
生物多样性 ›› 2018, Vol. 26 ›› Issue (10): 1103-1115. DOI: 10.17520/biods.2018159 cstr: 32101.14.biods.2018159
所属专题: 土壤生物与土壤健康
毕艳孟1,2, 孙振钧1,*( )
)
收稿日期:2018-06-01
接受日期:2018-08-03
出版日期:2018-10-20
发布日期:2019-01-06
通讯作者:
孙振钧
作者简介:# 共同第一作者
基金资助:
Yanmeng Bi1,2, Zhenjun Sun1,*()
Received:2018-06-01
Accepted:2018-08-03
Online:2018-10-20
Published:2019-01-06
Contact:
Sun Zhenjun
About author:# Co-first authors
摘要:
连作障碍不仅严重影响作物产量, 而且会导致土壤生物多样性下降、有益微生物减少及病原菌增加等一系列微生态失衡问题。土壤微生态失衡反作用于植物, 导致植物发生更严重的病害、减产等。作为土壤生态系统工程师, 蚯蚓的取食、掘洞和爬行等活动对土壤微生态具有重要的调控作用, 既可以改善土壤环境, 又可以强化土壤生物群落功能, 有望为缓解作物的连作障碍问题提供一条新途径。本文总结了土壤微生态与土壤功能维持及蚯蚓调控土壤生物作用的研究进展, 在此基础上, 结合蚯蚓对化感物质降解作用的研究, 分析了蚯蚓通过调控土壤微生态缓解作物连作障碍的微生物作用机制的三条途径: 直接调控微生物群落、通过改变化感物质组成以及通过调控土壤动物区系调控微生物群落。蚯蚓对微生物群落的调控可改善失衡的土壤根际微生态, 有效缓解作物连作障碍。
毕艳孟, 孙振钧 (2018) 蚯蚓调控土壤微生态缓解连作障碍的作用机制. 生物多样性, 26, 1103-1115. DOI: 10.17520/biods.2018159.
Yanmeng Bi, Zhenjun Sun (2018) Mechanisms of earthworms to alleviate continuous cropping obstacles through regulating soil microecology. Biodiversity Science, 26, 1103-1115. DOI: 10.17520/biods.2018159.
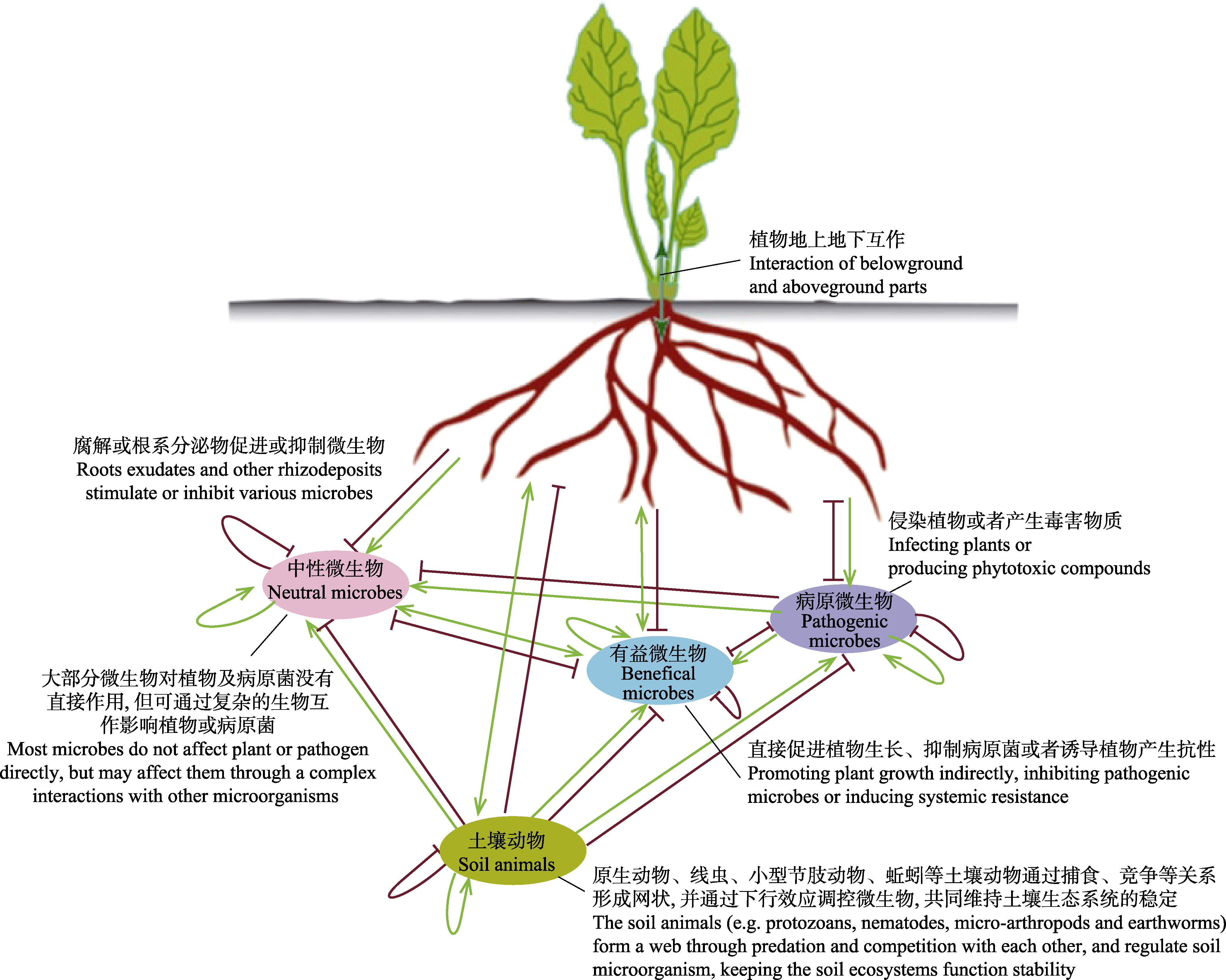
图1 基于各功能群相互作用的土壤微生态平衡(仿自Berendsen et al, 2012)。其中绿线加箭头表示促进作用, 红线加短线表示抑制作用。
Fig. 1 The balance of ecosystem (based on the interaction of functional groups) (modified from Berendsen et al, 2012). Green line indicates promoting effect and red line indicates inhibiting effect.
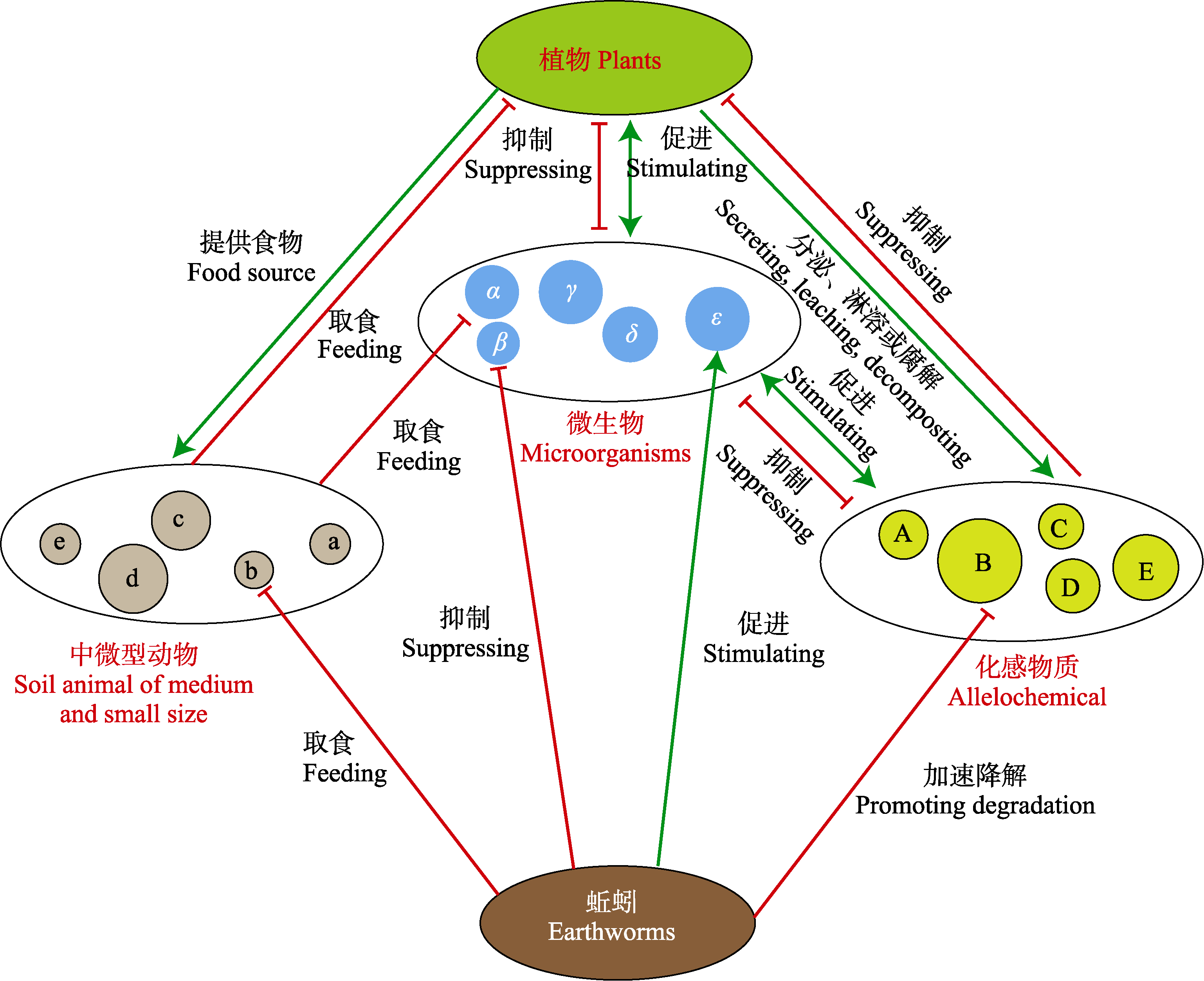
图2 蚯蚓调控连作障碍的土壤微生态机制
Fig. 2 The mechanism of earthworm regulating soil microecology
| [1] | Abbasi MK, Tahir MM, Sabir N, Khurshid M (2015) Impact of the addition of different plant residues on nitrogen mineralization-immobilization turnover and carbon content of a soil incubated under laboratory conditions. Solid Earth, 6, 197-205. |
| [2] | Akca I, Yilmaz NDK, Kizilkaya R (2014) Evaluation of suppression of rhizomania disease by earthworm (Lumbricus terrestris L.) and its effects on soil microbial activity in different sugar beet cultivars. Archives of Agronomy and Soil Science, 60, 1565-1575. |
| [3] | Andriuzzi WS, Phuong-Thi N, Geisen S, Keith AM, Dumack K, Bolger T, Bonkowski M, Brussaard L, Faber JH, Chabbi A, Rumpel C, Schmidt O (2016) Organic matter composition and the protist and nematode communities around anecic earthworm burrows. Biology and Fertility of Soils, 52, 91-100. |
| [4] | Angst Š, Mueller CW, Cajthaml T, Angst G, Lhotáková Z, Bartuška M, Špaldoňová A, Frouz J (2017) Stabilization of soil organic matter by earthworms is connected with physical protection rather than with chemical changes of organic matter. Geoderma, 289, 29-35. |
| [5] | Bais HP, Weir TL, Perry LG, Gilroy S, Vivanco JM (2006) The role of root exudates in rhizosphere interactions with plants and other organisms. Annual Review of Plant Biology, 57, 233-266. |
| [6] | Barbosa JZ, Demetrio WC, Silva CM, Dionísio JA (2017) Earthworms (Amynthas spp.) increase common bean growth, microbial biomass, and soil respiration. Semina: Ciências Agrárias, 38, 2887-2898. |
| [7] | Berendsen RL, Pieterse CM, Bakker PA (2012) The rhizosphere microbiome and plant health. Trends in Plant Science, 17, 478-486. |
| [8] | Berg G, Opelt K, Zachow C, Lottmann J, Gotz M, Costa R, Smalla K (2006) The rhizosphere effect on bacteria antagonistic towards the pathogenic fungus Verticillium differs depending on plant species and site. FEMS Microbiology Ecology, 56, 250-261. |
| [9] | Bi YM (2016) Effect and Mechanism for Earthworm to Remit Continuous Cropping Obstacle of Strawberry. PhD dissertation, China Agricultural University, Beijing. (in Chinese with English abstract) |
| [毕艳孟 (2016) 蚯蚓缓解草莓连作障碍的效应及机理研究.博士学位论文, 中国农业大学, 北京.] | |
| [10] | Bi YM, Tian GL, Wang C, Feng CL, Zhang Y, Zhang LS, Sun ZJ (2016) Application of leaves to induce earthworms to reduce phenolic compounds released by decomposing plants. European Journal of Soil Biology, 75, 31-37. |
| [11] | Bi YM, Tian GL, Wang C, Zhang Y, Wang DN, Zhang FF, Zhang LS, Sun ZJ (2018) Differential effects of two earthworm species on fusarium wilt of strawberry. Applied Soil Ecology, 126, 174-181. |
| [12] | Blouin M, Zuily-Fodil Y, Pham-Thi AT, Laffray D, Reversat G, Pando A, Tondoh J, Lavelle P (2005) Belowground organism activities affect plant aboveground phenotype, inducing plant tolerance to parasites. Ecology Letters, 8, 202-208. |
| [13] | Bonkowski M, Griffiths BS, Ritz K (2000) Food preferences of earthworms for soil fungi. Pedobiologia, 44, 666-676. |
| [14] | Bonkowski M, Schaefer M (1997) Interactions between earthworms and soil protozoa: A trophic component in the soil food web. Soil Biology & Biochemistry, 29, 499-502. |
| [15] | Boyer J, Reversat G, Lavelle P, Chabanne A (2013) Interactions between earthworms, and plant-parasitic nematodes. European Journal of Soil Biology, 59, 43-47. |
| [16] | Brussaard L, de Ruiter PC, Brown GG (2007) Soil biodiversity for agricultural sustainability. Agriculture Ecosystems & Environment, 121, 233-244. |
| [17] | Burtelow AE, Bohlen PJ, Groffman PM (1998) Influence of exotic earthworm invasion on soil organic matter, microbial biomass and denitrification potential in forest soils of the northeastern United States. Applied Soil Ecology, 9, 197-202. |
| [18] | Butenschoen O, Ji R, Schaffer A, Scheu S (2009) The fate of catechol in soil as affected by earthworms and clay. Soil Biology & Biochemistry, 41, 330-339. |
| [19] | Cao J, Wang C, Huang Y, Ji DG, Lou Y (2015) Effects of earthworm on soil microbes and biological fertility: A review. Chinese Journal of Applied Ecology, 26, 1579-1586. (in Chinese with English abstract) |
| [曹佳, 王冲, 皇彦, 纪丁戈, 楼屹 (2015) 蚯蚓对土壤微生物及生物肥力的影响研究进展. 应用生态学报, 26, 1579-1586.] | |
| [20] | Cao J, Wang C, Ji D (2016) Improvement of the soil nitrogen content and maize growth by earthworms and arbuscular mycorrhizal fungi in soils polluted by oxytetracycline. Science of the Total Environment, 571, 926-934. |
| [21] | Chapuis-Lardy L, Brauman A, Bernard L, Pablo AL, Toucet J, Mano MJ, Weber L, Brunet D, Razafimbelo T, Chotte JL, Blanchart E (2010) Effect of the endogeic earthworm Pontoscolex corethrurus on the microbial structure and activity related to CO2 and N2O fluxes from a tropical soil (Madagascar). Applied Soil Ecology, 45, 201-208. |
| [22] | Chen XY, Liu MQ, Hu F, Mao XF, Li HX (2007) Contributions of soil micro-fauna (protozoa and nematodes) to rhizosphere ecological functions. Acta Ecologica Sinica, 27, 3132-3143. (in Chinese with English abstract) |
| [陈小云, 刘满强, 胡锋, 毛小芳, 李辉信 (2007) 根际微型土壤动物——原生动物和线虫的生态功能. 生态学报, 27, 3132-3143.] | |
| [23] | Chung EJ, Hossain MT, Khan A, Kim KH, Jeon CO, Chung YR (2015) Bacillus oryzicola sp. nov., an endophytic bacterium isolated from the roots of rice with antimicrobial, plant growth promoting, and systemic resistance inducing activities in rice. Plant Pathology Journal, 31, 152-164. |
| [24] | Corno G, Jürgens K (2006) Direct and indirect effects of protist predation on population size structure of a bacterial strain with high phenotypic plasticity. Applied & Environmental Microbiology, 72, 78-86. |
| [25] | Dai JJ, Zhang C, Zhou B, Sun YT, Huang YT, Ren ZL, Dai J (2015) Effects of earthworm gut on microbial community structure in heavy metal contaminated soils. Journal of China Agricultural University, 20, 95-102. (in Chinese with English abstract) |
| [代金君,张池,周波,孙迎韬,黄钰婷,任宗玲,戴军 (2015) 蚯蚓肠道对重金属污染土壤微生物群落结构的影响. 中国农业大学学报, 20, 95-102.] | |
| [26] | Dempsey MA, Fisk MC, Fahey TJ (2011) Earthworms increase the ratio of bacteria to fungi in northern hardwood forest soils, primarily by eliminating the organic horizon. Soil Biology & Biochemistry, 43, 2135-2141. |
| [27] | Doran JW, Sarrantonio M, Liebig MA (1996) Soil health and sustainability. In: Advances in Agronomy (ed. Sparks DL), pp. 1-54. Elsevier Academic Press Inc., San Diego. |
| [28] | Doran JW, Zeiss MR (2000) Soil health and sustainability: Managing the biotic component of soil quality. Applied Soil Ecology, 15, 3-11. |
| [29] | Drake HL, Horn MA (2007) As the worm turns: The earthworm gut as a transient habitat for soil microbial biomes. Annual Review of Microbiology, 61, 169-189. |
| [30] | Du XF, Li YB, Liu F, Su XL, Li Q (2018) Structure and ecological functions of soil micro-food web. Chinese Journal of Applied Ecology, 29, 403-411. (in Chinese with English abstract) |
| [杜晓芳, 李英滨, 刘芳, 宿晓琳, 李琪 (2018) 土壤微食物网结构与生态功能. 应用生态学报, 29, 403-411.] | |
| [31] | Edwards CA, Bohlen PJ (1996) Biology and Ecology of Earthworm, 3rd edn. Chapman and Hall, Springer, London. |
| [32] | Edwards CA, Fletcher KE (1988) Interactions between earthworms and microorganisms in organic-matter breakdown. Agriculture Ecosystems & Environment, 24, 235-247. |
| [33] | Eisenhauer N, Schlaghamerský J, Reich PB, Frelich LE (2011) The wave towards a new steady state: Effects of earthworm invasion on soil microbial functions. Biological Invasions, 13, 2191-2196. |
| [34] | Elmer WH (2009) Influence of earthworm activity on soil microbes and soilborne diseases of vegetables. Plant Disease, 93, 175-179. |
| [35] | Elmer WH, Ferrandino FJ (2009) Suppression of Verticillium wilt of eggplant by earthworms. Plant Disease, 93, 485-489. |
| [36] | Fang XL, Kuo J, You MP, Finnegan PM, Barbetti MJ (2012) Comparative root colonisation of strawberry cultivars Camarosa and Festival by Fusarium oxysporum f. sp. fragariae. Plant and Soil, 358, 71-85. |
| [37] | Gao MX, Taylor MK, Callaham MA (2017) Trophic dynamics in a simple experimental ecosystem: Interactions among centipedes, Collembola and introduced earthworms. Soil Biology & Biochemistry, 115, 66-72. |
| [38] | Garbeva P, van Veen JA, van Elsas JD (2004) Microbial diversity in soil: Selection microbial populations by plant and soil type and implications for disease suppressiveness. Annual Review of Phytopathology, 42, 243-270. |
| [39] | Geisen S, Koller R, Hünninghaus M, Dumack K, Urich T, Bonkowski M (2016) The soil food web revisited: Diverse and widespread mycophagous soil protists. Soil Biology & Biochemistry, 94, 10-18. |
| [40] | Gomez-Brandon M, Aira M, Lores M, Dominguez J (2011) Epigeic earthworms exert a bottleneck effect on microbial communities through gut associated processes. PLoS ONE, 6, e24786. |
| [41] | González G, Seastedt TR (2001) Soil fauna and plant litter decomposition in tropical and subalpine forests. Ecology, 82, 955-964. |
| [42] | Gopalakrishnan S, Pande S, Sharma M, Humayun P, Kiran BK, Sandeep D, Vidya MS, Deepthi K, Rupela O (2011) Evaluation of actinomycete isolates obtained from herbal vermicompost for the biological control of Fusarium wilt of chickpea. Crop Protection, 30, 1070-1078. |
| [43] | Gyaneshwar P, Kumar GN, Parekh LJ, Poole PS (2003) Role of soil microorganisms in improving P nutrition of plants. System Sciences & Comprehensive Studies in Agriculture, 245, 83-93. |
| [44] | He JZ, Ge Y, Xu ZH, Chen CR (2009) Linking soil bacterial diversity to ecosystem multifunctionality using backward-elimination boosted trees analysis. Journal of Soils and Sediments, 9, 547-554. |
| [45] | Hu YX, Sun ZJ, Zhou FY, Liu XY, Cao AC (2002) Study on suppressness effect of vermicompost to soil-borne disease of cucumber seedlings. Acta Ecologica Sinica, 22, 1106-1115. (in Chinese with English abstract) |
| [胡艳霞, 孙振钧, 周法永, 刘小秧, 曹坳程 (2002) 蚯蚓粪对黄瓜苗期土传病害的抑制作用. 生态学报, 22, 1106-1115.] | |
| [46] | Huang K, Li FS, Wei YF, Chen XM, Fu XY (2013a) Changes of bacterial and fungal community compositions during vermicomposting of vegetable wastes by Eisenia foetida. Bioresource Technology, 150, 235-241. |
| [47] | Huang LF, Song LX, Xia XJ, Mao WH, Shi K, Zhou YH, Yu JQ (2013b) Plant-soil feedbacks and soil sickness: From mechanisms to application in agriculture. Journal of Chemical Ecology, 39, 232-242. |
| [48] | Hume EA, Horrocks AJ, Fraser PM, Curtin D, Meenken ED, Chng S, Beare MH (2015) Alleviation of take-all in wheat by the earthworm Aporrectodea caliginosa (Savigny). Applied Soil Ecology, 90, 18-25. |
| [49] | Ilieva-Makulec K, Makulec G (2002) Effect of the earthworm Lumbricus rubellus on the nematode community in a peat meadow soil. European Journal of Soil Biology, 38, 59-62. |
| [50] | Janvier C, Villeneuve F, Alabouvette C, Edel-Hermann V, Mateille T, Steinberg C (2007) Soil health through soil disease suppression: Which strategy from descriptors to indicators? Soil Biology & Biochemistry, 39, 1-23. |
| [51] | Jilani G, Mahmood S, Chaudhry AN, Hassan I, Akram M (2008) Allelochemicals: Sources, toxicity and microbial transformation in soil—A review. Annals of Microbiology, 58, 351-357. |
| [52] | Jing ZW, Chen RR, Wei SP, Feng YZ, Zhang JB, Lin XG (2017) Response and feedback of C mineralization to P availability driven by soil microorganisms. Soil Biology & Biochemistry, 105, 111-120. |
| [53] | Jones CG, Lawton JH, Shachak M (1994) Organisms as ecosystem engineers. Oikos, 69, 373-386. |
| [54] | Kim J-S, Lee J, Lee C-H, Woo SY, Kang H, Seo S-G, Kim S-H (2015) Activation of Pathogenesis-related genes by the rhizobacterium, Bacillus sp. JS, which induces systemic resistance in tobacco plants. Plant Pathology Journal, 31, 195-201. |
| [55] | Koike ST, Kirkpatrick SC, Gordon TR (2009) Fusarium wilt of strawberry caused by Fusarium oxysporum in California. Plant Disease, 93, 1077. |
| [56] | Kulmatiski A, Beard KH, Stevens JR, Cobbold SM (2008) Plant-soil feedbacks: A meta-analytical review. Ecology Letters, 11, 980-992. |
| [57] | Lambers H, Mougel C, Jaillard B, Hinsinger P (2009) Plant_microbe_soil interactions in the rhizosphere: An evolutionary perspective. Plant and Soil, 321, 83-115. |
| [58] | Larkin RP (2015) Soil health paradigms and implications for disease management. Annual Review of Phytopathology, 53, 199-221. |
| [59] | Lee KE (1985) Earthworms,Their Ecology and Relationships with Soils and Land Use,Academic Press, , Sydney. |
| [60] | Li H, Du ZY, Liu Q, Shi YX (2016) Effect of earthworm-mycorrhiza interaction on soil enzyme activities, root growth and nutrients uptake of sweet potato. Journal of Plant Nutrition & Fertilizer, 22, 209-215. (in Chinese with English abstract) |
| [李欢, 杜志勇, 刘庆, 史衍玺 (2016) 蚯蚓菌根互作对土壤酶活、甘薯根系生长及养分吸收的影响. 植物营养与肥料学报, 22, 209-215.] | |
| [61] | Li HQ, Liu QZ, Zhang LL, Wang YL, Zhang H, Bai PH, Luan XB (2014) Accumulation of phenolic acids in the monocultured strawberry soils and their effect on soil nematodes. Chinese Journal of Ecology, 33, 169-175. (in Chinese with English abstract) |
| [李贺勤, 刘奇志, 张林林, 王玉玲, 张航, 白鹏华, 栾小兵 (2014) 草莓连作土壤酚酸类物质积累对土壤线虫的影响. 生态学杂志, 33, 169-175.] | |
| [62] | Li X, Fisk MC, Fahey TJ, Bohlen PJ (2002) Influence of earthworm invasion on soil microbial biomass and activity in a northern hardwood forest. Soil Biology & Biochemistry, 34, 1929-1937. |
| [63] | Li XG, Ding CF, Zhang TL, Wang XX (2014) Fungal pathogen accumulation at the expense of plant-beneficial fungi as a consequence of consecutive peanut monoculturing. Soil Biology & Biochemistry, 72, 11-18. |
| [64] | Lipiec J, Frąc M, Brzezińska M, Turski M, Oszust K (2016) Linking microbial enzymatic activities and functional diversity of soil around earthworm burrows and casts. Frontiers in Microbiology, 7, 1361. |
| [65] | Liu X, Zhang JL, Gu TY, Zhang WM, Shen QR, Yin SX, Qiu HZ (2014) Microbial community diversities and taxa abundances in soils along a seven-year gradient of potato monoculture using high throughput pyrosequencing approach. PLoS ONE, 9, e86610. |
| [66] | Loranger-Merciris G, Cabidoche YM, Deloné B, Quénéhervé P, Ozier-Lafontaine H (2012) How earthworm activities affect banana plant response to nematodes parasitism. Applied Soil Ecology, 52, 1-8. |
| [67] | Marinissen JCY, Bok J (1988) Earthworm-amended soil structure: Its influence on Collembola population in grassland. Pedobiologia, 32, 243-252. |
| [68] | Mclean MA, Parkinson D (1998) Impacts of the epigeic earthworm Dendrobaena octaedra on oribatid mite community diversity and microarthropod abundances in pine forest floor: A mesocosm study. Applied Soil Ecology, 7, 125-136. |
| [69] | Mohan KV, Hrushikesh N, Sreehari K, Kumar TA, Vidyavathi N, Pallavi A (2011) Studies on bioremediation of phenol by earthworm. International Journal of Environmental Sciences, 1, 1268-1273. |
| [70] | Moore JC, Zwetsloot HJC, Ruiter PCD (1990) Statistical analysis and simulation modelling of the belowground food webs of two winter wheat management practices. Netherlands Journal of Agricultural Science, 38, 303-316. |
| [71] | Mota FC, Alves GCS, Giband M, Gomes ACMM, Sousa FR, Mattos VS, Barbosa VHS, Barroso PAV, Nicole M, Peixoto JR, Rocha MR, Carneiro RMDG (2013) New sources of resistance to Meloidogyne incognita race 3 in wild cotton accessions and histological characterization of the defence mechanisms. Plant Pathology, 62, 1173-1183. |
| [72] | Nannipieri P, Ascher J, Ceccherini MT, Landi L, Pietramellara G, Renella G (2010) Microbial diversity and soil functions. European Journal of Soil Science, 54, 655-670. |
| [73] | Natal-da-Luz T, Lee I, Verweij RA, Morais PV, Van Velzen MJ, Sousa JP, Van Gestel CA (2012) Influence of earthworm activity on microbial communities related with the degradation of persistent pollutants. Environmental Toxicology and Chemistry, 31, 794-803. |
| [74] | Niu XQ, Li JH, Zhang JL, Shen BY, Chai ZX, Wang D (2011) Changes of Fusarium in rhizosphere soil under potato continuous cropping systems in arid-irrigated area of Gansu Province. Acta Prataculturae Sinica, 20, 236-243. |
| [75] | Oka Y (2010) Mechanisms of nematode suppression by organic soil amendments—A review. Applied Soil Ecology, 44, 101-115. |
| [76] | Oldenburg E, Kramer S, Schrader S, Weinert J (2008) Impact of the earthworm Lumbricus terrestris on the degradation of Fusarium-infected and deoxynivalenol-contaminated wheat straw. Soil Biology & Biochemistry, 40, 3049-3053. |
| [77] | Salmon S, Ponge JF (2001) Earthworm excreta attract soil springtails: Laboratory experiments on Heteromurus nitidus (Collembola: Entomobryidae). Soil Biology & Biochemistry, 33, 1959-1969. |
| [78] | Sampedro L, Whalen JK (2007) Changes in the fatty acid profiles through the digestive tract of the earthworm Lumbricus terrestris L. Applied Soil Ecology, 35, 226-236. |
| [79] | Sauvadet M, Chauvat M, Cluzeau D, Maron PA, Villenave C, Bertrand I (2016) The dynamics of soil micro-food web structure and functions vary according to litter quality. Soil Biology & Biochemistry, 95, 262-274. |
| [80] | Schaefer M, Petersen SO, Filser J (2005) Effects of Lumbricus terrestris, Allolobophora chlorotica and Eisenia fetida on microbial community dynamics in oil-contaminated soil. Soil Biology & Biochemistry, 37, 2065-2076. |
| [81] | Schrader S, Wolfarth F, Oldenburg E (2013) Biological control of soil-borne phytopathogenic fungi and their mycotoxins by soil fauna—A review. Bulletin of the University of Agricultural Sciences & Veterinary, 70, 291-298. |
| [82] | Shan J, Liu J, Wang YF, Yan XY, Guo HY, Li XZ, Ji R (2013) Digestion and residue stabilization of bacterial and fungal cells, protein, peptidoglycan, and chitin by the geophagous earthworm Metaphire guillelmi. Soil Biology & Biochemistry, 64, 9-17. |
| [83] | Shi LB, Wang ZH, Wu HY, Liu J (2010) Influence of continuous tomato-cropping on second-stage juveniles of root-knot nematode and free-living nematodes from rhizosphere soil in plastic greenhouse. Acta Phytopathologica Sinica, 40, 81-89. (in Chinese with English abstract) |
| [时立波, 王振华, 吴海燕, 刘静 (2010) 连作年限对番茄根围土壤根结线虫二龄幼虫与自由生活线虫数量的影响. 植物病理学报, 40, 81-89.] | |
| [84] | Singh JS, Raghubanshi AS, Singh RS, Srivastava SC (1989) Microbial biomass acts as a source of plant nutrients in dry tropical forest and savanna. Nature, 338, 499-500. |
| [85] | Song YY, Zeng RS, Xu JF, Li J, Shen X, Yihdego WG (2010) Interplant communication of tomato plants through underground common mycorrhizal networks. PLoS ONE, 5, e13324. |
| [86] | Tao J, Chen XY, Liu MQ, Hu F, Griffiths B, Li HX (2009) Earthworms change the abundance and community structure of nematodes and protozoa in a maize residue amended rice-wheat rotation agro-ecosystem. Soil Biology & Biochemistry, 41, 898-904. |
| [87] | Tao J, Xu YJ, Griffiths BS, Hu F, Chen XY, Jiao JG, Li HX (2011) Earthworms reduce the abundance of nematodes and enchytraeids in a soil mesocosm experiment despite abundant food resources. Soil Science Society of America Journal, 75, 1774-1778. |
| [88] | Teste FP, Kardol P, Turner BL, Wardle DA, Zemunik G, Renton M, Laliberte E (2017) Plant-soil feedback and the maintenance of diversity in Mediterranean-climate shrublands. Science, 355, 173-176. |
| [89] | Thakuria D, Schmidt O, Finan D, Egan D, Doohan FM (2010) Gut wall bacteria of earthworms: A natural selection process. The ISME Journal, 4, 357-366. |
| [90] | Toljander JF, Lindahl BD, Paul LR, Elfstrand M, Finlay RD (2007) Influence of arbuscular mycorrhizal mycelial exudates on soil bacterial growth and community structure. FEMS Microbiology Ecology, 61, 295-304. |
| [91] | Vaitauskiene K, Sarauskis E, Naujokiene V, Liakas V (2015) The influence of free-living nitrogen-fixing bacteria on the mechanical characteristics of different plant residues under no-till and strip-till conditions. Soil & Tillage Research, 154, 91-102. |
| [92] | Watkins AJ, Nicol GW, Shaw LJ (2009) Use of an artificial root to examine the influence of 8-hydroxyquinoline on soil microbial activity and bacterial community structure. Soil Biology & Biochemistry, 41, 580-585. |
| [93] | Wolfarth F, Schrader S, Oldenburg E, Weinert J (2011) Contribution of the endogeic earthworm species Aporrectodea caliginosa to the degradation of deoxynivalenol and Fusarium biomass in wheat straw. Mycotoxin Research, 27, 215-220. |
| [94] | Wu FY, Wan JHC, Wu SC, Wong MH (2012) Effects of earthworms and plant growth-promoting rhizobacteria (PGPR) on availability of nitrogen, phosphorus, and potassium in soil. Journal of Plant Nutrition and Soil Science, 175, 423-433. |
| [95] | Wu JH, Song CY, Chen JK (2007) Effect of microbivorous nematodes on plant growth and soil nutrient cycling: A review. Biodiversity Science, 15, 124-133. (in Chinese with English abstract) |
| [吴纪华, 宋慈玉, 陈家宽 (2007) 食微线虫对植物生长及土壤养分循环的影响. 生物多样性, 15, 124-133.] | |
| [96] | Wu YP, Lü LY, Bi YM, Zhang Y, Sun ZJ (2013) Effects of earthworm inoculation on saline-alkali soil nutrient, soil organisms and plant cultivation. Journal of China Agricultural University, 18, 45-51. (in Chinese with English abstract) |
| [伍玉鹏, 吕丽媛, 毕艳孟, 张一, 孙振钧 (2013) 接种蚯蚓对盐碱土养分、土壤生物及植被的影响. 中国农业大学学报, 18, 45-51.] | |
| [97] | Wu YP, Muhammad S, Zhao JS, Hao R, Hu RG (2015) Effect of the earthworm gut-stimulated denitrifiers on soil nitrous oxide emissions. European Journal of Soil Biology, 70, 104-110. |
| [98] | Wu ZW, Wang MD, Liu XY, Chen HG, Jia XC (2009) Phenolic compounds accumulation in continuously cropped Rehmannia glutinosa soil and their effects on R. glutinosa growth. Chinese Journal of Ecology, 28, 660-664. (in Chinese with English abstract) |
| [吴宗伟, 王明道, 刘新育, 陈红歌, 贾新成 (2009) 重茬地黄土壤酚酸的动态积累及其对地黄生长的影响. 生态学杂志, 28, 660-664.] | |
| [99] | Wurst S, De Devn GB, Orwin K (2013) Soil biodiversity and functions. In: Soil Ecology and Ecosystem Services (eds Wall DH, Bardgett RD, Behan-Pelletier V, Herrick JE, Jones TH, Six J, Strong DR, van der Putten WH), pp. 28-45. Oxford University Press, Oxford. |
| [100] | Yahaya ANA, Hossain MS, Edyvean R (2017) Analysis of phenolic compounds in empty fruit bunches in oyster mushroom cultivation and in vermicomposting. BioResource, 12, 4594-4605. |
| [101] | Yao Y, He J, Zhang Y, Li YF, Wu YP (2015) Effects of Eisenia fetida inoculation on soil organic carbon and soil microorganisms under rice straw application. Journal of Agro-Environment Science, 34, 110-117. (in Chinese with English abstract) |
| [姚影, 何静, 张一, 李钰飞, 伍玉鹏 (2015) 赤子爱胜蚓(Eisenia fetida)对秸秆施入后土壤有机碳和微生物的影响. 农业环境科学学报, 34, 110-117.] | |
| [102] | Ye SF, Yu JQ, Peng YH, Zheng JH, Zou LY (2004) Incidence of Fusarium wilt in Cucumis sativus L. is promoted by cinnamic acid, an autotoxin in root exudates. Plant and Soil, 263, 143-150. |
| [103] | Zhang BG, Li GT, Shen TS, Wang JK, Sun Z (2000) Changes in microbial biomass C, N, and P and enzyme activities in soil incubated with the earthworms Metaphire guillelmi or Eisenia fetida. Soil Biology & Biochemistry, 32, 2055-2062. |
| [104] | Zhang N, Wang DD, Liu YP, Li SQ, Shen QR, Zhang RF (2014) Effects of different plant root exudates and their organic acid components on chemotaxis, biofilm formation and colonization by beneficial rhizosphere-associated bacterial strains. Plant and Soil, 374, 689-700. |
| [105] | Zhang WW, Cao J, Zhang SD, Wang C (2016) Effect of earthworms and arbuscular mycorrhizal fungi on the microbial community and maize growth under salt stress. Applied Soil Ecology, 107, 214-223. |
| [106] | Zhang WX, Chen DM, Zhao CC (2007) Functions of earthworm in ecosystem. Biodiversity Science, 15, 142-153. (in Chinese with English abstract) |
| [张卫信, 陈迪马, 赵灿灿 (2007) 蚯蚓在生态系统中的作用. 生物多样性, 15, 142-153.] | |
| [107] | Zhang XP, Hou WL, Chen P (2001) Soil animal guilds and their ecological distribution in the northeast of China. Chinese Journal of Applied & Environmental Biology, 7, 370-374. (in Chinese with English abstract) |
| [张雪萍, 侯威岭, 陈鹏 (2001) 东北森林土壤动物同功能种团及其生态分布. 应用与环境生物学报, 7, 370-374.] | |
| [108] | Zhang Y, Wang GC, Wu YP, Zhao H, Zhang YF, Sun ZJ (2013) PCR-DGGE analysis of earthworm gut bacteria diversity in stress of Escherichia coli O157: H7. Advances in Bioscience and Biotechnology, 4, 437-441. |
| [109] | Zhang Y, Zhang DJ, Li X, Zhang J (2018) Contribution of soil fauna to the degradation of recalcitrant components in Cinnamomum camphora foliar litter in different-sized gaps in Pinus massoniana plantations. Journal of Forestry Research. https://doi.org/10.1007/s11676-018-0609-6. |
| [110] | Zhen WC, Dai L, Hu TL, Cao KQ (2005) Study on dynamics of soil microorganisms under strawberry (Fragria ananassa Duch) continuous cropping. Journal of Agricultural University of Hebei, 28, 70-72, 87. (in Chinese with English abstract) |
| [甄文超, 代丽, 胡同乐, 曹克强 (2005) 连作草莓土壤微生物区系动态的研究. 河北农业大学学报, 28, 70-72, 87.] | |
| [111] | Zheng XQ, Fan XF, Zhang HL, Li SX, Wang JQ, Zhang JQ, Wang LJ, Tao XB, Lü WG (2015) Effects of Pheretima guillelmi cultivation time on microbial community diversity and characteristics of carbon metabolism in vegetable soil. Journal of Agricultural Resources & Environment, 32, 596-602. (in Chinese with English abstract) |
| [郑宪清,范晓芬,张翰林,李双喜,王金庆,张娟琴,王良军,陶晓斌,吕卫光 (2015) 威廉环毛蚓耕作时间对菜田土壤微生物群落多样性及碳代谢特征的影响. 农业资源与环境学报, 32, 596-602.] | |
| [112] | Zhu XY, Chang L, Liu J, Zhou MH, Li JJ, Gao B, Wu DH (2016) Exploring the relationships between soil fauna, different tillage regimes and CO2 and N2O emissions from black soil in China. Soil Biology & Biochemistry, 103, 106-116. |
| [113] | Zhu YH, Li KZ, Lu L (2012) Rhizosphere soil fauna and its effects on plant growth: A review. Chinese Journal of Ecology, 31, 2688-2693. (in Chinese with English abstract) |
| [朱永恒, 李克中, 陆林 (2012) 根际土壤动物及其对植物生长的影响. 生态学杂志, 31, 2688-2693.] |
| [1] | 宋威, 程才, 王嘉伟, 吴纪华. 土壤微生物对植物多样性–生态系统功能关系的调控作用[J]. 生物多样性, 2025, 33(4): 24579-. |
| [2] | 刘淑琪, 崔东, 江智诚, 刘江慧, 闫江超. 短期氮、水添加和刈割减弱了苦豆子型退化草地土壤生物多样性与生态系统多功能性的联系[J]. 生物多样性, 2025, 33(3): 24305-. |
| [3] | 王嘉陈, 徐汤俊, 许唯, 张高季, 尤艺瑾, 阮宏华, 刘宏毅. 城市景观格局对大蚰蜒种群遗传结构的影响[J]. 生物多样性, 2025, 33(1): 24251-. |
| [4] | 孙怡欣, 侯春雨, 周磊, 魏雪, 马金豪, 薛娟, 李小涵, 吴鹏飞. 青藏高原盆栽一年生和多年生豆科牧草对土壤线虫群落的影响[J]. 生物多样性, 2024, 32(7): 24040-. |
| [5] | 马骅, 李常青, 余品锋, 陈杰, 贺天耀, 王可洪. 澎溪河消落带大型土壤动物群落分布格局及其影响因素[J]. 生物多样性, 2024, 32(7): 24117-. |
| [6] | 李雪萌, 蒋际宝, 张曾鲁, 刘晓静, 王亚利, 吴宜钊, 李银生, 邱江平, 赵琦. 宝天曼国家级自然保护区蚯蚓物种多样性及其影响因素[J]. 生物多样性, 2024, 32(4): 23352-. |
| [7] | 王党军, 谢午阳, 林小元, 乔秀娟, 徐耀粘, 田秋香, 刘峰, 张娅妮, 左娟, 江明喜. 八大公山森林土壤动物群落与叶经济谱及凋落物分解速率的关系[J]. 生物多样性, 2024, 32(12): 24261-. |
| [8] | 朱晓华, 高程, 王聪, 赵鹏. 尿素对土壤细菌与真菌多样性影响的研究进展[J]. 生物多样性, 2023, 31(6): 22636-. |
| [9] | 沈诗韵, 潘远飞, 陈丽茹, 土艳丽, 潘晓云. 喜旱莲子草原产地和入侵地种群的植物-土壤反馈差异[J]. 生物多样性, 2023, 31(3): 22436-. |
| [10] | 杨预展, 余建平, 钱海源, 陈小南, 陈声文, 袁志林. 钱江源国家公园体制试点区水稻田土壤微生物群落的格局及其驱动机制[J]. 生物多样性, 2023, 31(2): 22392-. |
| [11] | 赵雯, 王丹丹, 热依拉·木民, 黄开钏, 刘顺, 崔宝凯. 阿尔山地区兴安落叶松林土壤微生物群落结构[J]. 生物多样性, 2023, 31(2): 22258-. |
| [12] | 谢致敬, 刘相钰, 孙晓铭, 刘继亮, 刘占锋, 张晓珂, 陈军, 杨效东, 朱波, 柯欣, 吴东辉. 中国土壤动物多样性监测网络建设、进展与展望[J]. 生物多样性, 2023, 31(12): 23365-. |
| [13] | 闫冰, 陆晴, 夏嵩, 李俊生. 城市土壤微生物多样性研究进展[J]. 生物多样性, 2022, 30(8): 22186-. |
| [14] | 李季蔓, 靳楠, 胥毛刚, 霍举颂, 陈小云, 胡锋, 刘满强. 不同干旱水平下蚯蚓对番茄抗旱能力的影响[J]. 生物多样性, 2022, 30(7): 21488-. |
| [15] | 程建伟, 王亚东, 王桠楠, 李莹, 郭颖, 白正, 刘新民, 李永宏. 半干旱草原大中型土壤动物在畜粪分解中的作用[J]. 生物多样性, 2022, 30(12): 22575-. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
备案号:京ICP备16067583号-7
Copyright © 2022 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn