
生物多样性 ›› 2011, Vol. 19 ›› Issue (4): 414-423. DOI: 10.3724/SP.J.1003.2011.10289 cstr: 32101.14.SP.J.1003.2011.10289
陈圣宾1, 欧阳志云1,*( ), 方瑜1, 李振基2
), 方瑜1, 李振基2
收稿日期:2010-12-03
接受日期:2011-04-20
出版日期:2011-07-20
发布日期:2011-07-29
通讯作者:
欧阳志云
作者简介:* E-mail: zyouyang@rcees.ac.cn基金资助:
Shengbin Chen1, Zhiyun Ouyang1,*(), Yu Fang1, Zhenji Li2
Received:2010-12-03
Accepted:2011-04-20
Online:2011-07-20
Published:2011-07-29
Contact:
Zhiyun Ouyang
摘要:
生物特有现象的地理格局及其形成机制是生物地理学的重要研究内容。本文通过整合173个地区的中国种子植物特有属编目资料、环境和空间因子数据, 运用多元回归和方差分解的方法, 探索了中国种子植物特有属丰富度及其占全部种子植物属丰富度的比例(特有属比例)与环境(生境异质性和气候)和空间因子的关系。结果表明: (1)特有属丰富度及特有属比例具有很强的空间变异性, 在华中地区最高, 而靠近国界和大陆边缘的地区较低; 相比而言, 种子植物属丰富度的空间变异性较弱, 且表现出显著的纬度梯度性; (2)特有属丰富度及特有属比例主要由空间因子和生境异质性(地形的复杂性)决定, 即在大的空间尺度上, 地理位置决定一个地区特有属比例的理论值, 生境异质性和气候因子对其进行微调; 而种子植物属丰富度的地理格局主要受气候和生境异质性的影响。(3)中国种子植物特有属是主观性非常强的概念, 特有属比例所反映的植物区系系统发育信息可能会很低; 空间因子所解释的方差中到底有多少是系统发育因素, 还需要进一步的研究。本文最后讨论了当前特有属定义和判定的不足之处。虽然理论上认为特有属的判定不应以行政边界为标准, 但是目前几乎所有的中国特有属划分方法均以国界为准, 这在一定程度上降低了中国种子植物特有属概念的科学内涵和在实践中的作用。因此,我们建议在理论和实践中对中国种子植物特有属概念采取审慎的态度。
陈圣宾, 欧阳志云, 方瑜, 李振基 (2011) 中国种子植物特有属的地理分布格局. 生物多样性, 19, 414-423. DOI: 10.3724/SP.J.1003.2011.10289.
Shengbin Chen, Zhiyun Ouyang, Yu Fang, Zhenji Li (2011) Geographic patterns of endemic seed plant genera diversity in China. Biodiversity Science, 19, 414-423. DOI: 10.3724/SP.J.1003.2011.10289.
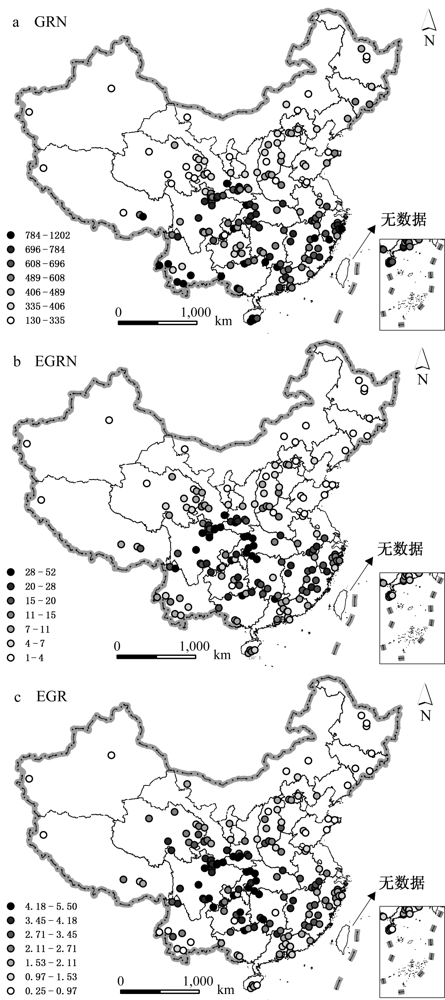
图1 中国种子植物属丰富度(GRN)、特有属丰富度(EGRN)和特有属比例(EGR)的地理分布格局。
Fig. 1 Geographic patterns of genera richness (GRN), endemic genera richness (EGRN) and endemic genera ratio (EGR) for seed plants.
| 属丰富度 GRN | 特有属丰富度 EGRN | |
|---|---|---|
| 特有属丰富度 EGRN | 0.703* | |
| 特有属比例 EGR | 0.311* | 0.852* |
表1 中国种子植物属丰富度(GRN)、特有属丰富度(EGRN)和特有属比例(EGR)之间的相关性(丰富度数据均经过自然对数转换)
Table 1 Correlation between genera richness (GRN), endemic genera richness (EGRN), and endemic genera ratio (EGR) of seed plants in China. Richness data were ln-transformed.
| 属丰富度 GRN | 特有属丰富度 EGRN | |
|---|---|---|
| 特有属丰富度 EGRN | 0.703* | |
| 特有属比例 EGR | 0.311* | 0.852* |
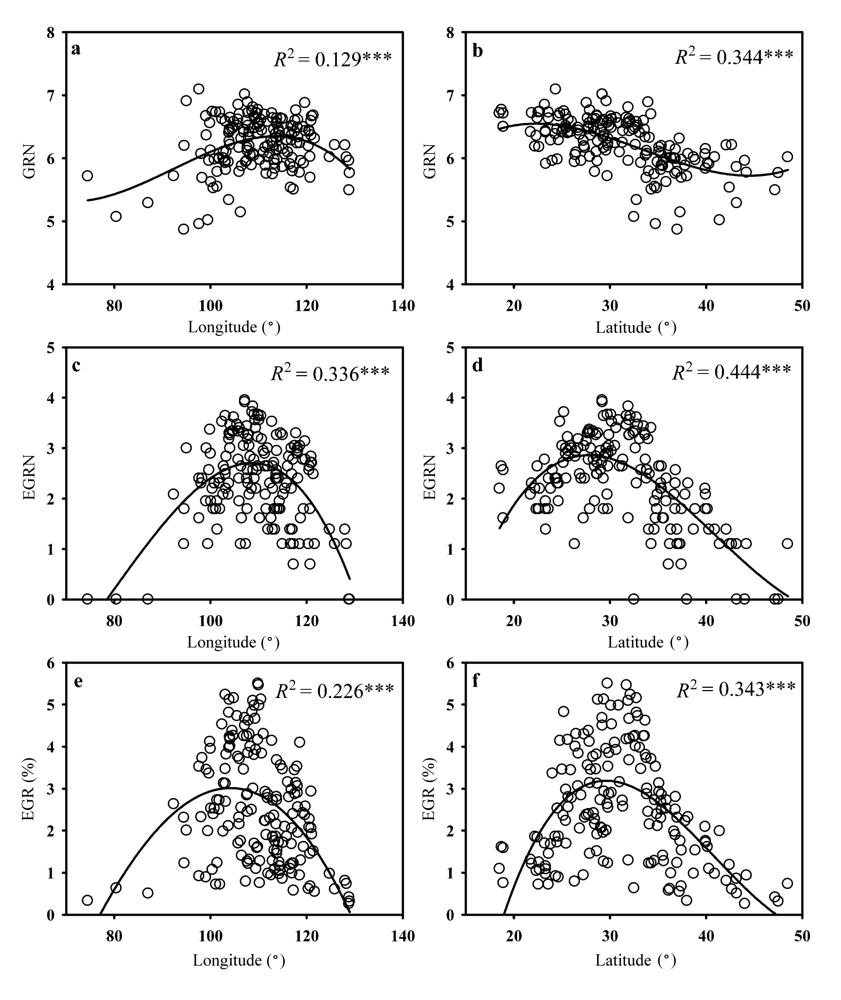
图2 种子植物属丰富度(GRN) (a, b)、特有属丰富度(EGRN) (c, d)和特有属比例(EGR) (e, f)与经纬度的关系, 丰富度数据均经过自然对数转换。
Fig. 2 Changes of genera richness (GRN) (a, b), endemic genera richness (EGRN) (c, d), and endemic genera ratio (EGR) (e, f) of seed plants along with longitude and latitude. Richness data were ln-transformed.
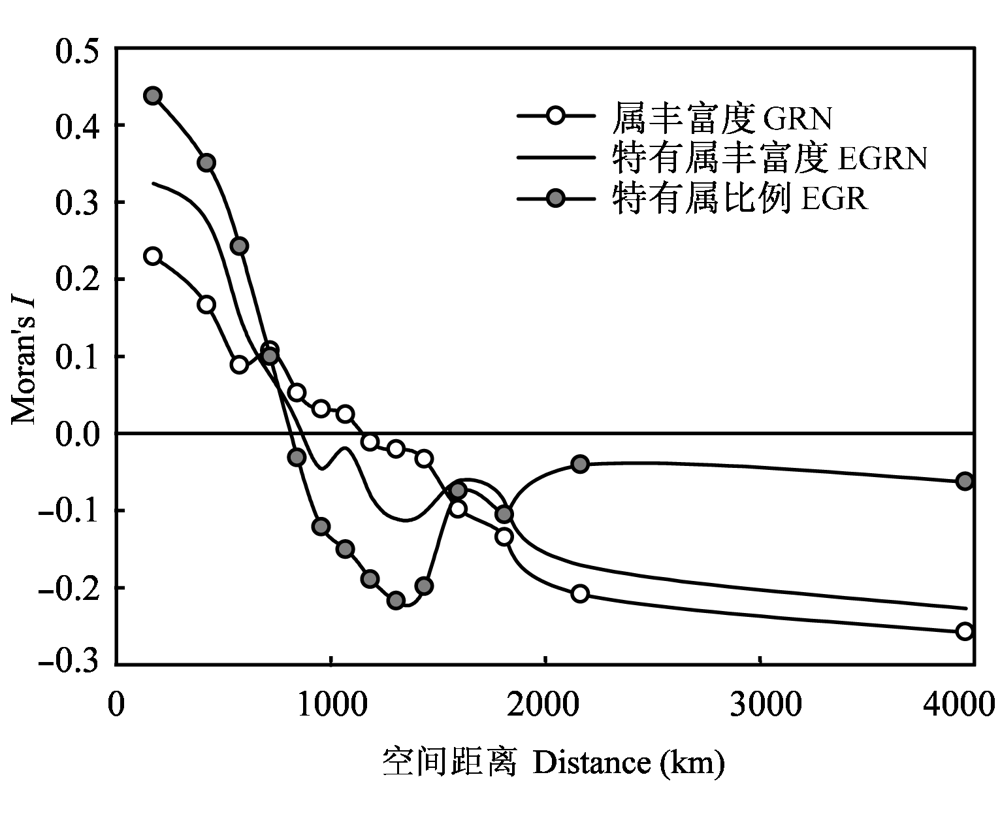
图3 不同空间距离上中国种子植物属丰富度(GRN)、特有属丰富度(EGRN)和特有属比例(EGR)的空间自相关特征
Fig. 3 Moran’s I of genera richness (GRN), endemic genera richness (EGRN), and endemic genera ratio (EGR) of seed plants at different distance classes
| 自变量 Independent variables | 属丰富度 GRN | 特有属丰富度 EGRN | 特有属比例 EGR | |||||
|---|---|---|---|---|---|---|---|---|
| 一次 Linear | 二次 Quadratic | 一次 Linear | 二次 Quadratic | 一次 Linear | 二次 Quadratic | |||
| 面积 AREA (–) | 0.051* | 0 0.008 | 0.005 | |||||
| 海拔幅度 REL (+) | 0.004 | 0.050* | 0.067* | 0.084* | 0.106* | 0.102* | ||
| 年均温 TEM (+) | 0.304* | 0.304* | 0.100* | 0.192* | 0.001 | 0.130* | ||
| 最冷月均温 TCM (+) | 0.377* | 0.376* | 0.189* | 0.353* | 0.010 | 0.220* | ||
| 年降雨量 PRE (+) | 0.389* | 0.506* | 0.148* | 0.308* | 0.001 | 0.144* | ||
| 最干月降雨量 PDM (+) | 0.210* | 0.282* | 0.108* | 0.115* | 0.005 | 0.006 | ||
| 潜在蒸散 PET (+) | 0.342* | 0.380* | 0.075* | 0.214* | 0.002 | 0.126* | ||
| 实际蒸散 AET (+) | 0.465* | 0.520* | 0.198* | 0.309* | 0.008 | 0.116* | ||
表2 种子植物属丰富度(GRN)、特有属丰富度(EGRN)和特有属比例(EGR)与各个环境因子的回归系数
Table 2 Regression coefficient of genera richness (GRN), endemic genera richness (EGRN) and endemic genera ratio (EGR) of seed plants with each environmental factor
| 自变量 Independent variables | 属丰富度 GRN | 特有属丰富度 EGRN | 特有属比例 EGR | |||||
|---|---|---|---|---|---|---|---|---|
| 一次 Linear | 二次 Quadratic | 一次 Linear | 二次 Quadratic | 一次 Linear | 二次 Quadratic | |||
| 面积 AREA (–) | 0.051* | 0 0.008 | 0.005 | |||||
| 海拔幅度 REL (+) | 0.004 | 0.050* | 0.067* | 0.084* | 0.106* | 0.102* | ||
| 年均温 TEM (+) | 0.304* | 0.304* | 0.100* | 0.192* | 0.001 | 0.130* | ||
| 最冷月均温 TCM (+) | 0.377* | 0.376* | 0.189* | 0.353* | 0.010 | 0.220* | ||
| 年降雨量 PRE (+) | 0.389* | 0.506* | 0.148* | 0.308* | 0.001 | 0.144* | ||
| 最干月降雨量 PDM (+) | 0.210* | 0.282* | 0.108* | 0.115* | 0.005 | 0.006 | ||
| 潜在蒸散 PET (+) | 0.342* | 0.380* | 0.075* | 0.214* | 0.002 | 0.126* | ||
| 实际蒸散 AET (+) | 0.465* | 0.520* | 0.198* | 0.309* | 0.008 | 0.116* | ||
| 模型 Model | 自变量 Independent variables | R2 (%) | AIC | P |
|---|---|---|---|---|
| 属丰富度(GRN) | ||||
| 生境异质性模型 Heterogeneity model | AREA, REL, REL2 | 9.1 | 178 | < 0.001 |
| 气候模型 Climatic model | PRE, PRE2, PDM, PDM2, PET, TCM, TEM2 | 57.4 | 52 | < 0.001 |
| 空间模型 Spatial model | X, XY, Y2, X3, X2Y, XY2 | 46.6 | 96 | < 0.001 |
| 综合模型 Integrative model | AREA, REL2, PRE, PRE2, PET, X, XY, XY2 | 66.7 | 35 | < 0.001 |
| 特有属丰富(EGRN) | ||||
| 生境异质性模型 Heterogeneity model | AREA, REL, REL2 | 11.9 | 430 | < 0.001 |
| 气候模型 Climatic model | TEM, TEM2, TCM, PRE, PRE2, PDM, PDM2, PET, AET | 60.1 | 305 | < 0.001 |
| 空间模型 Spatial model | X, Y, XY, Y2, X3, X2Y, XY2, Y3 | 58.0 | 306 | < 0.001 |
| 综合模型 Integrative model | AREA, REL2, PRE, PRE2, X, Y, Y2, X2Y, XY2 | 73.9 | 240 | < 0.001 |
| 特有属比例(EGR) | ||||
| 生境异质性模型 Heterogeneity model | AREA, REL, REL2 | 12.2 | 578 | < 0.001 |
| 气候模型 Climatic model | TEM, TEM2, TCM, PET, PRE, PRE2 | 49.1 | 490 | < 0.001 |
| 空间模型 Spatial model | X, Y, X2, XY, Y2, X2Y, XY2, Y3 | 53.5 | 471 | < 0.001 |
| 综合模型 Integrative model | REL, REL2, TEM, TCM, PET, Y, X2, XY, X2Y, XY2, Y3 | 62.2 | 229 | < 0.001 |
表3 种子植物属丰富度(GRN)、特有属丰富度(EGRN)和特有属比例(EGR)的生境异质性、气候、空间和综合模型
Table 3 Summary of heterogeneity, climate, spatial and integrative models for genera richness (GRN), endemic genera richness (EGRN) and endemic genera ratio (EGR) of seed plants
| 模型 Model | 自变量 Independent variables | R2 (%) | AIC | P |
|---|---|---|---|---|
| 属丰富度(GRN) | ||||
| 生境异质性模型 Heterogeneity model | AREA, REL, REL2 | 9.1 | 178 | < 0.001 |
| 气候模型 Climatic model | PRE, PRE2, PDM, PDM2, PET, TCM, TEM2 | 57.4 | 52 | < 0.001 |
| 空间模型 Spatial model | X, XY, Y2, X3, X2Y, XY2 | 46.6 | 96 | < 0.001 |
| 综合模型 Integrative model | AREA, REL2, PRE, PRE2, PET, X, XY, XY2 | 66.7 | 35 | < 0.001 |
| 特有属丰富(EGRN) | ||||
| 生境异质性模型 Heterogeneity model | AREA, REL, REL2 | 11.9 | 430 | < 0.001 |
| 气候模型 Climatic model | TEM, TEM2, TCM, PRE, PRE2, PDM, PDM2, PET, AET | 60.1 | 305 | < 0.001 |
| 空间模型 Spatial model | X, Y, XY, Y2, X3, X2Y, XY2, Y3 | 58.0 | 306 | < 0.001 |
| 综合模型 Integrative model | AREA, REL2, PRE, PRE2, X, Y, Y2, X2Y, XY2 | 73.9 | 240 | < 0.001 |
| 特有属比例(EGR) | ||||
| 生境异质性模型 Heterogeneity model | AREA, REL, REL2 | 12.2 | 578 | < 0.001 |
| 气候模型 Climatic model | TEM, TEM2, TCM, PET, PRE, PRE2 | 49.1 | 490 | < 0.001 |
| 空间模型 Spatial model | X, Y, X2, XY, Y2, X2Y, XY2, Y3 | 53.5 | 471 | < 0.001 |
| 综合模型 Integrative model | REL, REL2, TEM, TCM, PET, Y, X2, XY, X2Y, XY2, Y3 | 62.2 | 229 | < 0.001 |
| 因变量 Response variables | 独立效应 Pure components (%) | 交互效应 Shared components (%) | 解释的总方差 Total variance explained (%) | |||||
|---|---|---|---|---|---|---|---|---|
| H | C | S | HC | HS | CS | HCS | ||
| 属丰富度 GRN | 8.2 | 9.1 | 2.4 | -6.3 | 1.2 | 36.9 | -7.1 | 66.7 |
| 特有属丰富度 EGRN | 7.5 | 5.7 | 8.9 | 1.7 | 5.2 | 41.8 | 2.0 | 73.9 |
| 特有属比例 EGR | 2.5 | 1.9 | 9.8 | 6.8 | 6.1 | 35.9 | 3.2 | 62.2 |
表4 生境异质性(H)、气候因子(C)和空间因子(S)的独立效应及其交互效应(HC、HS、CS、HCS)所能解释的种子植物属丰富度(GRN)、特有属丰富度(EGRN)和特有属比例(EGR)的方差(%)
Table 4 Spatial variance in genera richness (GRN), endemic genera richness (EGRN), and endemic genera ratio (EGR) of seed plants explained exclusively by heterogeneity (H), climatic factors (C) and spatial factors (S), as well as jointly by the interactions between these groups (HC, HS, CS and HCS)
| 因变量 Response variables | 独立效应 Pure components (%) | 交互效应 Shared components (%) | 解释的总方差 Total variance explained (%) | |||||
|---|---|---|---|---|---|---|---|---|
| H | C | S | HC | HS | CS | HCS | ||
| 属丰富度 GRN | 8.2 | 9.1 | 2.4 | -6.3 | 1.2 | 36.9 | -7.1 | 66.7 |
| 特有属丰富度 EGRN | 7.5 | 5.7 | 8.9 | 1.7 | 5.2 | 41.8 | 2.0 | 73.9 |
| 特有属比例 EGR | 2.5 | 1.9 | 9.8 | 6.8 | 6.1 | 35.9 | 3.2 | 62.2 |
| [1] | Akaike H (1974) A new look at the statistical model identification. IEEE Transactions on Automatic Control, 19, 716-723. |
| [2] | Anderson S (1994) Area and endemism. Quarterly Review of Biology, 69, 451-471. |
| [3] | Borcard D, Legendre P, Drapeau P (1992) Partialling out the spatial component of ecological variation. Ecology, 73, 1045-1055. |
| [4] | Casazza G, Zappa E, Mariotti MG, Médail F, Minuto L (2008) Ecological and historical factors affecting distribution pattern and richness of endemic plant species: the case of the Maritime and Ligurian Alps hotspot. Diversity and Distributions, 14, 47-58. |
| [5] | Chang H (张宏达) (1997) Plant endemism and biodiversity. Ecologic Science (生态科学), 16, 9-17. (in Chinese with English abstract) |
| [6] | Chen SB, Jiang GM, Ouyang ZY, Xu WH, Xiao Y (2011) Relative importance of water, energy, and heterogeneity in determining regional pteridophyte and seed plant richness in China. Journal of Systematics and Evolution, 49, 95-107. |
| [7] | Diniz-Filho JAF, Bini LM, Hawkins BA (2003) Spatial autocorrelation and red herrings in geographical ecology. Global Ecology and Biogeography, 12, 53-64. |
| [8] | Dutilleul P (1993) Modifying the t test for assessing the correlation between two spatial processes. Biometrics, 49, 305-314. |
| [9] | Feng JM (冯建孟), Zhu YY (朱有勇) (2010) On the genera of seed plants endemic to China in Yunnan. Ecology and Environmental Sciences (生态环境学报), 19, 621-625. (in Chinese with English abstract) |
| [10] | Hao RM (郝日明) (1997) On the areal-types of the Chinese endemic genera of seed plants. Acta Phytotaxonomica Sinica (植物分类学报), 35, 500-510. (in Chinese with English abstract) |
| [11] | Huang JH, Chen JH, Ying JS, Ma KP (2011) Features and distribution patterns of Chinese endemic seed plant species. Journal of Systematics and Evolution, 49, 81-94. |
| [12] | Lamoreux JF, Morrison JC, Ricketts TH, Olson DM, Dinerstein E, McKnight MW, Shugart HH (2006) Global tests of biodiversity concordance and the importance of endemism. Nature, 440, 212-214. |
| [13] | Legendre P, Legendre L (1998) Numerical Ecology. Elsevier Science, Amsterdam. |
| [14] | Lei FM (雷富民), Tang QQ (唐芊芊), An SC (安书成) (2004) Species differentiation and distribution of endemic genera of birds in China. Journal of Shaanxi Normal University (陕西师范大学学报), 32, 104-114. (in Chinese with English abstract) |
| [15] | Li XW (李锡文) (1994) Two big biodiversity centres of Chinese endemic genera of seed plants and their characteristics in Yunnan Province. Acta Botanica Yunnanica (云南植物研究), 16, 209-220. (in Chinese with English abstract) |
| [16] | Ma J (马骥), Deng HZ (邓虹珠), Chao Z (晁志), Liu CM (刘传明), Zhang HW (张宏伟) (2004) Medicinal plant resources of the Chinese endemic genera of seed plants. China Journal of Chinese Materia Medica (中国中药杂志), 29, 123-129. (in Chinese with English abstract) |
| [17] | Mac Nally R (2000) Regression and model-building in conservation biology, biogeography and ecology: the distinction between―and reconciliation of―‘predictive’ and ‘explanatory’ models. Biodiversity and Conservation, 9, 655-671. |
| [18] | Moran PAP (1950) Notes on continuous stochastic phenomena. Biometrika, 37, 17-23. |
| [19] | Rangel TFLVB, Diniz-Filho JAF, Bini LM (2006) Towards an integrated computational tool for spatial analysis in macroecology and biogeography. Global Ecology and Biogeography, 15, 321-327. |
| [20] | Shen ZH (沈泽昊), Zhang XS (张新时) (2000) A quantitative analysis on the floristic elements of the Chinese subtropical region and their spatial patterns. Acta Phytotaxonomica Sinica (植物分类学报), 38, 366-380. (in Chinese with English abstract) |
| [21] | Wang HS (王荷生) (1989) A study on the origin of spermatophytic genera endemic to China. Acta Botanica Yunnanica (云南植物研究), 11, 1-16. (in Chinese with English abstract) |
| [22] | Wang HS (王荷生), Zhang YL (张镱锂) (1994a) The distribution patterns of spermatophytic families and genera endemic to China. Acta Geographica Sinica (地理学报), 49, 403-417. (in Chinese with English abstract) |
| [23] | Wang HS (王荷生), Zhang YL (张镱锂) (1994b) The biodiversity and characters of spermatophytic genera endemic to China. Acta Botanica Yunnanica (云南植物研究), 16, 209-220. (in Chinese with English abstract) |
| [24] | Willig MR, Kaufman DM, Stevens RD (2003) Latitudinal gradients of biodiversity: pattern, process, scale, and synthesis. Annual Review of Ecology, Evolution, and Systematics, 34, 273-309. |
| [25] | Wu ZY (吴征镒) (1991) The areal-types of Chinese genera of seed plants. Acta Botanica Yunnanica (云南植物研究), 13(Suppl.), 1-139. (in Chinese with English abstract) |
| [26] | Wu ZY (吴征镒), Sun H (孙航), Zhou ZK (周浙昆), Li DZ (李德铢), Peng H (彭华) (2011) Floristics of Seed Plants from China (中国种子植物区系地理). Science Press, Beijing. (in Chinese) |
| [27] | Wu ZY (吴征镒), Sun H (孙航), Zhou ZK (周浙昆), Peng H (彭华), Li DZ (李德铢) (2005) Origin and differentiation of endemism in the flora of China. Acta Botanica Yunnanica (云南植物研究), 27, 577-604. (in Chinese with English abstract) |
| [28] | Xu J (许瑾), Liu ED (刘恩德), Peng H (彭华) (2008) Distribution change of Craspedolobium schochii and unacceptability of its endemism to China. Journal of Tropical and Subtropical Botany (热带亚热带植物学报), 16, 266-270. (in Chinese with English abstract) |
| [29] | Ying TS (应俊生) (1996) Areography of the endemic genera of seed plants in China. Acta Phytotaxonomica Sinica (植物分类学报), 34, 479-485. (in Chinese with English Abstract) |
| [30] | Ying TS (应俊生), Zhang YL (张玉龙) (1994) The Endemic Genera of Seed Plants of China (中国种子植物特有属). Science Press, Beijing. (in Chinese) |
| [31] | Zhu H (朱华) (2008) Distribution patterns of genera of Yunnan seed plants with references to their biogeographical significances. Advances in Earth Science (地球科学进展), 23, 830-839. (in Chinese with English abstract) |
| [32] | Zhu H, Ma YX, Yan LC, Hu HB (2007) The relationship between geography and climate in the generic-level patterns of Chinese seed plants. Acta Phytotaxonomica Sinica (植物分类学报), 45, 134-166. |
| [1] | 杨舒涵, 王贺, 陈磊, 廖蓥飞, 严光, 伍一宁, 邹红菲. 松嫩平原异质生境对土壤线虫群落特征的影响[J]. 生物多样性, 2024, 32(1): 23295-. |
| [2] | 王明慧, 陈昭铨, 李帅锋, 黄小波, 郎学东, 胡子涵, 尚瑞广, 刘万德. 云南普洱季风常绿阔叶林不同种子扩散方式的优势种空间点格局分析[J]. 生物多样性, 2023, 31(9): 23147-. |
| [3] | 李世东. 中国和美国国家公园时空发展及驱动因素[J]. 生物多样性, 2023, 31(6): 23040-. |
| [4] | 谢艳秋, 黄晖, 王春晓, 何雅琴, 江怡萱, 刘子琳, 邓传远, 郑郁善. 福建海岛滨海特有植物种-面积关系及物种丰富度决定因素[J]. 生物多样性, 2023, 31(5): 22345-. |
| [5] | 陈宏, 冼晓青, 陈宜雪, 林娜, 王苗苗, 李志鹏, 赵健. 海岛型城市红火蚁发生程度空间格局及驱动因子——以福建海坛岛为例[J]. 生物多样性, 2023, 31(5): 22501-. |
| [6] | 丁炳扬, 金孝锋, 张永华, 李根有, 陈征海, 张方钢. 浙江野生种子植物的分布格局与区系分区[J]. 生物多样性, 2023, 31(4): 22515-. |
| [7] | 张伟, 翟东东, 熊飞, 刘红艳, 陈元元, 王莹, 廖传松, 段辛斌, 田辉伍, 邓华堂, 陈大庆. 三峡库区鱼类群落结构和功能多样性[J]. 生物多样性, 2023, 31(2): 22136-. |
| [8] | 田希, 刘文聪, 饶杰生, 王晓凤, 杨涛, 陈稀, 张秋雨, 刘其明, 徐衍潇, 张旭, 沈泽昊. 云南鸡足山半湿润常绿阔叶林的林隙干扰格局与成因[J]. 生物多样性, 2023, 31(11): 23219-. |
| [9] | 孙维悦, 舒江平, 顾钰峰, 莫日根高娃, 杜夏瑾, 刘保东, 严岳鸿. 基于保护基因组学揭示荷叶铁线蕨的濒危机制[J]. 生物多样性, 2022, 30(7): 21508-. |
| [10] | 金孝锋, 鲁益飞, 丁炳扬, 李根有, 陈征海, 张方钢. 浙江种子植物物种编目[J]. 生物多样性, 2022, 30(6): 21408-. |
| [11] | 付飞, 魏慧玉, 常育腾, 王备新, 陈凯. 澜沧江中游水生昆虫生活史和生态学性状多样性的海拔格局: 气候和土地利用的影响[J]. 生物多样性, 2022, 30(5): 21332-. |
| [12] | 孙佳欢, 刘冬, 朱家祺, 张书宁, 高梅香. 小麦-玉米轮作农田土壤螨多样性空间分布格局[J]. 生物多样性, 2022, 30(12): 22292-. |
| [13] | 许祖昌, 罗亚皇, 秦声远, 朱光福, 李德铢. 中国竹类植物馆藏标本现状与地理分布[J]. 生物多样性, 2021, 29(7): 897-909. |
| [14] | 赵万义, 刘忠成, 叶华谷, 王蕾, 陈功锡, 刘克明, 詹选怀, 廖文波. 罗霄山脉种子植物区系及其南北分化特征[J]. 生物多样性, 2020, 28(7): 842-853. |
| [15] | 宋垚彬, 徐力, 段俊鹏, 张卫军, 申屠晓露, 李天翔, 臧润国, 董鸣. 西藏极小种群野生植物密叶红豆杉种群的性比及雌雄空间格局[J]. 生物多样性, 2020, 28(3): 269-276. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
备案号:京ICP备16067583号-7
Copyright © 2026 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn
![]()