
生物多样性 ›› 2017, Vol. 25 ›› Issue (8): 799-806. DOI: 10.17520/biods.2015218 cstr: 32101.14.biods.2015218
褚建民1, 李毅夫2, 张雷1, 李斌1, 高明远1, 唐晓倩1, 倪建伟1, 许新桥1,*( )
)
收稿日期:2016-11-22
接受日期:2017-02-17
出版日期:2017-08-20
发布日期:2017-08-31
通讯作者:
许新桥
作者简介:具体评估过程包括信息汇总(各个渠道的标本信息、野外调查信息及文献资料)、逐条比对IUCN红色名录等级与标准、确定等级、填写评估说明。在具体评估过程中, 针对不同类群设计信息调查表, 通过电话和邮件向多位同行征询物种的居群信息。
基金资助:
Jianmin Chu1, Yifu Li2, Lei Zhang1, Bin Li1, Mingyuan Gao1, Xiaoqian Tang1, Jianwei Ni1, Xinqiao Xu1,*()
Received:2016-11-22
Accepted:2017-02-17
Online:2017-08-20
Published:2017-08-31
Contact:
Xu Xinqiao
摘要:
长柄扁桃(Amygdalus pedunculata)在我国分布于内蒙古和陕西, 是一种濒危灌木, 其资源现状只有零星的文献记录, 这限制了对其资源数量和保护现状的评估。为此, 本文通过野外调查来确立其自然分布区范围。我们选择了8个环境因子, 运用基于规则集的遗传算法(genetic algorithm for rule-set prediction, GARP)模型和最大熵(maximum entropy, MaxEnt)模型进行模拟, 预测了长柄扁桃在中国的潜在适宜分布区; 同时基于4个模型精度评估指标(Kappa、真实技巧统计法、总精度和受试者工作特征曲线下的面积)对模型进行验证, 采用刀切法评估了预测变量的重要性。结果表明, 两种模型均能准确预测长柄扁桃的地理分布规律, 但MaxEnt模型的4个预测精度指标都大于GARP模型。根据模型结果可判断长柄扁桃的适宜分布区以内蒙古中部地区为主, 东起大兴安岭南部, 向西可至贺兰山、乌兰布和沙漠以东, 涵盖了毛乌素沙地、库布齐沙漠和浑善达克沙地, 以及乌拉山、大青山等土石山区。其低适宜分布区可辐射至辽宁、河北、山西、陕西等省部分地区, 另外在宁夏和甘肃中部地区也有零星的低适宜分布区。变量重要性分析结果表明, 与降水相关的变量是决定长柄扁桃地理分布的重要环境因素。
褚建民, 李毅夫, 张雷, 李斌, 高明远, 唐晓倩, 倪建伟, 许新桥 (2017) 濒危物种长柄扁桃的潜在分布与保护策略. 生物多样性, 25, 799-806. DOI: 10.17520/biods.2015218.
Jianmin Chu, Yifu Li, Lei Zhang, Bin Li, Mingyuan Gao, Xiaoqian Tang, Jianwei Ni, Xinqiao Xu (2017) Potential distribution range and conservation strategies for the endangered species Amygdalus pedunculata. Biodiversity Science, 25, 799-806. DOI: 10.17520/biods.2015218.
| 数据代码 Code | 环境因子 Environmental variables |
|---|---|
| BIO2 | 平均温度日较差 Mean diurnal temperature range |
| BIO3 | 等温性 Isothermality |
| BIO5 | 最热月最高温度 Max. temperature of the warmest month |
| BIO6 | 最冷月最低温度 Min. temperature of the coldest month |
| BIO7 | 平均温度年较差 Temperature annual range |
| BIO13 | 最湿月降水量 Precipitation of wettest month |
| BIO14 | 最干月降水量 Precipitation of driest month |
| BIO15 | 降水量季节性 Precipitation seasonality |
表1 长柄扁桃分布模型构建所需环境因子
Table 1 Environmental variables used in the distribution modeling of Amygdalus pedunculata
| 数据代码 Code | 环境因子 Environmental variables |
|---|---|
| BIO2 | 平均温度日较差 Mean diurnal temperature range |
| BIO3 | 等温性 Isothermality |
| BIO5 | 最热月最高温度 Max. temperature of the warmest month |
| BIO6 | 最冷月最低温度 Min. temperature of the coldest month |
| BIO7 | 平均温度年较差 Temperature annual range |
| BIO13 | 最湿月降水量 Precipitation of wettest month |
| BIO14 | 最干月降水量 Precipitation of driest month |
| BIO15 | 降水量季节性 Precipitation seasonality |
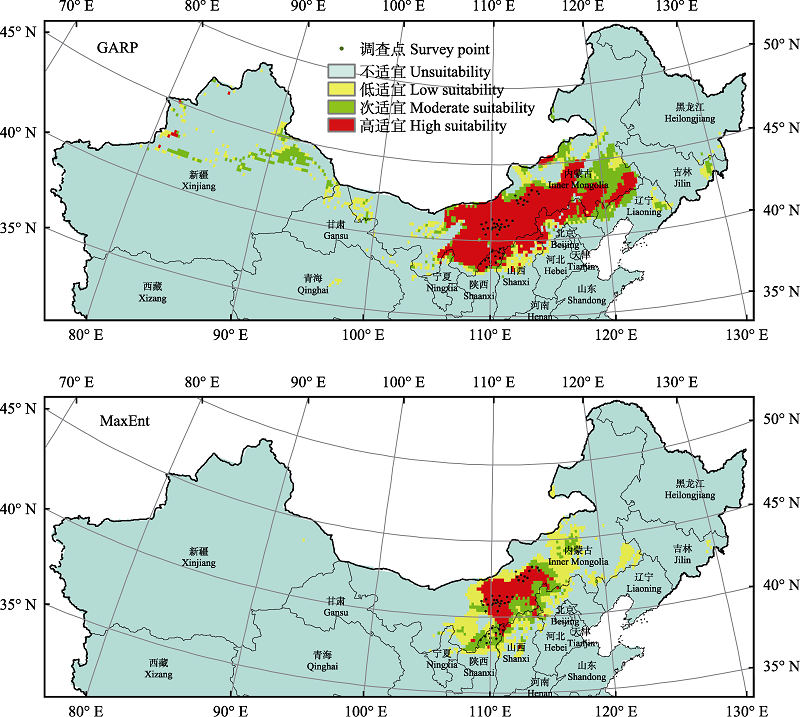
图1 基于规则集的遗传算法(GARP)模型和最大熵(MaxEnt)模型预测的长柄扁桃潜在分布区
Fig. 1 Potential distributions of Amygdalus pedunculata predicted by genetic algorithm for rule-set prediction (GARP) model and maximum entropy (MaxEnt) model
| 模型类型 Model type | Kappa | 真实技巧统计法 True skill statistic (TSS) | 总体精度 Overall accuracy | 受试者工作特征曲线下面积 Area under the receiver operating characteristic curve (AUC) |
|---|---|---|---|---|
| MaxEnt | 0.98 ± 0.01 | 0.98 ± 0.14 | 0.99 ± 0.01 | 1.00 ± 0.002 |
| GARP | 0.95 ± 0.01 | 0.95 ± 0.01 | 0.98 ± 0.01 | 0.98 ± 0.01 |
表2 基于规则集的遗传算法(GARP)模型和最大熵(MaxEnt)模型的预测精度
Table 2 Predictive accuracy of genetic algorithm for rule-set prediction (GARP) model and maximum entropy (MaxEnt) model
| 模型类型 Model type | Kappa | 真实技巧统计法 True skill statistic (TSS) | 总体精度 Overall accuracy | 受试者工作特征曲线下面积 Area under the receiver operating characteristic curve (AUC) |
|---|---|---|---|---|
| MaxEnt | 0.98 ± 0.01 | 0.98 ± 0.14 | 0.99 ± 0.01 | 1.00 ± 0.002 |
| GARP | 0.95 ± 0.01 | 0.95 ± 0.01 | 0.98 ± 0.01 | 0.98 ± 0.01 |
| 预测面积 Prediction area (×10,000 km2) | ||
|---|---|---|
| MaxEnt | GARP | |
| 高适宜区 High suitability area | 11.9373 | 48.2647 |
| 次适宜区 Moderate suitability area | 12.1625 | 24.2406 |
| 低适宜区 Low suitability area | 26.0358 | 18.4617 |
| 总计 Total | 50.1356 | 90.9670 |
表3 GARP模型和MaxEnt模型预测面积对比 Table 3 Comparison of prediction area between the GARP model and the MaxEnt model
Table 3 Comparison of prediction area between the GARP model and the MaxEnt model
| 预测面积 Prediction area (×10,000 km2) | ||
|---|---|---|
| MaxEnt | GARP | |
| 高适宜区 High suitability area | 11.9373 | 48.2647 |
| 次适宜区 Moderate suitability area | 12.1625 | 24.2406 |
| 低适宜区 Low suitability area | 26.0358 | 18.4617 |
| 总计 Total | 50.1356 | 90.9670 |
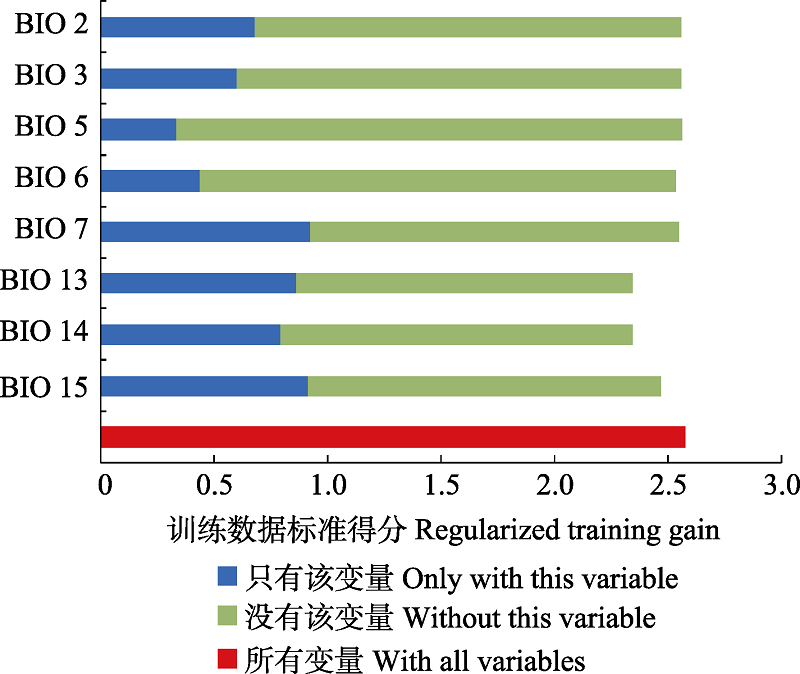
图2 MaxEnt模型中各环境因子变量的重要性。图中深蓝色越长, 表示利用该因子单独建模时其重要性越大, 而浅绿色表示除该因子外其他环境因子贡献程度的总和。变量含义见表1。
Fig. 2 Importance of environmental factors estimated by MaxEnt model. Blue bars indicate the importance of the variable of interest. Green bars indicate the importance of all variables without the variable of interest. The longer the bar, the more important the variables. The abbreviations of climatic variables are the same as in Table 1.
| [1] | Allouche O, Tsoar A, Kadmon R (2006) Assessing the accuracy of species distribution models: prevalence, Kappa and the true skill statistic (TSS). Journal of Applied Ecology, 43, 1223-1232. |
| [2] | Calenge C (2006) The package “adehabitat” for the R software: a tool for the analysis of space and habitat use by animals. Ecological Modelling, 197, 516-519. |
| [3] | Chu JM, Xu XQ, Zhang YL (2013) Production and properties of biodiesel produced from Amygdalus pedunculata Pall. Bioresource Technology, 134, 374-376. |
| [4] | Fu LG, Jin JM (1991) China Plant Red Data Book, Vol. 1: Rare and Endangered Plants. Science Press, Beijing. (in Chinese) |
| [傅立国, 金鉴明 (1991) 中国植物红皮书: 稀有濒危植物(第一册). 科学出版社, 北京.] | |
| [5] | Graham B (2001) Neutral macroecology. Science, 293, 2413-2418. |
| [6] | Guisan A, Thuiller W (2005) Predicting species distribution: offering more than simple habitat models. Ecology Letters, 8, 993-1009. |
| [7] | Guo CH, Luo M, Ma YH, Ma XW (2005) Advances of characteristic research of threatened long carpopodium almond. Journal of Northwest A & F University (Natural Science Edition), 33(12), 125-129. (in Chinese with English abstract) |
| [郭春会, 罗梦, 马玉华, 马小卫 (2005) 沙地濒危植物长柄扁桃特性研究进展. 西北农林科技大学学报(自然科学版), 33(12), 125-129.] | |
| [8] | Hampe A (2004) Bioclimate envelope models: what they detect and what they hide. Global Ecology and Biogeography, 13, 469-471. |
| [9] | Hao YQ, Zhao XF (2011) Distinguishing early-acting inbreeding depression from late-acting ovarian self-incompatibility. Biodiversity Science, 19,106-112. (in Chinese with English abstract) |
| [郝祎祺, 赵鑫峰 (2011) 被子植物早期近交衰退与晚期自交不亲和. 生物多样性, 19, 106-112.] | |
| [10] | Hijmans RJ, Cameron SE, Parra JL, Jones PG, Jarvis A (2005) Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology, 25, 1965-1978. |
| [11] | Ji ZL, Qian AD (1981) Investigation of the resources of Amygdalus pedunculata Pall. and Prunus mongolica Maxim. from the natural distribution in China. China Fruits, (2), 38-40. (in Chinese) |
| [姬钟亮, 钱安东 (1981) 长柄扁桃和蒙古扁桃在我国自然分布区的调查. 中国果树, (2), 38-40.] | |
| [12] | Lei GH, Liu LT, Han C, Shen YH, Huo YM (2009) Analysis of tocopherol content in endangered long carpopodium almond nutlet. Journal of Northwest University (Natural Science Edition), 39, 777-779. (in Chinese with English abstract) |
| [雷根虎, 刘丽婷, 韩超, 申烨华, 霍艳敏 (2009) 沙地濒危植物长柄扁桃仁中维生素E含量分析. 西北大学学报(自然科学版), 39, 777-779.] | |
| [13] | Li C, Li GP, Chen Q, Bai B, Shen YH, Zhang YL (2010) Fatty acid composition analysis of the seed oil of Amygdalus pedunculata Pall. China Oils and Fats, 35(4), 77-79. (in Chinese with English abstract) |
| [李聪, 李国平, 陈俏, 白斌, 申烨华, 张应龙 (2010) 长柄扁桃油脂肪酸成分分析. 中国油脂, 35(4), 77-79.] | |
| [14] | Li QY, Wang XP (2013) Elevational pattern of species richness in the Three Gorges region of the Yangtze River: effect of climate, geometric constraints, area and topographical heterogeneity. Biodiversity Science, 21, 141-152. (in Chinese with English abstract) |
| [李巧燕, 王襄平 (2013) 长江三峡库区物种多样性的垂直分布格局: 气候、几何限制、面积及地形异质性的影响. 生物多样性, 21, 141-152.] | |
| [15] | Lu Q, Wang JH, Chu JM (2012) Desert Plants in China. China Forestry Publishing House, Beijing. (in Chinese) |
| [卢琦, 王继和, 褚建民(2012) 中国荒漠植物图鉴. 中国林业出版社, 北京.] | |
| [16] | Ma Y, Ma RC (2004) Identification of genetic relationship of almond by AFLP. Journal of Fruit Science, 21, 552-555. (in Chinese with English abstract) |
| [马艳, 马荣才 (2004) 扁桃种质资源的AFLP分析. 果树学报, 21, 552-555.] | |
| [17] | McCune B, Grace J (2002) Analysis of Ecological Communities. Mjm Software Design, Glenden Beach, OR. |
| [18] | Moilanen A (2005) Reserve selection using nonlinear species distribution models. The American Naturalist, 165, 695-706. |
| [19] | Monserud RA, Leemans R (1992) Comparing global vegetation maps with the Kappa statistic. Ecological Modelling, 62, 275-293. |
| [20] | Özkan K, Mert A, Şentürk Ö (2011) Estimation of potential distribution of non-wood trading species richness using classification and regression tree technique: A case study from the Lakes District, Turkey. 2nd International Non- Wood Forest Products Symposium, Isparta, Turkey. |
| [21] | Phillips SJ, Miroslav K, Schapire RE (2004) A maximum entropy approach to species distribution modeling. Proceedings of the Twenty-First International Conference on Machine Learning, 95, 655-662. |
| [22] | Qiao HJ, Hu JH, Huang JH (2013) Theoretical basis, future directions, and challenges for ecological niche models. Scientia Sinica Vitae, 43, 915-927. (in Chinese with English abstract) |
| [乔慧捷, 胡军华, 黄继红 (2013) 生态位模型的理论基础、发展方向与挑战. 中国科学: 生命科学, 43, 915-927.] | |
| [23] | Qiao HJ, Soberón J, Peterson AT (2015) No silver bullets in correlative ecological niche modeling: insights from testing among many potential algorithms for niche estimation. Methods in Ecology and Evolution,6, 1126-1136. |
| [24] | Saupe EE, Qiao HJ, Hendricks JR, Portell RW, Hunter SJ, Soberón J, Lieberman BS (2015) Niche breadth and geographic range size as determinants of species survival on geological time scales. Global Ecology & Biogeography, 24, 1159-1169. |
| [25] | Stockwell D, Peters D (1999) The GARP modeling system: problems and solutions to automated spatial prediction. International Journal of Geographical Information Science, 13, 143-158. |
| [26] | Su GX (1997) Germplasm resources of almonds in China. China Seeds, 2, 6-8. |
| (in Chinese) [苏贵兴 (1987) 我国的扁桃种质资源. 作物品种资源, 2, 6-8.] | |
| [27] | van Proosdij ASJ, Sosef MSM, Wieringa JJ, Raes N (2015) Minimum required number of specimen records to develop accurate species distribution models. Ecography, 39, 542-552. |
| [28] | Wu B, Su ZZ, Chen ZX (2007) A revised potential extent of desertification in China. Journal of Desert Research, 27,911-917. (in Chinese with English abstract) |
| [吴波, 苏志珠, 陈仲新 (2007) 中国荒漠化潜在发生范围的修订. 中国沙漠, 27, 911-917.] | |
| [29] | Xu XQ, Chu JM (2013) Analysis on development potential and countermeasures of Amygdalus pedunculata industry. Forest Resources Management, 1, 22-25. (in Chinese with English abstract) |
| [许新桥, 褚建民 (2013) 长柄扁桃产业发展潜势分析及问题对策研究. 林业资源管理, 1, 22-25.] | |
| [30] | Xu XT, Yang Y, Wang LS (2008) Geographic distribution and potential distribution estimation of Pseudotaxus chienii. Journal of Plant Ecology (Chinese Version), 32,1134-1145. |
| [徐晓婷, 杨永, 王利松 (2008) 白豆杉的地理分布及潜在分布区估计. 植物生态学报, 32, 1134-1145.] | |
| [31] | Xu ZL, Peng HH, Peng SZ (2015) The development and evaluation of species distribution models. Acta Ecologica Sinica, 35, 557-567. |
| (in Chinese with English abstract) [许仲林, 彭焕华, 彭守章 (2015) 物种分布模型的发展及评价方法. 生态学报, 35, 557-567.] | |
| [32] | Zhang DY, Jiang XH (1999) Progress in studies of genetic diversity and conservation biology of endangered plant species. Chinese Biodiversity, 7, 31-37. (in Chinese with English abstract) |
| [张大勇, 姜新华 (1999) 遗传多样性与濒危植物保护生物学研究进展. 生物多样性, 7, 31-37.] | |
| [33] | Zhang L, Liu SR, Sun PS, Wang TL (2011) Comparative evaluation of the effects of climate change on the potential distribution of Pinus massoniana. Chinese Journal of Plant Ecology, 35, 1091-1105. |
| [张雷, 刘世荣, 孙鹏森, 王同立 (2011) 气候变化对马尾松潜在分布影响预估的多模型比较. 植物生态学报, 35, 1091-1105.] | |
| [34] | Zhang P, Shen YH, Wang XL, Hou RT (2007) Determination of amygdalin in Amygdalus pedunculata Pall by high performance liquid chromatography. Chinese Journal of Analysis Laboratory, 26(10), 80-83. (in Chinese with English abstract) |
| [张萍, 申烨华, 王晓玲, 侯睿婷 (2007) 高效液相色谱法测定长柄扁桃仁中的苦杏仁甙. 分析实验室, 26(10), 80-83.] | |
| [35] | Zhang WJ, Chen JK (2003) Advances in study of the distribution area of species. Biodiversity Science, 11, 364-369. (in Chinese with English abstract) |
| [张文驹, 陈家宽 (2003) 物种分布区研究进展. 生物多样性, 11, 364-369.] | |
| [36] | Zhang YB, Ma KP (2008) Geographic distribution patterns and status assessment of threatened plants in China. Biodiversity and Conservation, 17, 1783-1798. |
| [37] | Zhao YZ (1992) Atlas of Endangered Rare Plants in Inner Mongolia. China Agricultural Science and Technology Press, Beijing. (in Chinese) |
| [赵一之 (1992) 内蒙古珍稀濒危植物图谱. 中国农业科技出版社, 北京.] | |
| [38] | Zhu GP, Liu GQ, Bu WJ, Gao YB (2013) Ecological niche modeling and its applications in biodiversity conservation. Biodiversity Science, 21, 90-98. (in Chinese with English abstract) |
| [朱耿平, 刘国卿,卜文俊, 高玉葆 (2013) 生态位模型的基本原理及其在生物多样性保护中的应用. 生物多样性, 21, 90-98. ] | |
| [39] | Zuo WY, Lao N, Geng YY, Ma KP (2007) Predicting species’ potential distribution—SVM compared with GARP. Chinese Journal of Plant Ecology (Chinese Version), 31, 711-719. (in Chinese with English abstract) |
| [左闻韵, 劳逆, 耿玉英, 马克平 (2007) 预测物种潜在分布区——比较SVM与GARP. 植物生态学报, 31, 711-719.] |
| [1] | 易木荣, 卢萍, 彭勇, 汤勇, 许久恒, 尹浩萍, 张路杨, 翁晓东, 底明晓, 雷隽, 卢宸祺, 曹如君, 戴年华, 占德洋, 童媚, 楼智明, 丁永刚, 柴静, 车静. 北潦河金家水支流江西大鲵野外种群现状及栖息地评估[J]. 生物多样性, 2025, 33(4): 24145-. |
| [2] | 李沫潼, 何拓, 李薇, 廖菁, 曾岩. 从CITES的术语看野生动植物国际贸易监管规则[J]. 生物多样性, 2025, 33(4): 24545-. |
| [3] | 何拓, 曾岩, 殷亚方, 张坤, 袁良琛, 董晖, 周志华. 为野生植物保护和可持续贸易奠定科学基础——CITES植物委员会第27次会议评述[J]. 生物多样性, 2024, 32(9): 24390-. |
| [4] | 王艳丽, 张英, 戚春林, 张昌达, 史佑海, 杜彦君, 丁琼. 海南热带雨林国家公园生物多样性热点与保护空缺区域识别: 基于大型真菌与植物视角[J]. 生物多样性, 2024, 32(7): 24081-. |
| [5] | 董廷玮, 黄美玲, 韦旭, 马硕, 岳衢, 刘文丽, 郑佳鑫, 王刚, 马蕊, 丁由中, 薄顺奇, 王正寰. 上海地区金线侧褶蛙种群的潜在空间分布格局及其景观连通性[J]. 生物多样性, 2023, 31(8): 22692-. |
| [6] | 刘伟, 王濡格, 范天巧, 娜依曼·阿不都力江, 宋新航, 肖书平, 郭宁, 帅凌鹰. 福建省明溪县黑冠鹃隼生境适宜性[J]. 生物多样性, 2023, 31(7): 22660-. |
| [7] | 陈金锋, 吴欣静, 林海, 崔国发. 《国家重点保护野生动物名录》和其他保护名录对比分析[J]. 生物多样性, 2023, 31(6): 22639-. |
| [8] | 魏辰, 佟一杰, 曾岩, 白明, 万霞. 警惕物种致危和生物入侵的风险: 我国主要电商平台的甲虫贸易调查[J]. 生物多样性, 2023, 31(6): 22541-. |
| [9] | 鲍虞园, 李银康, 林吴颖, 周志琴, 肖晓波, 颉晓勇. 中国南海北部近海鲎资源调查及北部湾潮间带中华鲎幼鲎潜在栖息地评估[J]. 生物多样性, 2023, 31(5): 22407-. |
| [10] | 邓昶, 郝杰威, 高德, 任明迅, 张莉娜. 海南受威胁苔藓植物适生热点区域识别与保护[J]. 生物多样性, 2023, 31(4): 22580-. |
| [11] | 牛晓锋, 王晓梅, 张研, 赵志鹏, 樊恩源. 鲟鱼分子鉴定方法的整合应用[J]. 生物多样性, 2022, 30(6): 22034-. |
| [12] | 刘艳艳, 刘畅, 魏晓新. 我国及周边地区松属白松亚组系统学研究进展和保护现状[J]. 生物多样性, 2022, 30(2): 21344-. |
| [13] | 施雨含, 任宗昕, 王维嘉, 徐鑫, 刘杰, 赵延会, 王红. 中国-喜马拉雅三种黄耆属植物与其传粉熊蜂的空间分布预测[J]. 生物多样性, 2021, 29(6): 759-769. |
| [14] | 王文婷, 杨婷婷, 金磊, 蒋家民. 未来气候变化下两种红景天植物的脆弱性[J]. 生物多样性, 2021, 29(12): 1620-1628. |
| [15] | 袁海生, 魏玉莲, 周丽伟, 秦问敏, 崔宝凯, 何双辉. 东北4种林木干基腐朽病原真菌潜在分布范围预测及其生态位分析[J]. 生物多样性, 2019, 27(8): 873-879. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
备案号:京ICP备16067583号-7
Copyright © 2022 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn