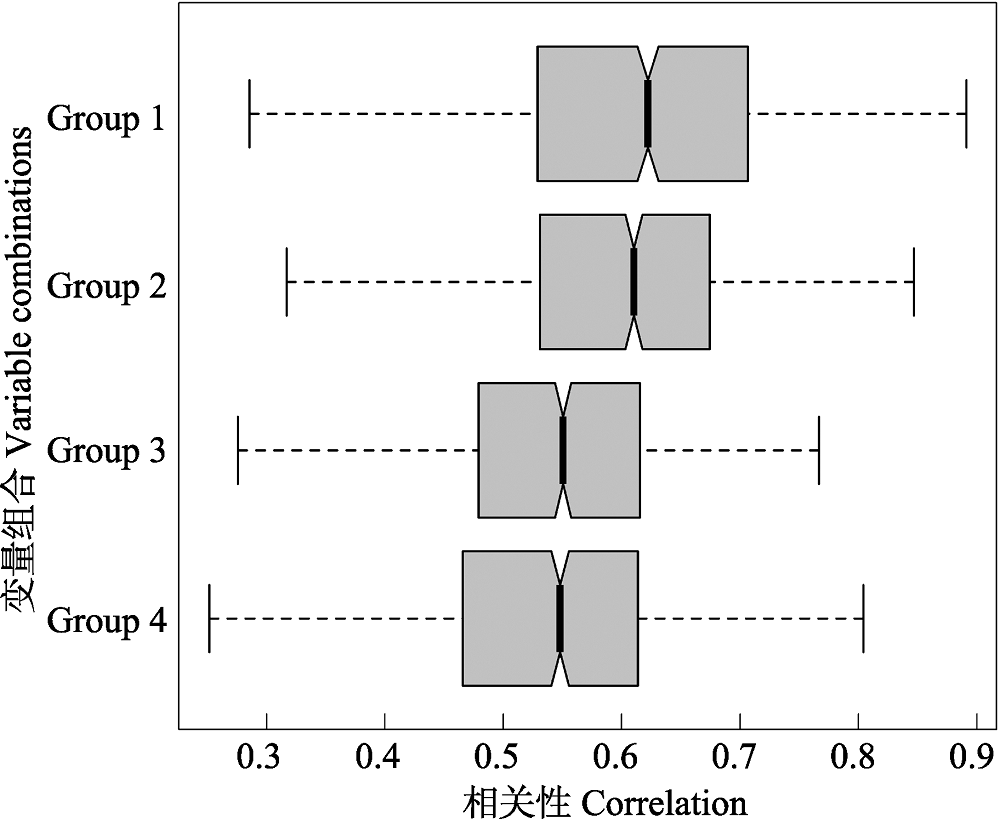
图2 随机森林模型得到的与昆虫物种多样性相关性最高的4个组合。Group 1为Anom MAP + Plant diversity, Group 2为Anom MAP + Altitudinal range + Plant diversity, Group 3为MAT + Anom MAP + Altitudinal range + Plant diversity, Group 4为MAP + Anom MAP + Altitudinal range + Plant diversity。Anom MAP表示古降水变化。
Fig. 2 Four combinations of variables most associated with insect species diversity obtained from the random forest model. Group 1 are Anom MAP and Plant diversity, Group 2 are Anom MAP, Altitudinal range and Plant diversity, Group 3 are MAT, Anom MAP, Altitudinal range and Plant diversity, Group 4 are MAP, Anom MAP, Altitudinal range and Plant diversity. Anom MAP is the change of MAP between LGM and contemporary time.
| [1] | Aranda R, Graciolli G (2015) Spatial temporal distribution of the Hymenoptera in the Brazilian Savanna and the effects of habitat heterogeneity on these patterns. Journal of Insect Conservation, 19, 1173-1187. |
| [2] | Araújo MB, Rozenfeld A (2014) The geographic scaling of biotic interactions. Ecography, 37, 406-415. |
| [3] | Basset Y, Cizek L, Cuénoud P, Didham RK, Guilhaumon F, Missa O, Novotny V, Ødegaard F, Roslin T, Schmidl J, Tishechkin AK, Winchester NN, Roubik DW, Aberlenc HP, Bail J, Barrios H, Bridle JR, Castaño-Meneses G, Corbara B, Curletti G, da Rocha WD, Bakker DD, Delabie JH, Dejean A, Fagan LL, Floren A, Kitching RL, Medianero E, Miller SE, de Orivel EJ, Pollet M, Rapp M, Ribeiro SP, Roisin Y, Schmidt JB, Sørensen L, Leponce M (2012) Arthropod diversity in a tropical forest. Science, 338, 1481-1484. |
| [4] | Boyer SL, Markle TM, Baker CM, Luxbacher AM, Kozak KH (2016) Historical refugia have shaped biogeographical patterns of species richness and phylogenetic diversity in mite harvestmen (Arachnida, Opiliones, Cyphophthalmi) endemic to the Australian Wet Tropics. Journal of Biogeography, 43, 1400-1411. |
| [5] | Breiman L (2001) Random forests. Machine Learning, 45, 5-32. |
| [6] | Brown JH (2014) Why are there so many species in the tropics? Journal of Biogeography, 41, 8-22. |
| [7] | Currie DJ (1991) Energy and large scale patterns of animal and plant species richness. The American Naturalist, 137, 27-49. |
| [8] | Diniz-Filho JAF, Marco PD, Hawkins BA (2010) Defying the curse of ignorance: Perspectives in insect macroecology and conservation biogeography. Insect Conservation and Diversity, 3, 172-179. |
| [9] | Fine P (2015) Ecological and evolutionary drivers of geographic variation in species diversity. Annual Review of Ecology, Evolution, and Systematics, 46, 369-392. |
| [10] | Gebeyehu S, Samways MJ (2006) Topographic heterogeneity plays a crucial role for grasshopper diversity in a Southern African megabiodiversity hotspot. Biodiversity and Conservation, 15, 231-244. |
| [11] | Harrison S, Cornell H (2008) Toward a better understanding of the regional causes of local community richness. Ecology Letters, 11, 969-979. |
| [12] | Hasumi H, Emori S (2004) K-1 Coupled Model (MIROC) description. K-1 Technical Report No.1. Center for Climate System Research, University of Tokyo, Tokyo. |
| [13] | Hawkins BA (2001) Ecology’s oldest pattern? Trends in Ecology and Evolution, 16, 470. |
| [14] | Hijmans RJ, Cameron SE, Parra JL, Jones PG, Jarvis A (2005) Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology, 25, 1965-1978. |
| [15] | Liang CX, Feng G, Si XF, Mao LF, Yang GS, Svenning J-C, Yang J (2018) Bird species richness is associated with phylogenetic relatedness, plant species richness, and altitudinal range in Inner Mongolia. Ecology and Evolution, 8, 53-58. |
| [16] | Marini L, Fontana P, Battisti A, Gaston KJ (2010) Agricultural management, vegetation traits and landscape drive orthopteran and butterfly diversity in a grassland-forest mosaic: A multi-scale approach. Insect Conservation and Diversity, 2, 213-220. |
| [17] | Marini L, Paolo F, Sebastian K, Andrea B, Kevin G (2009) Impact of farm size and topography on plant and insect diversity of managed grasslands in the Alps. Biological Conservation, 142, 394-403. |
| [18] | Misof B, Liu SL, Meusemann K, Peters RS, Donath A, Mayer C, Frandsen PB, Ware J, Flouri T, Beutel RJ, Niehuis O, Petersen M, Izquierdo-Carrasco F, Wappler T, Rust J, Aberer AJ, Aspöck U, Aspöck H, Bartel D, Blanke A, Berger S, Böhm A, Buckley TR, Calcott B, Chen JQ, Friedrich F, Fukui M, Fujita M, Greve C, Grobe P, Gu SC, Huang Y, Jermiin LS, Kawahara AY, Krogmann L, Kubiak M, Lanfear R, Letsch H, Li YY, Li ZY, Li JG, Lu HR, Machida R, Mashimo Y, Kapli P, McKenna DD, Meng GL, Nakagaki Y, Navarrete-Heredia JL, Ott M, Ou YX, Pass G, Podsiadlowski L, Poh H, von Reumont BM, Schütte K, Sekiya K, Shimizu S, Slipinski A, Stamatakis A, Song WH, Su X, Szucsich NU, Tan MH, Tan XM, Tang M, Tang JB, Timelthaler G, Tomizuka S, Trautwein M, Tong XL, Uchifune T, Walzl MG, Wiegmann BM, Wilbrandt J, Wipfler B, Wong TKF, Wu Q, Wu GX, Xie YL, Yang SZ, Yang Q, Yeates DK, Yoshizawa K, Zhang Q, Zhang R, Zhang WW, Zhang YH, Zhao J, Zhou CR, Zhou LL, Ziesmann T, Zou SJ, Li YR, Xu X, Zhang Y, Yang HM, Wang J, Wang J, Kjer KM, Zhou X (2014) Phylogenomics resolves the timing and pattern of insect evolution. Science, 346, 763-767. |
| [19] | Nonnaizab(1999) Insects of Inner Mongolia, China. Inner Mongolia People’s Publishing House, Hohhot. (in Chinese) |
| [能乃扎布 (1999) 内蒙古昆虫. 内蒙古人民出版社, 呼和浩特.] | |
| [20] | O’Brien EM (1998) Water-energy dynamics, climate, and prediction of woody plant species richness: An interim general model. Journal of Biogeography, 25, 379-398. |
| [21] | O’Brien EM, Field R, Whittaker RJ (2000) Climatic gradients in woody plant (tree and shrub) diversity: water-energy dynamics, residual variation, and topography. Oikos, 89, 588-600. |
| [22] | Otto-Bliesner B, Brady E, Clauzet G, Thomas R, Levis S, Kothavala Z (2006) Last Glacial Maximum and Holocene Climate in CCSM3. Journal of Climate, 19, 2526-2544. |
| [23] | Price PW (1991) The plant vigor hypothesis and herbivore attack. Oikos, 62, 244-251. |
| [24] | Qu YH, Song G, Gao B, Quan Q, Ericson PGP, Lei FM (2015) The influence of geological events on the endemism of East Asian birds studied through comparative phylogeography. Journal of Biogeography, 42, 179-192. |
| [25] | Sandel B, Arge L, Dalsgaard B, Davies RG, Gaston KJ, Sutherland WJ, Svenning J-C (2011) The influence of Late Quaternary climate change velocity on species endemism. Science, 334, 660-664. |
| [26] | Schuldt A, Baruffol M, Böhnke M, Bruelheide H, Hädtle W, Lang AC, Nadrowski K, von Oheimb G, Voigt W, Zhou HZ, Assmann T (2010) Tree diversity promotes insect herbivory in subtropical forests of south-east China. Journal of Ecology, 98, 917-926. |
| [27] | Shen MW, Chen SB, Bi MJ, Chen WD, Zhou KX (2016) Relationships between geographic patterns of ant species richness and environmental factors in China. Acta Ecologica Sinica, 36, 7732-7739. (in Chinese with English abstract) |
| [沈梦伟, 陈圣宾, 毕孟杰, 陈文德, 周可新 (2016) 中国蚂蚁丰富度地理分布格局及其与环境因子的关系. 生态学报, 36, 7732-7739.] | |
| [28] | Stein A, Gerstner K, Kreft H (2014) Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecology Letters, 17, 866-880. |
| [29] | Svenning J-C, Eiserhardt WL, Normand S, Ordonez A, Sandel B (2015) The influence of Paleoclimate on present day patterns in biodiversity and ecosystems. Annual Review of Ecology, Evolution, and Systematics, 46, 551-572. |
| [30] | Tews J, Brose U, Grimm V, Tielbörger K, Wichmann MC, Schwager M, Jeltsch F (2004) Animal species diversity driven by habitat heterogeneity/diversity: The importance of keystone structures. Journal of Biogeography, 31, 79-92. |
| [31] | Thomas CD, Bulman CR, Wilson RJ (2008) Where within a geographical range do species survive best? A matter of scale. Insect Conservation and Diversity, 1, 2-8. |
| [32] | Wenninger EJ, Inouye RS (2008) Insect community response to plant diversity and productivity in a sagebrush steppe ecosystem. Journal of Arid Environments, 72, 24-33. |
| [33] | Wisz MS, Pottier J, Kissling WD, Pellissier L, Lenoir J, Damgaard CF, Dormann CF, Forchhammer MC, Grytnes J-A, Guisan A, Heikkinen RK, Høye TT, Kühn I, Luoto M, Maiorano L, Nilsson M-C, Normand S, Öckinger E, Schmidt NM, Termansen M, Timmermann A, Wardle DA, Aastrup P, Svenning J-C (2013) The role of biotic interactions in shaping distributions and realised assemblages of species: Implications for species distribution modelling. Biological Reviews, 88, 15-30. |
| [34] | Xu RG (2007) Fauna Inner Mongolia. III. Inner Mongolia University Press, Hohhot. (in Chinese) |
| [旭日干 (2007) 内蒙古动物志. III. 内蒙古大学出版社, 呼和浩特.] | |
| [35] | Xu RG (2015)Fauna Inner Mongolia. IV. Inner Mongolia University Press, Hohhot. (in Chinese) |
| [旭日干 (2015) 内蒙古动物志. IV. 内蒙古大学出版社, 呼和浩特.] | |
| [36] | Xu RG (2016a) Fauna Inner Mongolia. V. Inner Mongolia University Press, Hohhot. (in Chinese) |
| [旭日干 (2016a) 内蒙古动物志. V. 内蒙古大学出版社, 呼和浩特.] | |
| [37] | Xu RG (2016b) Fauna Inner Mongolia. VI. Inner Mongolia University Press, Hohhot. (in Chinese) |
| [旭日干 (2016b) 内蒙古动物志. VI. 内蒙古大学出版社, 呼和浩特.] | |
| [38] | Zhang K, Lin SL, Ji YQ, Yang CX, Wang XY, Yang CY, Wang HS, Jiang HD, Harrison RD, Yu DW (2016) Plant diversity accurately predicts insect diversity in two tropical landscapes. Molecular Ecology, 25, 4407-4419. |
| [39] | Zhao YZ, Zhao LQ (2014) Key to the Vascular Plants of Inner Mongolia. Science Press, Beijing. (in Chinese) |
| [赵一之, 赵利清 (2014) 内蒙古维管植物检索表. 科学出版社, 北京.] | |
| [40] | Zhu H, Peng YY, Wang DL (2008) Effects of plant on insect diversity: A review. Chinese Journal of Ecology, 27, 2215-2221. |
| (in Chinese with English abstract) [朱慧, 彭媛媛, 王德利 (2008) 植物对昆虫多样性的影响. 生态学杂志, 27, 2215-2221.] | |
| [41] | Zobel M (1997) The relative role of species pools in determining plant species richness: An alternative explanation of species coexistence? Trends in Ecology and Evolution, 12, 266-269. |
| [1] | 张浩斌, 肖路, 刘艳杰. 夜间灯光对外来入侵植物和本地植物群落多样性和生长的影响[J]. 生物多样性, 2025, 33(4): 24553-. |
| [2] | 宋威, 程才, 王嘉伟, 吴纪华. 土壤微生物对植物多样性–生态系统功能关系的调控作用[J]. 生物多样性, 2025, 33(4): 24579-. |
| [3] | 陈楠, 张全国. 实验进化研究途径[J]. 生物多样性, 2024, 32(9): 24171-. |
| [4] | 牛红玉, 陈璐, 赵恒月, 古丽扎尔·阿不都克力木, 张洪茂. 城市化对动物的影响: 从群落到个体[J]. 生物多样性, 2024, 32(8): 23489-. |
| [5] | 连佳丽, 陈婧, 杨雪琴, 赵莹, 罗叙, 韩翠, 赵雅欣, 李建平. 荒漠草原植物多样性和微生物多样性对降水变化的响应[J]. 生物多样性, 2024, 32(6): 24044-. |
| [6] | 万凤鸣, 万华伟, 张志如, 高吉喜, 孙晨曦, 王永财. 草地植物多样性无人机调查的应用潜力[J]. 生物多样性, 2024, 32(3): 23381-. |
| [7] | 黄小龙, 蒙秉顺, 李海波, 冉伟, 杨伟, 王丞, 谢波, 张旭, 冉景丞, 张明明. 基于红外相机的黔金丝猴及其同域分布物种种间关联[J]. 生物多样性, 2024, 32(2): 23402-. |
| [8] | 张乃鹏, 梁洪儒, 张焱, 孙超, 陈勇, 王路路, 夏江宝, 高芳磊. 土壤类型和地下水埋深对黄河三角洲典型盐沼植物群落空间分异的影响[J]. 生物多样性, 2024, 32(2): 23370-. |
| [9] | 蒋陈焜, 郁文彬, 饶广远, 黎怀成, Julien B. Bachelier, Hartmut H. Hilger, Theodor C. H. Cole. 植物系统发生海报——以演化视角介绍植物多样性的科教资料项目[J]. 生物多样性, 2024, 32(11): 24210-. |
| [10] | 杨舒涵, 王贺, 陈磊, 廖蓥飞, 严光, 伍一宁, 邹红菲. 松嫩平原异质生境对土壤线虫群落特征的影响[J]. 生物多样性, 2024, 32(1): 23295-. |
| [11] | 陈又生, 宋柱秋, 卫然, 罗艳, 陈文俐, 杨福生, 高连明, 徐源, 张卓欣, 付鹏程, 向春雷, 王焕冲, 郝加琛, 孟世勇, 吴磊, 李波, 于胜祥, 张树仁, 何理, 郭信强, 王文广, 童毅华, 高乞, 费文群, 曾佑派, 白琳, 金梓超, 钟星杰, 张步云, 杜思怡. 西藏维管植物多样性编目和分布数据集[J]. 生物多样性, 2023, 31(9): 23188-. |
| [12] | 宋柱秋, 叶文, 董仕勇, 金梓超, 钟星杰, 王震, 张步云, 徐晔春, 陈文俐, 李世晋, 姚纲, 徐洲锋, 廖帅, 童毅华, 曾佑派, 曾云保, 陈又生. 广东省高等植物多样性编目和分布数据集[J]. 生物多样性, 2023, 31(9): 23177-. |
| [13] | 王明慧, 陈昭铨, 李帅锋, 黄小波, 郎学东, 胡子涵, 尚瑞广, 刘万德. 云南普洱季风常绿阔叶林不同种子扩散方式的优势种空间点格局分析[J]. 生物多样性, 2023, 31(9): 23147-. |
| [14] | 韩赟, 迟晓峰, 余静雅, 丁旭洁, 陈世龙, 张发起. 青海野生维管植物名录[J]. 生物多样性, 2023, 31(9): 23280-. |
| [15] | 梁彩群, 陈玉凯, 杨小波, 张凯, 李东海, 江悦馨, 李婧涵, 王重阳, 张顺卫, 朱子丞. 海南省野生维管植物编目和分布数据集[J]. 生物多样性, 2023, 31(6): 23067-. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
备案号:京ICP备16067583号-7
Copyright © 2022 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn