
生物多样性 ›› 2021, Vol. 29 ›› Issue (12): 1629-1637. DOI: 10.17520/biods.2021341 cstr: 32101.14.biods.2021341
叶俊伟1,2, 田斌1,*( )
)
收稿日期:2021-08-30
接受日期:2021-11-19
出版日期:2021-12-20
发布日期:2021-12-16
通讯作者:
田斌
作者简介:*E-mail: tianbin@swfu.edu.cn基金资助:
Junwei Ye1,2, Bin Tian1,*()
Received:2021-08-30
Accepted:2021-11-19
Online:2021-12-20
Published:2021-12-16
Contact:
Bin Tian
摘要:
扁核木(Prinsepia utilis)为中国西南地区温带森林重要的木本油料植物, 但对其野生资源种群遗传结构及成因的了解严重不足。我们采用核微卫星分子标记, 对32个扁核木自然种群共377个个体的群体演化历史进行了探讨, 并评估其遗传资源。研究发现扁核木种群自西向东可划分4个遗传群组, 即喜马拉雅、横断山以及云贵高原西部和东部群组。其中, 最大的遗传分化存在于喜马拉雅和其他区域种群间。与喜马拉雅和云贵高原东部群组相比, 横断山和云贵高原西部群组混合了其他群组的遗传成分。种群动态历史分析显示中部2个群组在喜马拉雅和云贵高原东部群组形成后形成, 不同群组间的分化均发生在更新世晚期。地理隔离和环境隔离分析表明扁核木种群间的遗传分化主要由环境差异导致。环境差异分析显示不同群组间的气候存在不同程度的差异, 其中喜马拉雅和云贵高原东部群组与中部2个群组间的差异显著。此外, 结合该物种不同时期的生态位模拟数据, 我们认为喜马拉雅和云贵高原地区的遗传资源在未来需要优先保护。
叶俊伟, 田斌 (2021) 中国西南地区重要木本油料植物扁核木的遗传结构及成因. 生物多样性, 29, 1629-1637. DOI: 10.17520/biods.2021341.
Junwei Ye, Bin Tian (2021) Genetic structure and its causes of an important woody oil plant in Southwest China, Prinsepia utilis (Rosaceae). Biodiversity Science, 29, 1629-1637. DOI: 10.17520/biods.2021341.
| 代号 Code | 地点 Site | 经纬度 Location | 海拔 Altitude (m) | n | AO | HE | HO | RS | PAR | FIS |
|---|---|---|---|---|---|---|---|---|---|---|
| BAZY | 云南昆明 Kunming, Yunnan | 102.72˚ E, 25.40˚ N | 2,370 | 9 | 20 | 0.45 | 0.63 | 2.45 | 0.01 | -0.35 |
| BBM | 西藏波密 Bomi, Xizang | 95.34˚ E, 29.99˚ N | 2,750 | 14 | 22 | 0.54 | 0.76 | 2.61 | 0 | -0.37 |
| BDC | 四川稻城 Daocheng, Sichuan | 100.30˚ E, 28.58˚ N | 3,310 | 12 | 21 | 0.46 | 0.62 | 2.55 | 0.12 | -0.31 |
| BDCQ | 云南会泽 Huize, Yunnan | 103.10˚ E, 25.94˚ N | 2,220 | 10 | 22 | 0.48 | 0.67 | 2.74 | 0.07 | -0.35 |
| BES | 云南峨山 E’shan, Yunnan | 102.11˚ E, 24.28˚ N | 2,060 | 6 | 15 | 0.38 | 0.62 | - | - | -0.58 |
| BHD | 四川会东 Huidong, Sichuan | 102.78˚ E, 26.55˚ N | 2,370 | 14 | 22 | 0.47 | 0.65 | 2.61 | 0 | -0.36 |
| BHQ | 云南鹤庆 Heqing, Yunnan | 100.18˚ E, 26.56˚ N | 2,200 | 15 | 27 | 0.64 | 0.86 | 3.28 | 0 | -0.31 |
| BJL | 西藏吉隆 Jilong, Xizang | 85.36˚ E, 28.39˚ N | 3,140 | 12 | 19 | 0.46 | 0.65 | 2.40 | 0.03 | -0.38 |
| BJP | 云南澄江 Chengjiang, Yunnan | 102.86˚ E, 24.73˚ N | 2,180 | 15 | 25 | 0.58 | 0.81 | 2.93 | 0 | -0.38 |
| BLD | 云南玉龙 Yulong, Yunnan | 99.46˚ E, 27.18˚ N | 2,630 | 16 | 21 | 0.56 | 0.88 | 2.59 | 0 | -0.54 |
| BLJ | 云南丽江 Lijiang, Yunnan | 100.23˚ E, 27.18˚ N | 3,650 | 5 | 19 | 0.52 | 0.78 | - | - | -0.47 |
| BLJS | 云南老君山 Laojun Mountains, Yunnan | 103.93˚ E, 23.30˚ N | 1,640 | 15 | 25 | 0.55 | 0.84 | 2.83 | 0 | -0.51 |
| BLJW | 贵州安顺 Anshun, Guizhou | 105.94˚ E, 26.25˚ N | 1,400 | 15 | 23 | 0.49 | 0.69 | 2.63 | 0.10 | -0.36 |
| BLLX | 贵州龙里 Longli, Guizhou | 106.90˚ E, 26.43˚ N | 1,150 | 10 | 18 | 0.45 | 0.61 | 2.30 | 0 | -0.37 |
| BLP | 云南兰坪 Lanping, Yunnan | 99.42˚ E, 26.46˚ N | 2,540 | 12 | 24 | 0.61 | 0.98 | 2.93 | 0 | -0.57 |
| BMG | 云南马关 Maguan, Yunnan | 104.39˚ E, 23.02˚ N | 1,340 | 12 | 24 | 0.53 | 0.79 | 2.80 | 0.02 | -0.45 |
| BML | 四川木里 Muli, Sichuan | 101.25˚ E, 27.97˚ N | 2,500 | 12 | 26 | 0.61 | 0.96 | 3.05 | 0.19 | -0.55 |
| BNH | 云南南华 Nanhua, Yunnan | 101.01˚ E, 25.33˚ N | 2,320 | 9 | 21 | 0.52 | 0.75 | 2.73 | 0 | -0.39 |
| BQJ | 云南曲靖 Qujing, Yunnan | 103.91˚ E, 25.36˚ N | 1,900 | 8 | 20 | 0.49 | 0.74 | - | - | -0.44 |
| BQL | 贵州贵阳 Guiyang, Guizhou | 106.69˚ E, 26.60˚ N | 1,160 | 6 | 17 | 0.45 | 0.60 | - | - | -0.24 |
| BSZ | 云南师宗 Shizong, Yunnan | 103.98˚ E, 24.82˚ N | 1,870 | 12 | 25 | 0.52 | 0.71 | 2.91 | 0.03 | -0.34 |
| BTM | 西藏林芝 Linzhi, Xizang | 96.58˚ E, 30.50˚ N | 4,720 | 15 | 25 | 0.59 | 0.85 | 2.83 | 0.09 | -0.41 |
| BWM | 云南禄劝 Luquan, Yunnan | 102.83˚ E, 26.02˚ N | 2,760 | 14 | 22 | 0.47 | 0.69 | 2.49 | 0 | -0.46 |
| BWN | 贵州威宁 Weining, Guizhou | 104.11˚ E, 26.74˚ N | 2,450 | 13 | 24 | 0.48 | 0.68 | 2.61 | 0 | -0.38 |
| BWX | 云南维西 Weixi, Yunnan | 99.02˚ E, 27.80˚ N | 2,290 | 9 | 24 | 0.59 | 0.87 | 2.97 | 0.05 | -0.43 |
| BXC | 四川西昌 Xichang, Sichuan | 102.35˚ E, 27.70˚ N | 2,680 | 11 | 20 | 0.39 | 0.57 | 2.38 | 0.03 | -0.41 |
| BXCX | 云南西畴 Xichou, Yunnan | 104.79˚ E, 23.38˚ N | 1,450 | 9 | 25 | 0.53 | 0.75 | 2.86 | 0 | -0.37 |
| BXJ | 云南文山新街 Xinjie, Wenshan, Yunnan | 104.00˚ E, 23.20˚ N | 1,880 | 15 | 23 | 0.53 | 0.81 | 2.76 | 0 | -0.49 |
| BXZC | 云南开远 Kaiyuan, Yunnan | 103.62˚ E, 23.79˚ N | 1,480 | 10 | 21 | 0.54 | 0.80 | 2.77 | 0 | -0.43 |
| BYG | 西藏波密 Bomi, Xizang | 95.57˚ E, 30.50˚ N | 5,230 | 15 | 22 | 0.58 | 0.88 | 2.79 | 0.01 | -0.48 |
| BYXB | 四川越西 Yuexi, Sichuan | 102.45˚ E, 28.76˚ N | 2,970 | 12 | 16 | 0.39 | 0.60 | 2.12 | 0 | -0.49 |
| BZNX | 云南文山 Wenshan, Yunnan | 104.42˚ E, 23.69˚ N | 1,570 | 15 | 25 | 0.55 | 0.81 | 2.97 | 0 | -0.45 |
表1 扁核木种群取样和遗传多样性信息
Table 1 Sampling and genetic diversity information of Prinsepia utilis
| 代号 Code | 地点 Site | 经纬度 Location | 海拔 Altitude (m) | n | AO | HE | HO | RS | PAR | FIS |
|---|---|---|---|---|---|---|---|---|---|---|
| BAZY | 云南昆明 Kunming, Yunnan | 102.72˚ E, 25.40˚ N | 2,370 | 9 | 20 | 0.45 | 0.63 | 2.45 | 0.01 | -0.35 |
| BBM | 西藏波密 Bomi, Xizang | 95.34˚ E, 29.99˚ N | 2,750 | 14 | 22 | 0.54 | 0.76 | 2.61 | 0 | -0.37 |
| BDC | 四川稻城 Daocheng, Sichuan | 100.30˚ E, 28.58˚ N | 3,310 | 12 | 21 | 0.46 | 0.62 | 2.55 | 0.12 | -0.31 |
| BDCQ | 云南会泽 Huize, Yunnan | 103.10˚ E, 25.94˚ N | 2,220 | 10 | 22 | 0.48 | 0.67 | 2.74 | 0.07 | -0.35 |
| BES | 云南峨山 E’shan, Yunnan | 102.11˚ E, 24.28˚ N | 2,060 | 6 | 15 | 0.38 | 0.62 | - | - | -0.58 |
| BHD | 四川会东 Huidong, Sichuan | 102.78˚ E, 26.55˚ N | 2,370 | 14 | 22 | 0.47 | 0.65 | 2.61 | 0 | -0.36 |
| BHQ | 云南鹤庆 Heqing, Yunnan | 100.18˚ E, 26.56˚ N | 2,200 | 15 | 27 | 0.64 | 0.86 | 3.28 | 0 | -0.31 |
| BJL | 西藏吉隆 Jilong, Xizang | 85.36˚ E, 28.39˚ N | 3,140 | 12 | 19 | 0.46 | 0.65 | 2.40 | 0.03 | -0.38 |
| BJP | 云南澄江 Chengjiang, Yunnan | 102.86˚ E, 24.73˚ N | 2,180 | 15 | 25 | 0.58 | 0.81 | 2.93 | 0 | -0.38 |
| BLD | 云南玉龙 Yulong, Yunnan | 99.46˚ E, 27.18˚ N | 2,630 | 16 | 21 | 0.56 | 0.88 | 2.59 | 0 | -0.54 |
| BLJ | 云南丽江 Lijiang, Yunnan | 100.23˚ E, 27.18˚ N | 3,650 | 5 | 19 | 0.52 | 0.78 | - | - | -0.47 |
| BLJS | 云南老君山 Laojun Mountains, Yunnan | 103.93˚ E, 23.30˚ N | 1,640 | 15 | 25 | 0.55 | 0.84 | 2.83 | 0 | -0.51 |
| BLJW | 贵州安顺 Anshun, Guizhou | 105.94˚ E, 26.25˚ N | 1,400 | 15 | 23 | 0.49 | 0.69 | 2.63 | 0.10 | -0.36 |
| BLLX | 贵州龙里 Longli, Guizhou | 106.90˚ E, 26.43˚ N | 1,150 | 10 | 18 | 0.45 | 0.61 | 2.30 | 0 | -0.37 |
| BLP | 云南兰坪 Lanping, Yunnan | 99.42˚ E, 26.46˚ N | 2,540 | 12 | 24 | 0.61 | 0.98 | 2.93 | 0 | -0.57 |
| BMG | 云南马关 Maguan, Yunnan | 104.39˚ E, 23.02˚ N | 1,340 | 12 | 24 | 0.53 | 0.79 | 2.80 | 0.02 | -0.45 |
| BML | 四川木里 Muli, Sichuan | 101.25˚ E, 27.97˚ N | 2,500 | 12 | 26 | 0.61 | 0.96 | 3.05 | 0.19 | -0.55 |
| BNH | 云南南华 Nanhua, Yunnan | 101.01˚ E, 25.33˚ N | 2,320 | 9 | 21 | 0.52 | 0.75 | 2.73 | 0 | -0.39 |
| BQJ | 云南曲靖 Qujing, Yunnan | 103.91˚ E, 25.36˚ N | 1,900 | 8 | 20 | 0.49 | 0.74 | - | - | -0.44 |
| BQL | 贵州贵阳 Guiyang, Guizhou | 106.69˚ E, 26.60˚ N | 1,160 | 6 | 17 | 0.45 | 0.60 | - | - | -0.24 |
| BSZ | 云南师宗 Shizong, Yunnan | 103.98˚ E, 24.82˚ N | 1,870 | 12 | 25 | 0.52 | 0.71 | 2.91 | 0.03 | -0.34 |
| BTM | 西藏林芝 Linzhi, Xizang | 96.58˚ E, 30.50˚ N | 4,720 | 15 | 25 | 0.59 | 0.85 | 2.83 | 0.09 | -0.41 |
| BWM | 云南禄劝 Luquan, Yunnan | 102.83˚ E, 26.02˚ N | 2,760 | 14 | 22 | 0.47 | 0.69 | 2.49 | 0 | -0.46 |
| BWN | 贵州威宁 Weining, Guizhou | 104.11˚ E, 26.74˚ N | 2,450 | 13 | 24 | 0.48 | 0.68 | 2.61 | 0 | -0.38 |
| BWX | 云南维西 Weixi, Yunnan | 99.02˚ E, 27.80˚ N | 2,290 | 9 | 24 | 0.59 | 0.87 | 2.97 | 0.05 | -0.43 |
| BXC | 四川西昌 Xichang, Sichuan | 102.35˚ E, 27.70˚ N | 2,680 | 11 | 20 | 0.39 | 0.57 | 2.38 | 0.03 | -0.41 |
| BXCX | 云南西畴 Xichou, Yunnan | 104.79˚ E, 23.38˚ N | 1,450 | 9 | 25 | 0.53 | 0.75 | 2.86 | 0 | -0.37 |
| BXJ | 云南文山新街 Xinjie, Wenshan, Yunnan | 104.00˚ E, 23.20˚ N | 1,880 | 15 | 23 | 0.53 | 0.81 | 2.76 | 0 | -0.49 |
| BXZC | 云南开远 Kaiyuan, Yunnan | 103.62˚ E, 23.79˚ N | 1,480 | 10 | 21 | 0.54 | 0.80 | 2.77 | 0 | -0.43 |
| BYG | 西藏波密 Bomi, Xizang | 95.57˚ E, 30.50˚ N | 5,230 | 15 | 22 | 0.58 | 0.88 | 2.79 | 0.01 | -0.48 |
| BYXB | 四川越西 Yuexi, Sichuan | 102.45˚ E, 28.76˚ N | 2,970 | 12 | 16 | 0.39 | 0.60 | 2.12 | 0 | -0.49 |
| BZNX | 云南文山 Wenshan, Yunnan | 104.42˚ E, 23.69˚ N | 1,570 | 15 | 25 | 0.55 | 0.81 | 2.97 | 0 | -0.45 |
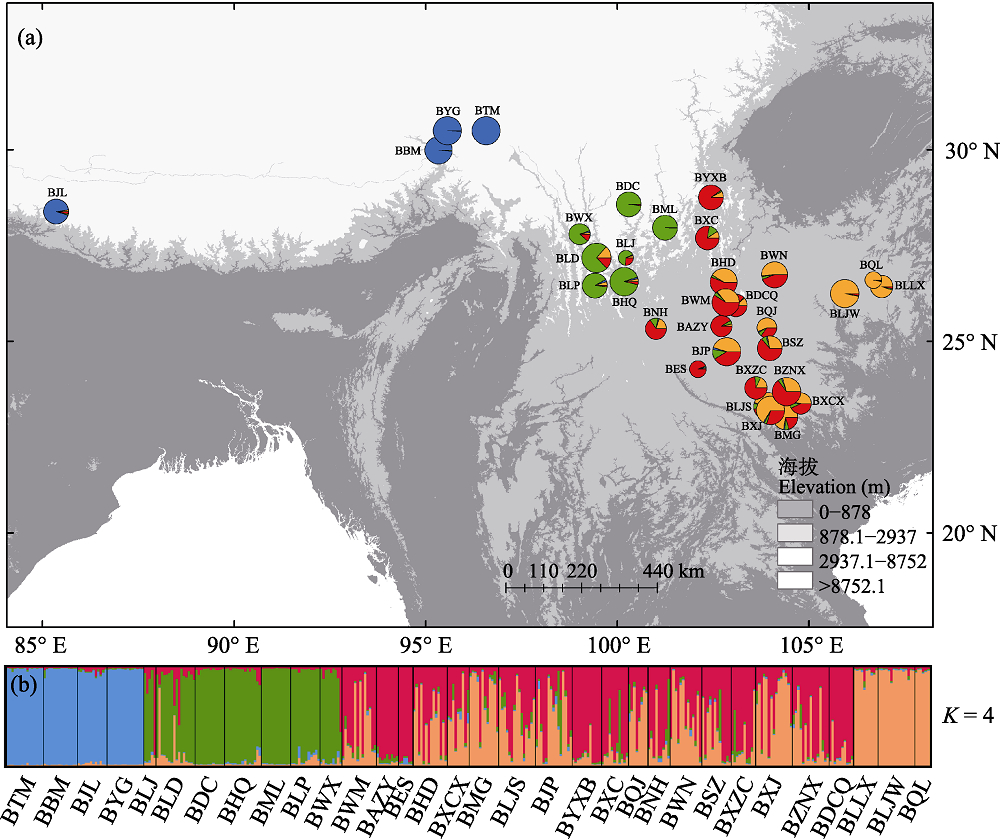
图1 扁核木32个种群遗传群组(K = 4)的分布(a)和不同个体所属群组比值的柱状图(b)。蓝色为喜马拉雅群组, 绿色为横断山群组, 红色和橙色分别是云贵高原西部和东部群组。种群代号见表1。
Fig. 1 Color-coded clustering (a) and histogram of the cluster assignment of different individuals (b) of the 32 Prinsepia utilis populations when K = 4. Blue represents Himalayas group, green represents Hengduan Mountains group, red and orange represent west and east Yunnan-Guizhou Plateau group, respectively. Population codes are the same as that in Table 1.
| 遗传群组 Genetic cluster | 等位基因数量 AO | 观测杂合度 HO | 期望杂合度 HE | 等位基因丰富度 RS | 私有等位基因丰富度 PAR |
|---|---|---|---|---|---|
| 喜马拉雅 Himalayas | 30 | 0.79 | 0.62 | 3.18 | 0.70 |
| 横断山 Hengduan Mountains | 35 | 0.86 | 0.65 | 3.39 | 0.53 |
| 云贵高原西部 West Yunnan-Guizhou Plateau | 36 | 0.73 | 0.56 | 3.04 | 0.19 |
| 云贵高原东部 East Yunnan-Guizhou Plateau | 26 | 0.65 | 0.50 | 2.69 | 0.24 |
表2 扁核木不同遗传群组的遗传多样性
Table 2 Genetic diversity of different genetic cluster of Prinsepia utilis
| 遗传群组 Genetic cluster | 等位基因数量 AO | 观测杂合度 HO | 期望杂合度 HE | 等位基因丰富度 RS | 私有等位基因丰富度 PAR |
|---|---|---|---|---|---|
| 喜马拉雅 Himalayas | 30 | 0.79 | 0.62 | 3.18 | 0.70 |
| 横断山 Hengduan Mountains | 35 | 0.86 | 0.65 | 3.39 | 0.53 |
| 云贵高原西部 West Yunnan-Guizhou Plateau | 36 | 0.73 | 0.56 | 3.04 | 0.19 |
| 云贵高原东部 East Yunnan-Guizhou Plateau | 26 | 0.65 | 0.50 | 2.69 | 0.24 |
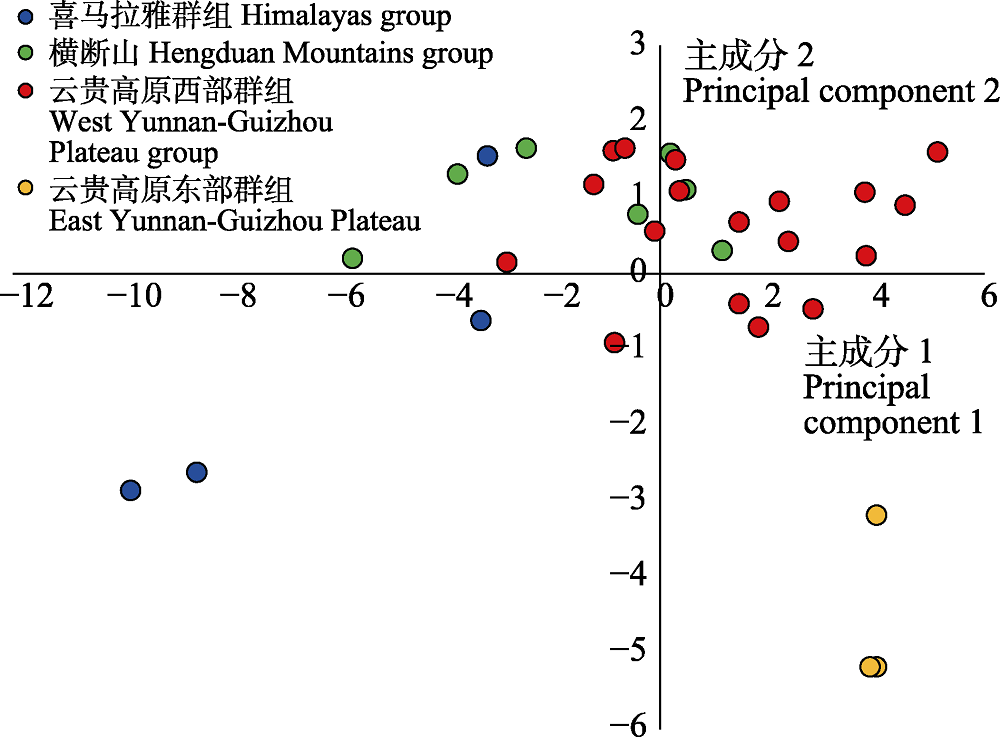
图2 扁核木32个种群间气候变量的主成分分析, 其中不同遗传群组的颜色与图1中相对应。
Fig. 2 Principal component analysis of climate variables of 32 Prinsepia utilis populations. Coded colors of different genetic clusters correspond to those in Fig. 1.
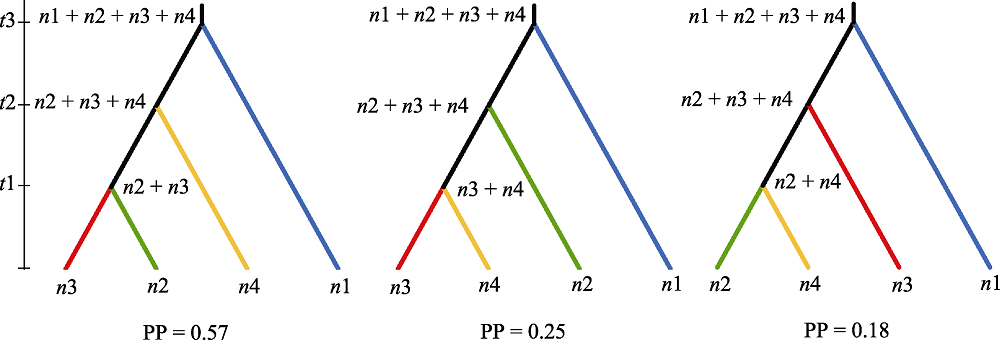
图3 DIYABC 分析中3种不同的扁核木种群分化历史示意图。n1-4分别为喜马拉雅、横断山以及云贵高原西部和东部遗传群组的有效种群大小, t1-3分别为不同分化事件的分化时间(以世代为单位), PP为后验概率。
Fig. 3 Illustration of three different scenarios of divergence history of Prinsepia utilis populations. n1-4 represent effective population sizes of Himalayas, Hengduan Mountains, west and east Yunnan-Guizhou Plateau genetic groups, respectively; t1-3 represent divergence time of different divergence events (in generations); and PP represents posterior probability.
| n1 | n2 | n3 | n4 | T1 | T2 | T3 | μ | P | |
|---|---|---|---|---|---|---|---|---|---|
| 中位值 Median | 6.84×104 | 6.30×104 | 5.84×104 | 3.90×104 | 8.53×104 | 1.25×105 | 5.11×105 | 4.30×10-6 | 0.21 |
| 95%置信区间下限 95% HPD lower | 3.30×104 | 2.68×104 | 2.39×104 | 1.21×104 | 1.80×104 | 2.73×104 | 1.92×105 | 2.19×10-6 | 0.11 |
| 95%置信区间上限 95% HPD upper | 9.53×104 | 9.44×104 | 9.15×104 | 8.50×104 | 3.49×105 | 4.49×105 | 8.82×105 | 1.01×10-5 | 0.70 |
表3 DIYABC模拟扁核木种群最优分化历史中不同参数的后验中位值及95%置信区间
Table 3 Posterior median and 95% highest posterior density interval (HPD) estimate for demographic parameters in the most possible divergence scenario of Prinsepia utilis in DIYABC
| n1 | n2 | n3 | n4 | T1 | T2 | T3 | μ | P | |
|---|---|---|---|---|---|---|---|---|---|
| 中位值 Median | 6.84×104 | 6.30×104 | 5.84×104 | 3.90×104 | 8.53×104 | 1.25×105 | 5.11×105 | 4.30×10-6 | 0.21 |
| 95%置信区间下限 95% HPD lower | 3.30×104 | 2.68×104 | 2.39×104 | 1.21×104 | 1.80×104 | 2.73×104 | 1.92×105 | 2.19×10-6 | 0.11 |
| 95%置信区间上限 95% HPD upper | 9.53×104 | 9.44×104 | 9.15×104 | 8.50×104 | 3.49×105 | 4.49×105 | 8.82×105 | 1.01×10-5 | 0.70 |
| [1] | Bai WN, Zhang DY (2014) Current status and future directions in plant phylogeography. Chinese Bulletin of Life Sciences, 26, 125-137. (in Chinese with English abstract) |
| [ 白伟宁, 张大勇 (2014) 植物亲缘地理学的研究现状与发展趋势. 生命科学, 26, 125-137.] | |
| [2] |
Cornuet JM, Pudlo P, Veyssier J, Dehne-Garcia A, Gautier M, Leblois R, Marin JM, Estoup A (2014) DIYABC v2.0: A software to make approximate Bayesian computation inferences about population history using single nucleotide polymorphism, DNA sequence and microsatellite data. Bioinformatics, 30, 1187-1189.
DOI URL |
| [3] |
Ding WN, Ree RH, Spicer RA, Xing YW (2020) Ancient orogenic and monsoon-driven assembly of the world’s richest temperate alpine flora. Science, 369, 578-581.
DOI URL |
| [4] |
Doyle JJ, Doyle JL (1990) Isolation of Plant DNA from fresh tissue. Focus, 12, 13-15.
DOI URL |
| [5] |
Evanno G, Regnaut S, Goudet J (2005) Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Molecular Ecology, 14, 2611-2620.
PMID |
| [6] |
Falush D, Stephens M, Pritchard JK (2007) Inference of population structure using multilocus genotype data: Dominant markers and null alleles. Molecular Ecology Notes, 7, 574-578.
DOI URL |
| [7] | Goudet J (2001) FSTAT, A Program to Estimate and Test Gene Diversities and Fixation Indices (version 2.9.3). https://www2.unil.ch/popgen/softwares/fstat.htm . (accessed on 2021-08-30) |
| [8] |
Guan B, Peng CC, Wang CH, Jin HZ, Zhang WD (2014) Chemical constituents from the aerial parts of Prinsepia utilis. Chemistry of Natural Compounds, 50, 1106-1107.
DOI URL |
| [9] |
Guan B, Peng CC, Zeng Q, Cheng XR, Yan SK, Jin HZ, Zhang WD (2013) Cytotoxic pentacyclic triterpenoids from Prinsepia utilis. Planta Medica, 79, 365-368.
DOI PMID |
| [10] | He QJ, He JW, Wang YP, Mu YQ, Yang WH, Yang HT, Huang XE, Wu YB, He SL, He JP (2016) Overview of Prinsepia utilis Royle. Chinese Agricultural Science Bulletin, 32(7), 74-78. (in Chinese with English abstract) |
| [ 和琼姬, 和加卫, 王宇萍, 木永青, 杨文宏, 杨洪涛, 黄杏娥, 吴永斌, 和寿莲, 和建平 (2016) 青刺果研究概述. 中国农学通报, 32(7), 74-78.] | |
| [11] |
Hijmans RJ, Cameron SE, Parra JL, Jones PG, Jarvis A (2005) Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology, 25, 1965- 1978.
DOI URL |
| [12] |
Huang SQ, Ma YL, Zhang CT, Cai SB, Pang MJ (2017) Bioaccessibility and antioxidant activity of phenolics in native and fermented Prinsepia utilis Royle seed during a simulated gastrointestinal digestion in vitro. Journal of Functional Foods, 37, 354-362.
DOI URL |
| [13] |
Jin WY, Li HW, Wei R, Huang BH, Liu B, Sun TT, Mabberley DJ, Liao PC, Yang Y (2020) New insights into biogeographical disjunctions between Taiwan and the Eastern Himalayas: The case of Prinsepia (Rosaceae). Taxon, 69, 278-289.
DOI URL |
| [14] |
Ju MM, Ma HC, Xin PY, Zhou ZL, Tian B (2015) Development and characterization of EST-SSR markers in Bombax ceiba (Malvaceae). Applications in Plant Sciences, 3, 1500001.
DOI URL |
| [15] |
Kalinowski ST (2005) Hp-rare 1.0: A computer program for performing rarefaction on measures of allelic richness. Molecular Ecology Notes, 5, 187-189.
DOI URL |
| [16] |
Li Q, Zhao Y, Xiang XG, Chen JK, Rong J (2019) Genetic diversity of crop wild relatives under threat in Yangtze River basin: Call for enhanced in situ conservation and utilization. Molecular Plant, 12, 1535-1538.
DOI URL |
| [17] | Li XW, Li J (1993) A preliminary floristic study on the seed plants from the region of Hengduan Mountain. Acta Botanica Yunnanica, 15, 217-231. (in Chinese with English abstract) |
| [ 李锡文, 李捷 (1993) 横断山脉地区种子植物区系的初步研究. 云南植物研究, 15, 217-231.] | |
| [18] |
Liu J, Luo YH, Li DZ, Gao LM (2017) Evolution and maintenance mechanisms of plant diversity in the Qinghai- Tibet Plateau and adjacent regions: Restrospect and prospect. Biodiversity Science, 25, 163-174. (in Chinese with English abstract)
DOI |
|
[ 刘杰, 罗亚皇, 李德铢, 高连明 (2017) 青藏高原及毗邻区植物多样性演化与维持机制: 进展及展望. 生物多样性, 25, 163-174.]
DOI |
|
| [19] |
Liu J, Möller M, Provan J, Gao LM, Poudel RC, Li DZ (2013) Geological and ecological factors drive cryptic speciation of yews in a biodiversity hotspot. New Phytologist, 199, 1093-1108.
DOI URL |
| [20] |
Lu LM, Mao LF, Yang T, Ye JF, Liu B, Li HL, Sun M, Miller JT, Mathews S, Hu HH, Niu YT, Peng DX, Chen YH, Smith SA, Chen M, Xiang KL, Le CT, Dang VC, Lu AM, Soltis PS, Soltis DE, Li JH, Chen ZD (2018) Evolutionary history of the angiosperm flora of China. Nature, 554, 234-238.
DOI URL |
| [21] |
Luo D, Yue JP, Sun WG, Xu B, Li ZM, Comes HP, Sun H (2016) Evolutionary history of the subnival flora of the Himalaya‐Hengduan Mountains: First insights from comparative phylogeography of four perennial herbs. Journal of Biogeography, 43, 31-43.
DOI URL |
| [22] |
Ma XG, Wang ZW, Tian B, Sun H (2019) Phylogeographic analyses of the East Asian endemic genus Prinsepia and the role of the East Asian monsoon system in shaping a north-south divergence pattern in China. Frontiers in Genetics, 10, 128.
DOI URL |
| [23] |
Ma YZ, Wang J, Hu QJ, Li JL, Sun YS, Zhang L, Abbott RJ, Liu JQ, Mao KS (2019) Ancient introgression drives adaptation to cooler and drier mountain habitats in a cypress species complex. Communications Biology, 2, 213.
DOI URL |
| [24] |
Muellner-Riehl AN (2019) Mountains as evolutionary arenas: Patterns, emerging approaches, paradigm shifts, and their implications for plant phylogeographic research in the Tibeto-Himalayan region. Frontiers in Plant Science, 10, 195.
DOI PMID |
| [25] |
Peakall R, Smouse PE (2012) GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-An update. Bioinformatics, 28, 2537-2539.
PMID |
| [26] |
Pritchard JK, Stephens M, Donnelly P (2000) Inference of population structure using multilocus genotype data. Genetics, 155, 945-959.
PMID |
| [27] |
Sun H, Zhang JW, Deng T, Boufford DE (2017) Origins and evolution of plant diversity in the Hengduan Mountains, China. Plant Diversity, 39, 161-166.
DOI URL |
| [28] |
van Oosterhout C, Hutchinson WF, Wills DPM, Shipley P (2004) Micro-checker: Software for identifying and correcting genotyping errors in microsatellite data. Molecular Ecology Notes, 4, 535-538.
DOI URL |
| [29] |
Wang S, Shi C, Gao LZ (2013) Plastid genome sequence of a wild woody oil species, Prinsepia utilis, provides insights into evolutionary and mutational patterns of Rosaceae chloroplast genomes. PLoS ONE, 8, e73946.
DOI URL |
| [30] |
Ward FK (1921) The Mekong-Salween Divide as a geographical barrier. The Geographical Journal, 58, 49-56.
DOI URL |
| [31] | Xing YW, Ree RH (2017) Uplift-driven diversification in the Hengduan Mountains, a temperate biodiversity hotspot. Proceedings of the National Academy of Sciences, USA, 114, E3444-E3451. |
| [32] |
Ye JW, Zhang Y, Wang XJ (2017) Phylogeographic breaks and the mechanisms of their formation in the Sino-Japanese floristic region. Chinese Journal of Plant Ecology, 41, 1003-1019. (in Chinese with English abstract)
DOI URL |
|
[ 叶俊伟, 张阳, 王晓娟 (2017) 中国-日本植物区系中的谱系地理间断及其形成机制. 植物生态学报, 41, 1003-1019.]
DOI |
| [1] | 贾晓旭, 陈皖强, 唐修君, 樊艳凤, 张静, 王海威, 高玉时. 西南地区家鸡线粒体DNA控制区遗传多样性和基因渗入[J]. 生物多样性, 2026, 34(5): 26003-. |
| [2] | 姜媛, 黄贝希, 贾学圆, 梁思, 谢雨彤, 范平, 宋刚. 一种基于不等长DNA序列的单倍型丰富度估算方法[J]. 生物多样性, 2026, 34(1): 25263-. |
| [3] | 王儒晓, 史博洋, 潘达, 孙红英. 中国特有华溪蟹属淡水蟹多样性格局及其保护空缺[J]. 生物多样性, 2025, 33(8): 25123-. |
| [4] | 周智成, 曹天玲, 刘如垚, 丁琪琪, 马轲, 杨丽萍, 周传江, 聂国兴, 汤永涛. 基于线粒体COI基因的黄河流域麦穗鱼种群遗传多样性与遗传结构[J]. 生物多样性, 2025, 33(8): 24501-. |
| [5] | 范平, 王欢, 温知新, 宋刚, 雷富民. 气候因子对鸟类遗传多样性与物种分布面积关系的影响[J]. 生物多样性, 2025, 33(8): 25072-. |
| [6] | 薛瑞翔, 马雪蓉, 吴炯文, 刘爱君, 张细权, 季从亮, 殷颖珊, 朱炜健, 罗庆斌. 中山麻鸭群体遗传多样性与遗传结构[J]. 生物多样性, 2025, 33(8): 24592-. |
| [7] | 范平, 温知新, 宋刚. 气候因子和人类活动对两栖及哺乳动物不同遗传多样性指标的影响[J]. 生物多样性, 2025, 33(8): 25022-. |
| [8] | 伍金山, 杨长乐, 马玉凤, 李亚旋, 高文家, 叶·库斯力, 叶樑洪, 杨宇骄, 徐梦琦, 廖廷琼, 钟林强, 单文娟. 艾比湖湿地国家级自然保护区马鹿遗传多样性及遗传结构[J]. 生物多样性, 2025, 33(12): 25233-. |
| [9] | 王嘉陈, 徐汤俊, 许唯, 张高季, 尤艺瑾, 阮宏华, 刘宏毅. 城市景观格局对大蚰蜒种群遗传结构的影响[J]. 生物多样性, 2025, 33(1): 24251-. |
| [10] | 冯晨, 张洁, 黄宏文. 统筹植物就地保护与迁地保护的解决方案: 植物并地保护(parallel situ conservation)[J]. 生物多样性, 2023, 31(9): 23184-. |
| [11] | 李庆多, 栗冬梅. 全球蝙蝠巴尔通体流行状况分析[J]. 生物多样性, 2023, 31(9): 23166-. |
| [12] | 齐海玲, 樊鹏振, 王跃华, 刘杰. 中国北方六省区胡桃的遗传多样性和群体结构[J]. 生物多样性, 2023, 31(8): 23120-. |
| [13] | 熊飞, 刘红艳, 翟东东, 段辛斌, 田辉伍, 陈大庆. 基于基因组重测序的长江上游瓦氏黄颡鱼群体遗传结构[J]. 生物多样性, 2023, 31(4): 22391-. |
| [14] | 蒲佳佳, 杨平俊, 戴洋, 陶可欣, 高磊, 杜予州, 曹俊, 俞晓平, 杨倩倩. 长江下游外来生物福寿螺的种类及其种群遗传结构[J]. 生物多样性, 2023, 31(3): 22346-. |
| [15] | 何艺玥, 刘玉莹, 张富斌, 秦强, 曾燏, 吕振宇, 杨坤. 梯级水利工程背景下的嘉陵江干流蛇鮈群体遗传多样性和遗传结构[J]. 生物多样性, 2023, 31(11): 23160-. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
备案号:京ICP备16067583号-7
Copyright © 2026 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn
![]()