
生物多样性 ›› 2014, Vol. 22 ›› Issue (5): 596-607. DOI: 10.3724/SP.J.1003.2014.14005 cstr: 32101.14.SP.J.1003.2014.14005
郑智, 龚大洁*( ), 孙呈祥, 李晓军, 李万江
), 孙呈祥, 李晓军, 李万江
收稿日期:2014-01-07
接受日期:2014-08-13
出版日期:2014-09-20
发布日期:2014-10-09
通讯作者:
龚大洁
基金资助:
Zhi Zheng, Dajie Gong*(), Chengxiang Sun, Xiaojun Li, Wanjiang Li
Received:2014-01-07
Accepted:2014-08-13
Online:2014-09-20
Published:2014-10-09
Contact:
Gong Dajie
摘要:
物种多样性和种域宽度沿环境梯度的分布格局及其成因机制一直是生物地理学和生态学讨论的重要议题。本研究采用多元回归模型和方差分离的方法判断面积、水分和能量、边界限制对秦岭两栖、爬行动物及其不同区系成分的物种丰富度海拔梯度分布格局的影响。结果表明, 秦岭两栖爬行动物及其不同区系成分的物种丰富度均呈单峰分布格局, 但峰值分布的海拔段有所差异。形成这种格局是各种因素相互作用的结果, 3种假设的独立解释力较低。水分能量动态假设对两栖、爬行动物物种的丰富度格局有很强的解释能力, 但水分和能量的解释力中有很大一部分属于边界限制、面积的协同作用, 在解释两栖动物的海拔分布格局时, 边界限制与水分和能量之间存在较强的共线性, 而在解释爬行动物的海拔分布格局时, 面积与水分和能量之间存在较强的共线性。同时, 本研究采用Stevens法和逐种法对Rapoport法则进行了验证。结果表明, 爬行动物物种种域的海拔梯度格局基本上支持Rapoport法则, 两栖动物很难判断是否支持Rapoport法则。
郑智, 龚大洁, 孙呈祥, 李晓军, 李万江 (2014) 秦岭两栖、爬行动物物种多样性海拔分布格局及其解释. 生物多样性, 22, 596-607. DOI: 10.3724/SP.J.1003.2014.14005.
Zhi Zheng, Dajie Gong, Chengxiang Sun, Xiaojun Li, Wanjiang Li (2014) Elevational pattern of amphibian and reptile diversity in Qinling Range and explanation. Biodiversity Science, 22, 596-607. DOI: 10.3724/SP.J.1003.2014.14005.
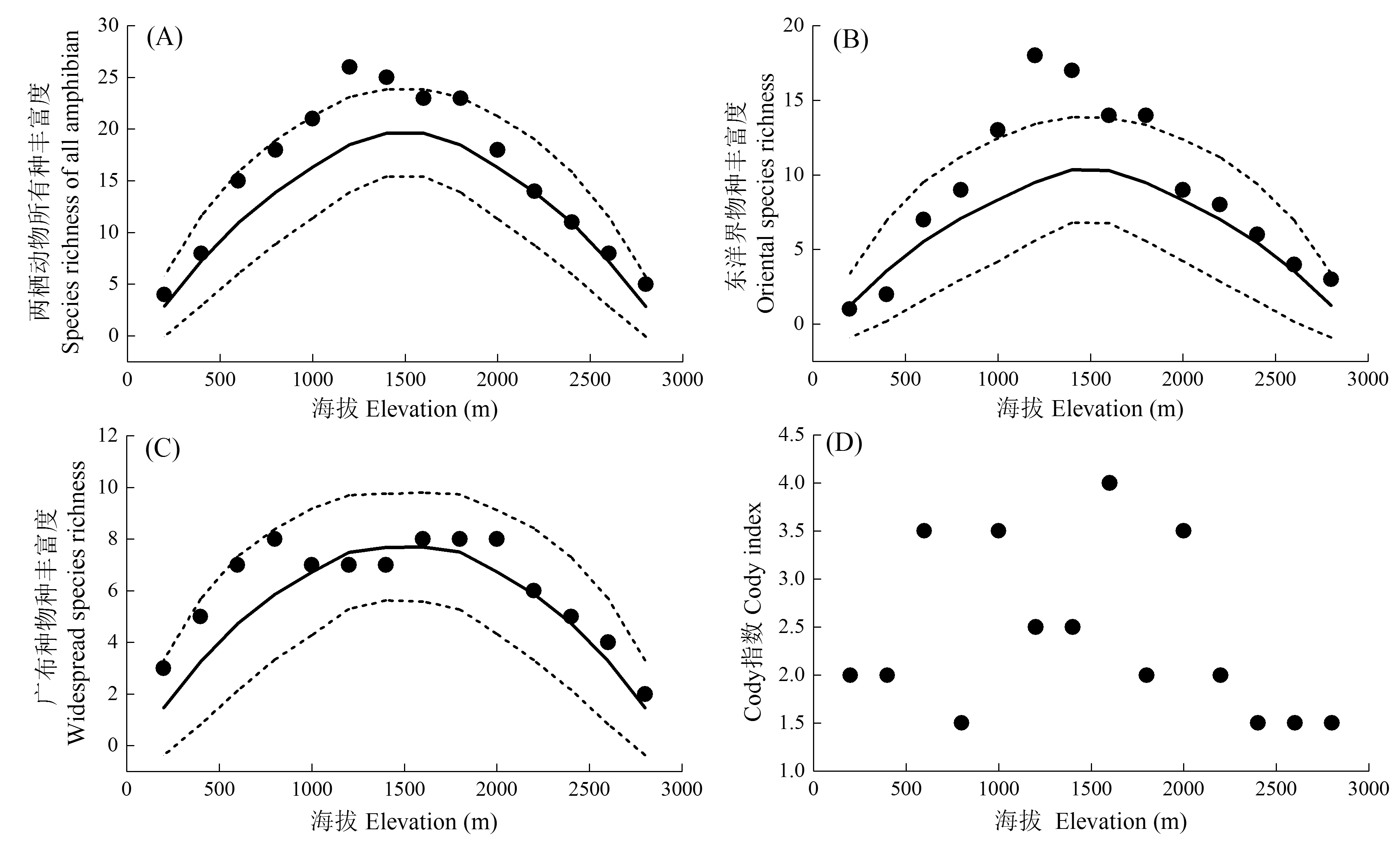
图1 两栖动物及其不同区系成分的物种多样性海拔分布格局。实心点表示实际的物种丰富度, 实线为中域效应模型预测的物种丰富度, 虚线表示模型预测的95%置信区间。
Fig. 1 Elevational patterns of species diversity of amphibians and different faunal components. Dot represents observed richness; solid line represents the predicted richness by mid-domain effect model, while dashed lines represent the 95% confident intervals of predicted richness.
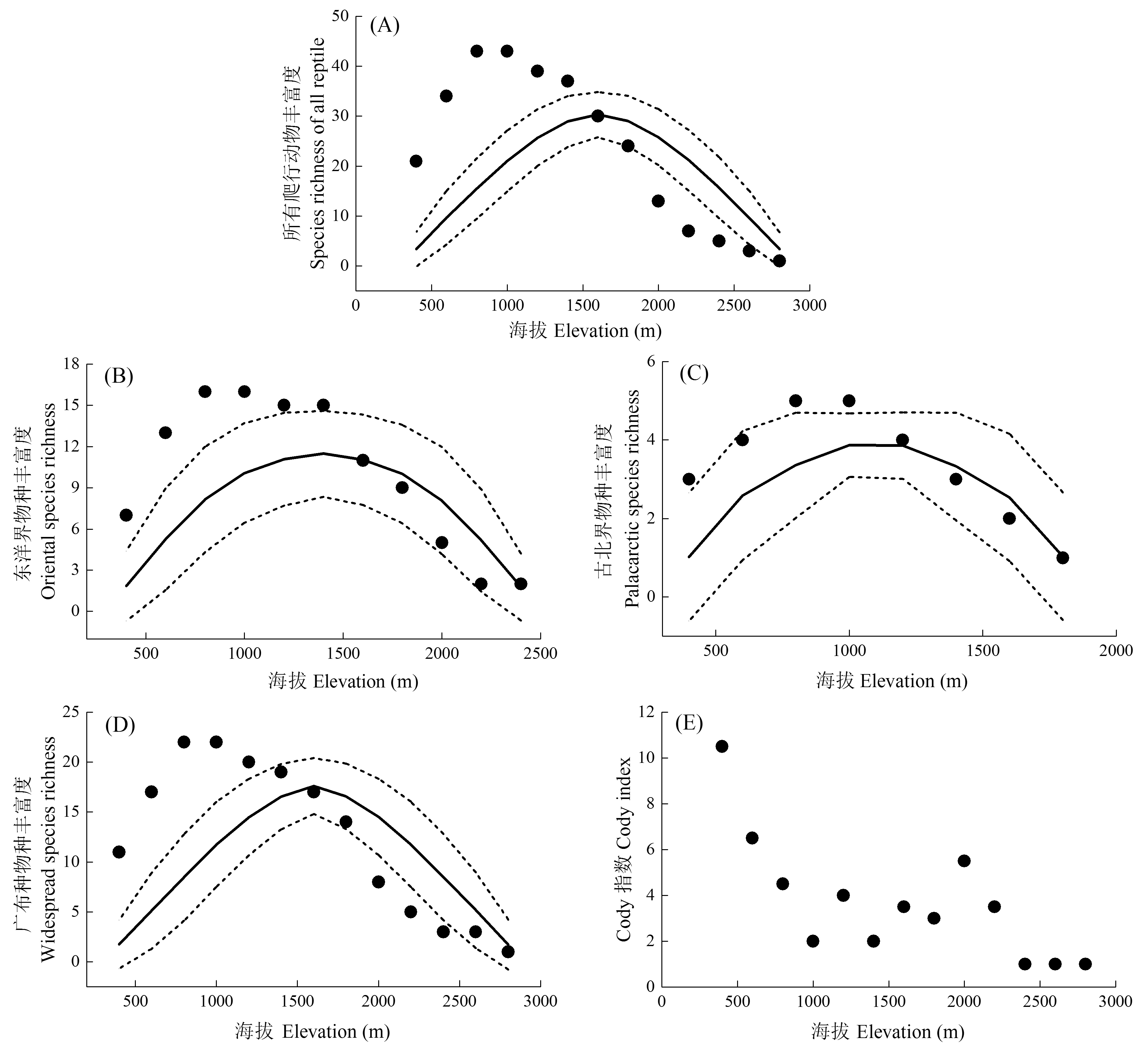
图2 爬行动物及其不同区系成分的物种多样性海拔分布格局。实心点表示实际的物种丰富度, 实线为中域效应模型预测的物种丰富度, 虚线表示模型预测的95%置信区间。
Fig. 2 Elevational distribution patterns of species diversity of reptiles and different faunal components. Dot represents observed richness; solid line represents the predicted richness by mid-domain effect model, while dashed lines represent the 95% confident intervals of predicted richness.
| 类群 Group | 种-面积关系 Species-area relationship (SAR) | 水分-能量假设 Water-energy dynamics (WED) | 中域效应假设 Mid-domain effect (MDE) |
|---|---|---|---|
| 两栖动物 Amphibian | 74.56*** | 94.59*** | 85.76*** |
| 东洋界 Oriental | 83.65*** | 81.94*** | 81.3*** |
| 广布种 Widespread | 47.50** | 87.8*** | 84.65*** |
| 爬行动物 Reptiles | 43.47** | 97.86*** | 21.49 |
| 东洋界 Oriental | 18.34 | 91.44*** | 34.14 |
| 广布种 Widespread | 46.66** | 95.35*** | 23.22 |
| 古北界 Palaearctic | 0.001 | 98.48*** | 41.39 |
表1 不同假设对秦岭两栖、爬行动物物种多样性海拔梯度格局的解释百分率(%)
Table 1 Percentage of variations explained by different hypotheses for the elevational species diversity patterns of amphibians and reptiles in the Qinling Range
| 类群 Group | 种-面积关系 Species-area relationship (SAR) | 水分-能量假设 Water-energy dynamics (WED) | 中域效应假设 Mid-domain effect (MDE) |
|---|---|---|---|
| 两栖动物 Amphibian | 74.56*** | 94.59*** | 85.76*** |
| 东洋界 Oriental | 83.65*** | 81.94*** | 81.3*** |
| 广布种 Widespread | 47.50** | 87.8*** | 84.65*** |
| 爬行动物 Reptiles | 43.47** | 97.86*** | 21.49 |
| 东洋界 Oriental | 18.34 | 91.44*** | 34.14 |
| 广布种 Widespread | 46.66** | 95.35*** | 23.22 |
| 古北界 Palaearctic | 0.001 | 98.48*** | 41.39 |
| 丰富度 Richness | 变量 Variables | AIC | 模型解释力 Model explanation (R2, %) |
|---|---|---|---|
| 两栖动物 Amphibian | Area, PET, PET2, PAN | -98.12 | 99.05 |
| 东洋界 Oriental | Area, PR, PET, PET2, PAN | -72.52 | 96.99 |
| 广布种 Widespread | PR, PAN | -84.55 | 94.08 |
| 爬行动物 Reptile | Area, PR, PET, PET2 | -80.82 | 99.44 |
| 东洋界 Oriental | Area, PR, PET, PET2, PAN | -61.12 | 97.55 |
| 古北界 Palaearctic | Area, PR, PET, PAN | -57.67 | 98.98 |
| 广布种 Widespread | Area, PR, PET, PET2 | -69.34 | 97.98 |
表2 秦岭两栖、爬行动物及其不同区系成分的物种丰富度与不同因子的最优线性模型
Table 2 Linear regression models for the elevational patterns of amphibians, reptiles, their different faunal components and different factors in the Qinling Range.
| 丰富度 Richness | 变量 Variables | AIC | 模型解释力 Model explanation (R2, %) |
|---|---|---|---|
| 两栖动物 Amphibian | Area, PET, PET2, PAN | -98.12 | 99.05 |
| 东洋界 Oriental | Area, PR, PET, PET2, PAN | -72.52 | 96.99 |
| 广布种 Widespread | PR, PAN | -84.55 | 94.08 |
| 爬行动物 Reptile | Area, PR, PET, PET2 | -80.82 | 99.44 |
| 东洋界 Oriental | Area, PR, PET, PET2, PAN | -61.12 | 97.55 |
| 古北界 Palaearctic | Area, PR, PET, PAN | -57.67 | 98.98 |
| 广布种 Widespread | Area, PR, PET, PET2 | -69.34 | 97.98 |
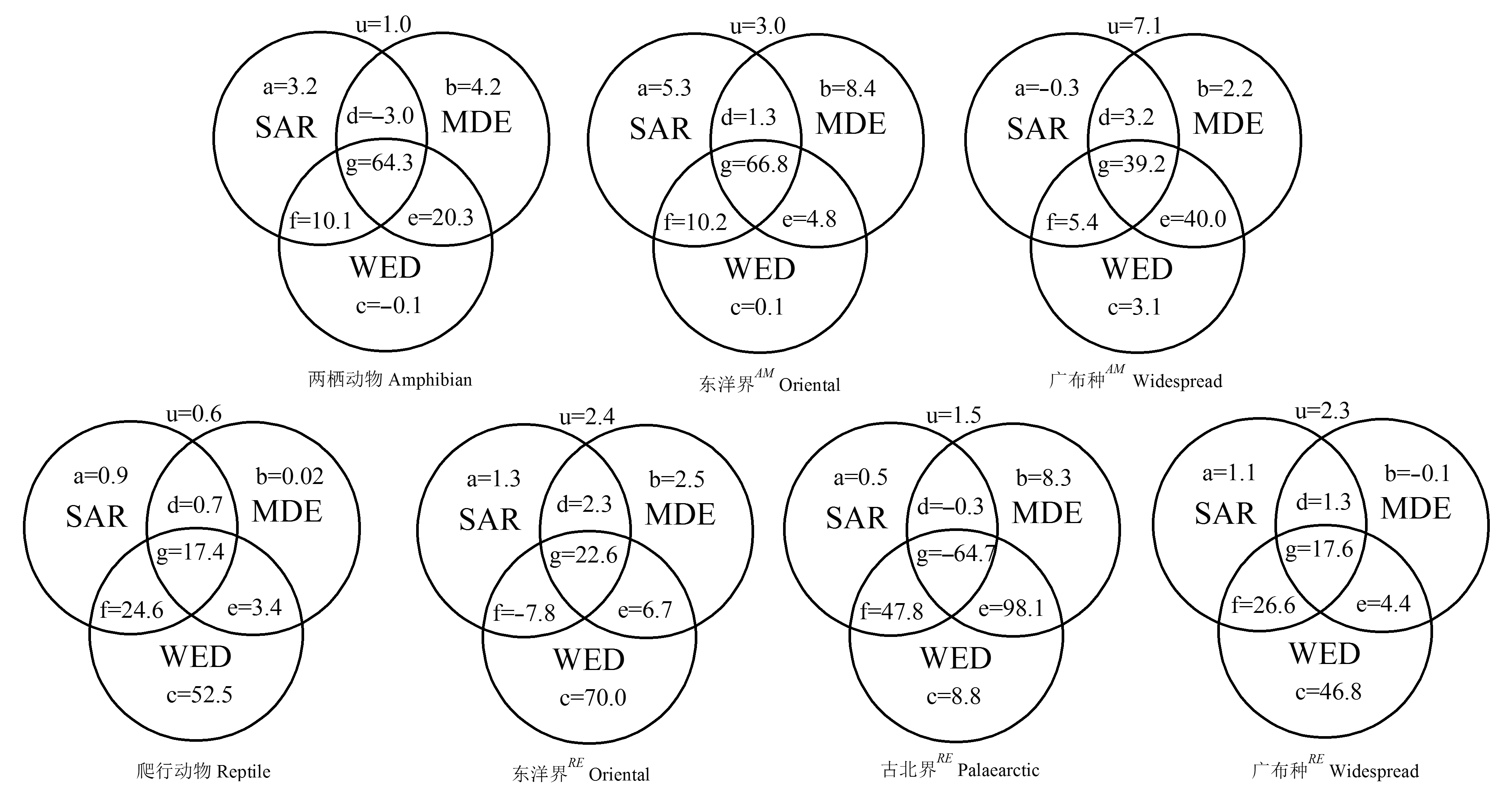
图3 不同环境因子对秦岭两栖、爬行动物及其不同区系成分的物种丰富度海拔梯度格局的影响(以变异解释百分率表示)。a、b、c分别为SAR、MDE、WED的独立作用; d、e、f、g分别表示三种假设之间的交互作用; u表示三种变量未解释量; AM为两栖动物不同区系成分; RE为爬行动物不同区系成分。
Fig. 3 The variation partitioning for the effects of area, water-energy and mid-domain effect on the richness of amphibians, reptiles and their different faunal components in Qinling Range, in terms of the percentage of sum of variances explained. The components a, b and c are the pure effects of area, water-energy, mid-domain effect, while d, e, f and g are the interaction between them. u is an unexplained component of three variables. AM represents different fauna components for amphibians, and RE represents different faunal components for reptiles.
| 物种种域 Species range | 模型 Model | 模型解释力 Model explanation (R2, %) | P |
|---|---|---|---|
| 两栖动物 Amphibian | R = -2.86 PAN + 3,459.22 | 61.64 | 5.33×10-4 |
| R = 15.64 MAT + 1,169.32 | 4.35 | 0.23 | |
| 东洋界 Oriental | R = -3.13 PAN + 3,342.92 | 66.83 | 2.17×10-4 |
| R = 18.61 MAT +823.87 | 7.85 | 0.17 | |
| 广布种 Widespread | R = 15.99 PAN - 8,215.62 | 68.52 | 1.57×10-4 |
| R = 32.74 MAT +3,067.28 | 1.8 | 0.65 | |
| 爬行动物 Reptile | R = -1.88 PAN +2,714.46 | 5.4 | 0.22 |
| R = -67.81 MAT + 2,137.26 | 81.6 | 1.42×10-5 | |
| 东洋界 Oriental | R = 0.25 PAN + 850.63 | 0.6 | 0.82 |
| R= -16.46 MAT +1,249.15 | 10.25 | 0.34 | |
| 古北界 Palaearctic | R = 1.19 PAN + 191.52 | 12.71 | 0.38 |
| R=-62.29 MAT + 1,968.37 | 53.34 | 0.02 | |
| 广布种 Widespread | R = 1.4 PAN + 2,480.54 | 0.47 | 0.33 |
| R = -65.11 MAT + 2,224.88 | 90.57 | 3.47×10-7 |
表3 秦岭两栖、爬行动物及其不同区系成分物种种域大小与环境因子的线性回归模型
Table 3 Linear regression models for the amphibians, reptiles, their different faunal component species range size and different factors in the Qinling Range.
| 物种种域 Species range | 模型 Model | 模型解释力 Model explanation (R2, %) | P |
|---|---|---|---|
| 两栖动物 Amphibian | R = -2.86 PAN + 3,459.22 | 61.64 | 5.33×10-4 |
| R = 15.64 MAT + 1,169.32 | 4.35 | 0.23 | |
| 东洋界 Oriental | R = -3.13 PAN + 3,342.92 | 66.83 | 2.17×10-4 |
| R = 18.61 MAT +823.87 | 7.85 | 0.17 | |
| 广布种 Widespread | R = 15.99 PAN - 8,215.62 | 68.52 | 1.57×10-4 |
| R = 32.74 MAT +3,067.28 | 1.8 | 0.65 | |
| 爬行动物 Reptile | R = -1.88 PAN +2,714.46 | 5.4 | 0.22 |
| R = -67.81 MAT + 2,137.26 | 81.6 | 1.42×10-5 | |
| 东洋界 Oriental | R = 0.25 PAN + 850.63 | 0.6 | 0.82 |
| R= -16.46 MAT +1,249.15 | 10.25 | 0.34 | |
| 古北界 Palaearctic | R = 1.19 PAN + 191.52 | 12.71 | 0.38 |
| R=-62.29 MAT + 1,968.37 | 53.34 | 0.02 | |
| 广布种 Widespread | R = 1.4 PAN + 2,480.54 | 0.47 | 0.33 |
| R = -65.11 MAT + 2,224.88 | 90.57 | 3.47×10-7 |
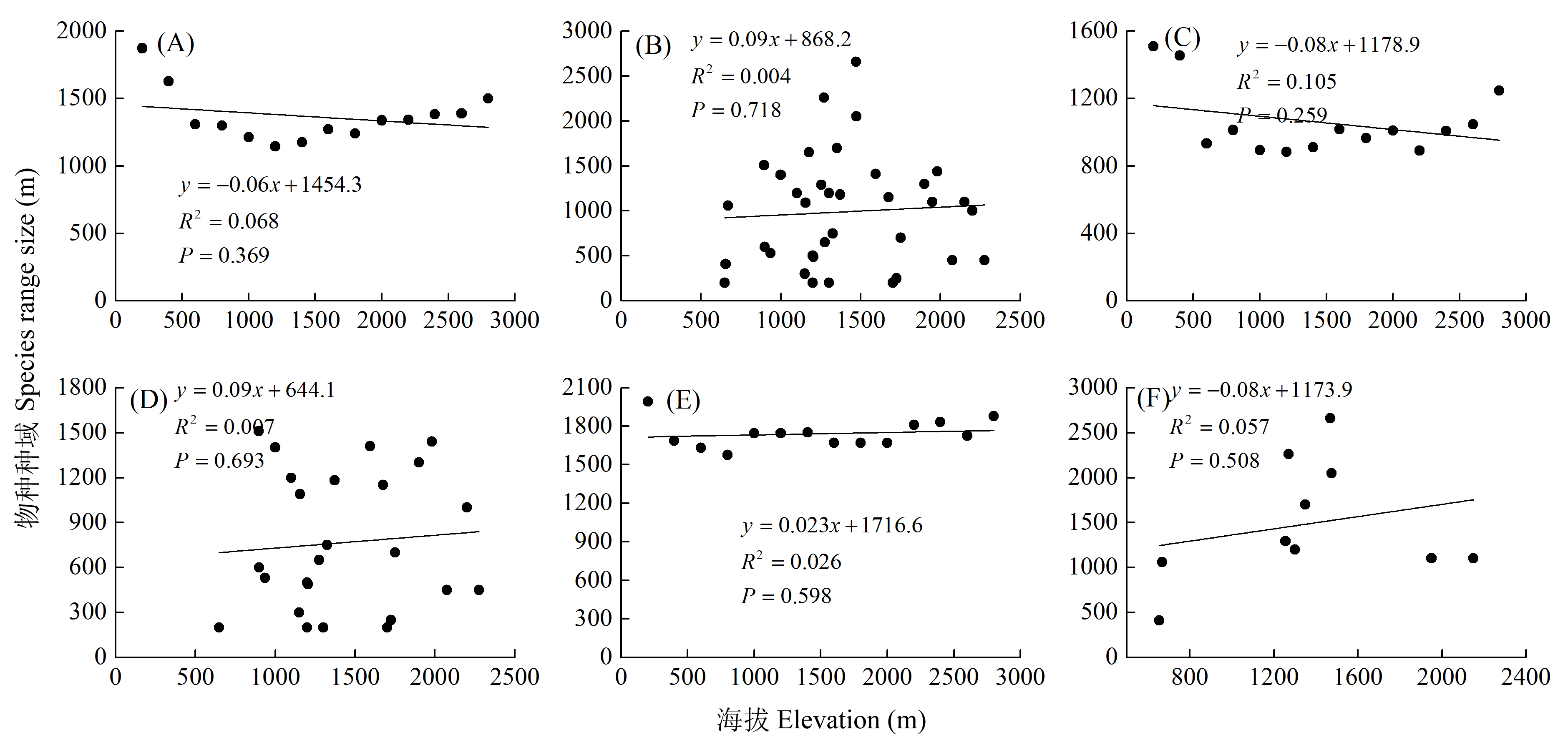
图4 两栖动物种域海拔分布的Rapoport法则验证。(A)所有种Steven方法; (B)所有种逐种法; (C)东洋界物种Steven方法; (D)东洋界物种逐种法; (E)广布种Steven方法; (F)广布种逐种法。
Fig. 4 Test of Rapoport’s rule for amphibian range along the elevational gradient. (A) Steven’s method for amphibians; (B) Cross-species method for amphibian; (C) Steven’s method for oriental; (D) Cross-species method for oriental; (E) Steven’s method for widespread; (F) Cross-species method for widespread
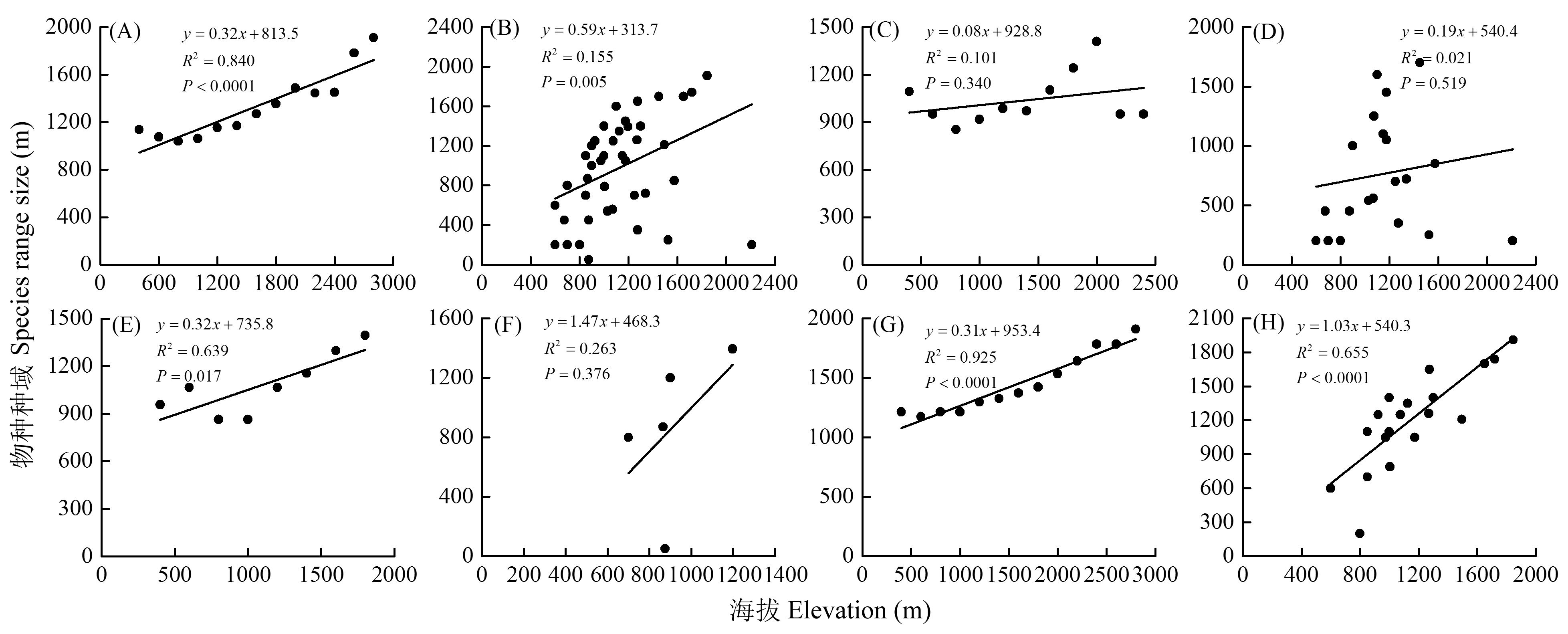
图5 爬行动物种域海拔分布的Rapoport法则验证。(A)所有种Steven’s方法; (B)所有种逐种法; (C)东洋界物种Steven方法; (D)东洋界物种逐种法; (E)古北界物种Steven方法; (F)古北界物种逐种法; (G)广布种Steven方法; (H)广布种逐种法
Fig. 5 Test of Rapoport’s rule for reptile range along the elevational gradients. (A) Steven’s method for reptiles; (B) Cross-species method for reptiles; (C) Steven’s method for oriental; (D) Cross-species method for oriental; (E) Steven’s method for Palaearctic; (F) Cross-species method for Palaearctic (G) Steven’s method for widespread; (H) Cross-species method for widespread
| 物种 Species | 区系成分 Faunal components | 海拔分布 Elevational range (m) | |
|---|---|---|---|
| 两栖动物 | Amphibain | ||
| 隐鳃鲵科 | Cryptobranchidae | ||
| 大鲵* | Andrias davidianus | 广布种 | 700-1,900 |
| 小鲵科 | Hynobiidae | ||
| 秦巴拟小鲵* | Pseudohynobius tsinpaensis | 东洋界 | 1,700-2,700 |
| 黄斑拟小鲵** | P. flavomaculatus | 东洋界 | 1,850-2,300 |
| 西藏山溪鲵* | Batrachuperus tibetanus | 广布种 | 1,260-2,700 |
| 蝾螈科 | Salamandridae | ||
| 文县瑶螈* | Yaotriton wenxianensis | 东洋界 | 1,100-1,300 |
| 角蟾科 | Megophryidae | ||
| 峨山掌突蟾** | Paramegophrys oshanensis | 东洋界 | 950-1,700 |
| 小角蟾** | Megophrys minor | 东洋界 | 1,400-2,100 |
| 南江角蟾** | M. nankiangensis | 东洋界 | 1,600-1,850 |
| 巫山角蟾* | M. wushanensis | 东洋界 | 950-1,450 |
| 川北齿蟾** | Oreolalax chuanbeiensis | 广布种 | 960-1,450 |
| 宁陕齿突蟾** | Scutiger (Scutiger) ningshanensis | 东洋界 | 1,250-2,550 |
| 平武齿突蟾** | S. (Scutiger) pingwuensis | 东洋界 | 2,050-2,500 |
| 胸腺猫眼蟾** | S. (Aelurophryne) glandulatus | 东洋界 | 1,600-1,800 |
| 蟾蜍科 | Bufonidae | ||
| 华西大蟾蜍* | Bufo gargarizans | 东洋界 | 300-1,700 |
| 中华大蟾蜍* | B. gargarizans | 广布种 | 140-2,400 |
| 花背蟾蜍* | Strauchbufo raddei | 古北界 | 350-2,000 |
| 雨蛙科 | Hylidae | ||
| 秦岭雨蛙* | Hyla tsinlingensis | 东洋界 | 890-2,300 |
| 蛙科 | Ranidae | ||
| 黑斑侧褶蛙* | Pelophylax nigromaculatus | 广布种 | 140-2,800 |
| 叉舌蛙科 | Dicroglossidae | ||
| 泽陆蛙* | Fejervarya multistriata | 东洋界 | 140-1,200 |
| 绿臭蛙* | Odorrana margaretae | 东洋界 | 1,000-1,300 |
| 大绿臭蛙* | O. graminea | 东洋界 | 1,200-1,400 |
| 棘腹蛙* | Quasipaa boulengeri | 广布种 | 610-1,900 |
| 太行隆肛蛙* | Feirana taihangnica | 东洋界 | 500-1,700 |
| 康县隆肛蛙* | F. kangxianensis | 东洋界 | 780-1,962 |
| 隆肛蛙* | F. quadranus | 广布种 | 500-2,200 |
| 中国林蛙* | Rana chensinensis | 广布种 | 450-2,500 |
| 崇安湍蛙* | Amolops chunganensis | 东洋界 | 670-1,200 |
| 四川湍蛙* | A. mantzorum | 东洋界 | 1,100-2,250 |
| 树蛙科 | Rhacophoridae | ||
| 斑腿泛树蛙* | Polypedates megacephalus | 东洋界 | 600-1,200 |
| 姬娃科 | Microhylidae | ||
| 合征姬蛙* | Microhyla mixtata | 广布种 | 610-1,700 |
| 饰纹姬娃* | M. fissipes | 东洋界 | 140-1,650 |
| 花姬蛙** | M. pulchra | 东洋界 | 550-750 |
| 北方狭口蛙* | Kaloula borealis | 广布种 | 450-860 |
| 四川狭口蛙** | K. rugifera | 东洋界 | 950-1,600 |
爬行动物 | Reptile | ||
| 龟科 | Testubinidae | ||
| 乌龟* | Chinemys reevesil | 广布种 | 450-1,550 |
| 鳖科 | Trionychidae | ||
| 鳖* | Trionyx sinensis | 广布种 | 450-1,500 |
| 山瑞鳖** | T. steindachneri | 广布种 | 500-600 |
| 石子龙科 | Scincidae | ||
| 蓝尾石龙子* | Eumeces elegans | 广布种 | 700-900 |
| 中国石龙子** | E. chinensis | 东洋界 | 2,110-2,310 |
| 黄纹石龙子* | E. xanthi | 广布种 | 640-1,900 |
| 秦岭滑蜥* | Leiolopisma tsinlingensis | 广布种 | 890-2,100 |
| 铜蜓蜥* | Sphenomorphus indicus | 东洋界 | 450-1,900 |
| 丽斑麻蜥* | Eremias argus | 300-1,100 | |
| 北草蜥* | Takydromus septentrionalis | 450-2,100 | |
| 鬣蜥科 | Agamidae | ||
| 丽纹龙蜥** | Japalura splendida | 东洋界 | 450-900 |
| 草绿龙蜥** | J. flaviceps | 东洋界 | 450-900 |
| 米仓龙蜥* | Japalura micangshanensis | 东洋界 | 760-1,300 |
| 壁虎科 | Gekkonidae | ||
| 多疣壁虎* | Gekko japonicus | 东洋界 | 400-1,400 |
| 无蹼壁虎* | G. swinhonis | 广布种 | 300-1,400 |
| 太白壁虎* | G. taibaiensis | 东洋界 | 900-1,600 |
| 蝰科 | Viperidae | ||
| 白头蝰** | Azemiop feae | 东洋界 | 980-1,700 |
| 短尾蝮* | Gloydius brevicaudus | 广布种 | 850-2,590 |
| 中介蝮* | G. intermedius | 古北界 | 500-1,894 |
| 高原蝮* | G. strauchi | 广布种 | 890-2,800 |
| 山烙铁头* | Ovophis monticola | 东洋界 | 1,150-2,000 |
| 菜花原矛头蝮* | Protobothrops jerdonii | 广布种 | 800-2,500 |
| 竹叶青* | Trimeresurus stejnegeri | 广布种 | 600-2,300 |
| 游蛇科 | Colubridae | ||
| 黑脊蛇* | Achalinus spinalis | 东洋界 | 450-1,900 |
| 绣链腹链蛇** | Amphiesma craspedogaster | 东洋界 | 650-1,700 |
| 棕黑腹链蛇** | A. sauteri | 东洋界 | 650-1,100 |
| 绞花林蛇* | Boiga kraepelini | 东洋界 | 700-900 |
| 黄脊游蛇** | Coluber spinalis | 古北界 | 430-1,300 |
| 翠青蛇* | Cyclophiops major | 广布种 | 450-1,700 |
| 赤链蛇* | Dinodon rufozonatum | 广布种 | 300-1,700 |
| 王锦蛇* | Elaphe carinata | 广布种 | 450-1,800 |
| 白条锦蛇* | E. dione | 古北界 | 300-1,500 |
| 玉斑锦蛇** | E. mandarian | 广布种 | 300-1,550 |
| 紫灰锦蛇** | E. porphyracea | 东洋界 | 600-1,700 |
| 黑眉锦蛇* | E. taeniura | 广布种 | 450-1,700 |
| 棕黑锦蛇** | E. schrenckii | 东洋界 | 300-900 |
| 团花锦蛇** | E. davidi | 古北界 | 850-900 |
| 双斑锦蛇* | E. bimaculate | 广布种 | 500-1,200 |
| 黑背白环蛇** | Lycodon ruhslratilyoo | 东洋界 | 1,100-1,450 |
| 双全白环蛇** | L. fasciatus | 广布种 | 650-1,700 |
| 宁陕小头蛇** | Oligodon ningshanensis | 东洋界 | 1,400-1,650 |
| 横纹小头蛇** | O. multizonatum | 东洋界 | 790-1,350 |
| 福建颈斑蛇** | Plagiopholis styani | 东洋界 | 500-700 |
| 大眼斜鳞蛇* | Pseudoxenodon macrops | 广布种 | 600-2,000 |
| 颈槽蛇* | Rhabdophis nuchalis | 广布种 | 600-2,000 |
| 虎斑颈槽蛇* | R. tigrina | 东洋界 | 300-1,900 |
| 黑头剑蛇** | Sibynophis chinensis | 广布种 | 610-1,400 |
| 乌华游蛇* | Sinonnatrix percarinate | 东洋界 | 450-1,700 |
| 小头坭蛇** | Trachischium tenuiceps | 东洋界 | 600-800 |
| 乌梢蛇* | Zaocys dhumnades | 广布种 | 450-1,800 |
附表1 秦岭两栖、爬行动物物种名录及其海拔分布
Table S1 List of amphibian and reptile species and their elevational range in the Qinling Mountain Ranges http://www.biodiversity-science.net/fileup/PDF/w2014-005-2.pdf
| 物种 Species | 区系成分 Faunal components | 海拔分布 Elevational range (m) | |
|---|---|---|---|
| 两栖动物 | Amphibain | ||
| 隐鳃鲵科 | Cryptobranchidae | ||
| 大鲵* | Andrias davidianus | 广布种 | 700-1,900 |
| 小鲵科 | Hynobiidae | ||
| 秦巴拟小鲵* | Pseudohynobius tsinpaensis | 东洋界 | 1,700-2,700 |
| 黄斑拟小鲵** | P. flavomaculatus | 东洋界 | 1,850-2,300 |
| 西藏山溪鲵* | Batrachuperus tibetanus | 广布种 | 1,260-2,700 |
| 蝾螈科 | Salamandridae | ||
| 文县瑶螈* | Yaotriton wenxianensis | 东洋界 | 1,100-1,300 |
| 角蟾科 | Megophryidae | ||
| 峨山掌突蟾** | Paramegophrys oshanensis | 东洋界 | 950-1,700 |
| 小角蟾** | Megophrys minor | 东洋界 | 1,400-2,100 |
| 南江角蟾** | M. nankiangensis | 东洋界 | 1,600-1,850 |
| 巫山角蟾* | M. wushanensis | 东洋界 | 950-1,450 |
| 川北齿蟾** | Oreolalax chuanbeiensis | 广布种 | 960-1,450 |
| 宁陕齿突蟾** | Scutiger (Scutiger) ningshanensis | 东洋界 | 1,250-2,550 |
| 平武齿突蟾** | S. (Scutiger) pingwuensis | 东洋界 | 2,050-2,500 |
| 胸腺猫眼蟾** | S. (Aelurophryne) glandulatus | 东洋界 | 1,600-1,800 |
| 蟾蜍科 | Bufonidae | ||
| 华西大蟾蜍* | Bufo gargarizans | 东洋界 | 300-1,700 |
| 中华大蟾蜍* | B. gargarizans | 广布种 | 140-2,400 |
| 花背蟾蜍* | Strauchbufo raddei | 古北界 | 350-2,000 |
| 雨蛙科 | Hylidae | ||
| 秦岭雨蛙* | Hyla tsinlingensis | 东洋界 | 890-2,300 |
| 蛙科 | Ranidae | ||
| 黑斑侧褶蛙* | Pelophylax nigromaculatus | 广布种 | 140-2,800 |
| 叉舌蛙科 | Dicroglossidae | ||
| 泽陆蛙* | Fejervarya multistriata | 东洋界 | 140-1,200 |
| 绿臭蛙* | Odorrana margaretae | 东洋界 | 1,000-1,300 |
| 大绿臭蛙* | O. graminea | 东洋界 | 1,200-1,400 |
| 棘腹蛙* | Quasipaa boulengeri | 广布种 | 610-1,900 |
| 太行隆肛蛙* | Feirana taihangnica | 东洋界 | 500-1,700 |
| 康县隆肛蛙* | F. kangxianensis | 东洋界 | 780-1,962 |
| 隆肛蛙* | F. quadranus | 广布种 | 500-2,200 |
| 中国林蛙* | Rana chensinensis | 广布种 | 450-2,500 |
| 崇安湍蛙* | Amolops chunganensis | 东洋界 | 670-1,200 |
| 四川湍蛙* | A. mantzorum | 东洋界 | 1,100-2,250 |
| 树蛙科 | Rhacophoridae | ||
| 斑腿泛树蛙* | Polypedates megacephalus | 东洋界 | 600-1,200 |
| 姬娃科 | Microhylidae | ||
| 合征姬蛙* | Microhyla mixtata | 广布种 | 610-1,700 |
| 饰纹姬娃* | M. fissipes | 东洋界 | 140-1,650 |
| 花姬蛙** | M. pulchra | 东洋界 | 550-750 |
| 北方狭口蛙* | Kaloula borealis | 广布种 | 450-860 |
| 四川狭口蛙** | K. rugifera | 东洋界 | 950-1,600 |
爬行动物 | Reptile | ||
| 龟科 | Testubinidae | ||
| 乌龟* | Chinemys reevesil | 广布种 | 450-1,550 |
| 鳖科 | Trionychidae | ||
| 鳖* | Trionyx sinensis | 广布种 | 450-1,500 |
| 山瑞鳖** | T. steindachneri | 广布种 | 500-600 |
| 石子龙科 | Scincidae | ||
| 蓝尾石龙子* | Eumeces elegans | 广布种 | 700-900 |
| 中国石龙子** | E. chinensis | 东洋界 | 2,110-2,310 |
| 黄纹石龙子* | E. xanthi | 广布种 | 640-1,900 |
| 秦岭滑蜥* | Leiolopisma tsinlingensis | 广布种 | 890-2,100 |
| 铜蜓蜥* | Sphenomorphus indicus | 东洋界 | 450-1,900 |
| 丽斑麻蜥* | Eremias argus | 300-1,100 | |
| 北草蜥* | Takydromus septentrionalis | 450-2,100 | |
| 鬣蜥科 | Agamidae | ||
| 丽纹龙蜥** | Japalura splendida | 东洋界 | 450-900 |
| 草绿龙蜥** | J. flaviceps | 东洋界 | 450-900 |
| 米仓龙蜥* | Japalura micangshanensis | 东洋界 | 760-1,300 |
| 壁虎科 | Gekkonidae | ||
| 多疣壁虎* | Gekko japonicus | 东洋界 | 400-1,400 |
| 无蹼壁虎* | G. swinhonis | 广布种 | 300-1,400 |
| 太白壁虎* | G. taibaiensis | 东洋界 | 900-1,600 |
| 蝰科 | Viperidae | ||
| 白头蝰** | Azemiop feae | 东洋界 | 980-1,700 |
| 短尾蝮* | Gloydius brevicaudus | 广布种 | 850-2,590 |
| 中介蝮* | G. intermedius | 古北界 | 500-1,894 |
| 高原蝮* | G. strauchi | 广布种 | 890-2,800 |
| 山烙铁头* | Ovophis monticola | 东洋界 | 1,150-2,000 |
| 菜花原矛头蝮* | Protobothrops jerdonii | 广布种 | 800-2,500 |
| 竹叶青* | Trimeresurus stejnegeri | 广布种 | 600-2,300 |
| 游蛇科 | Colubridae | ||
| 黑脊蛇* | Achalinus spinalis | 东洋界 | 450-1,900 |
| 绣链腹链蛇** | Amphiesma craspedogaster | 东洋界 | 650-1,700 |
| 棕黑腹链蛇** | A. sauteri | 东洋界 | 650-1,100 |
| 绞花林蛇* | Boiga kraepelini | 东洋界 | 700-900 |
| 黄脊游蛇** | Coluber spinalis | 古北界 | 430-1,300 |
| 翠青蛇* | Cyclophiops major | 广布种 | 450-1,700 |
| 赤链蛇* | Dinodon rufozonatum | 广布种 | 300-1,700 |
| 王锦蛇* | Elaphe carinata | 广布种 | 450-1,800 |
| 白条锦蛇* | E. dione | 古北界 | 300-1,500 |
| 玉斑锦蛇** | E. mandarian | 广布种 | 300-1,550 |
| 紫灰锦蛇** | E. porphyracea | 东洋界 | 600-1,700 |
| 黑眉锦蛇* | E. taeniura | 广布种 | 450-1,700 |
| 棕黑锦蛇** | E. schrenckii | 东洋界 | 300-900 |
| 团花锦蛇** | E. davidi | 古北界 | 850-900 |
| 双斑锦蛇* | E. bimaculate | 广布种 | 500-1,200 |
| 黑背白环蛇** | Lycodon ruhslratilyoo | 东洋界 | 1,100-1,450 |
| 双全白环蛇** | L. fasciatus | 广布种 | 650-1,700 |
| 宁陕小头蛇** | Oligodon ningshanensis | 东洋界 | 1,400-1,650 |
| 横纹小头蛇** | O. multizonatum | 东洋界 | 790-1,350 |
| 福建颈斑蛇** | Plagiopholis styani | 东洋界 | 500-700 |
| 大眼斜鳞蛇* | Pseudoxenodon macrops | 广布种 | 600-2,000 |
| 颈槽蛇* | Rhabdophis nuchalis | 广布种 | 600-2,000 |
| 虎斑颈槽蛇* | R. tigrina | 东洋界 | 300-1,900 |
| 黑头剑蛇** | Sibynophis chinensis | 广布种 | 610-1,400 |
| 乌华游蛇* | Sinonnatrix percarinate | 东洋界 | 450-1,700 |
| 小头坭蛇** | Trachischium tenuiceps | 东洋界 | 600-800 |
| 乌梢蛇* | Zaocys dhumnades | 广布种 | 450-1,800 |
| [1] | .Báldi A (2008) Habitat heterogeneity overrides the species-area relationship. Journal of Biogeography, 35, 675-681. |
| [2] | .Barry RG (2008) Mountain Weather and Climate. Cambridge University Press, Cambridge. |
| [3] | .Beketov MA (2009) The Rapoport effect is detected in a river system and is nested on nested organization. Global Ecology and Biogeography, 18, 498-506. |
| [4] | .Blackburn TM, Gaston KJ (2006) There’s more to macroecology than meets the eye. Global Ecology and Biogeography, 15, 537-540. |
| [5] | .Chi XL (池秀莲), Tang ZY (唐志尧) (2011) Effects of area, temperature and geometric constraints on elevational patterns of species richness: a case study in the Mountain Taibai, Qinling Mountains, China. Chinese Journal of Plant Ecology(植物生态学报), 35, 362-370. (in Chinese with English abstract) |
| [6] | .Colwell RK (2008) RangeModel: tools for exploring and assessing geometric constraints on species richness (the mid-domain effect) along transects. Ecography, 31, 4-7. |
| [7] | .Colwell RK, Rahbek C, Gotelli NJ (2004) The mid-domain effect and species richness patterns: what have we learned so far?The American Naturalist, 163, E1-E23. |
| [8] | .Cooper N, Bielby J, Thomas GH, Purvis A (2008) Macroe- cology and extinction risk correlates of frogs. Global Ecology and Biogeography, 17, 211-221. |
| [9] | .Currie DJ, Kerr JT (2008) Tests of the mid-domain hypothesis: a review of the evidence. Ecological Monographs, 78, 3-18. |
| [10] | .Diniz-Filho JAF, Tôrres NM (2002) Rapoport effect in South American Carnivora (Mammalia): null models under geometric and phylogenetic constraints. Brazilian Journal of Biology, 62, 437-444. |
| [11] | .Editorial Board for Mountains Climate Collected Works (山地气候文集编委会) (1984) Mountain Climate Collected Works (山地气候文集). Meteorological Press, Beijing. (in Chinese) |
| [12] | .Editorial Board of Physical Geography of China, CAS (中国科学院中国自然地理编委会) (1980) The Physical Geography of China, Volume of Physiognomy (中国自然地理·地貌卷). Science Press, Beijing. (in Chinese) |
| [13] | .Fang JY (方精云), Shen ZH (沈泽昊), Tang ZY (唐志尧), Wang ZH (王志恒) (2004) The protocol for the survey plan for plant species diversity of China’s Mountains. Biodiversity Science(生物多样性), 12, 5-9. (in Chinese with English abstract) |
| [14] | .Fei L (费梁), Ye CY (叶昌媛), Jiang JP (江建平) (2012) Colored Atlas of Chinese Amphibians and Their Distributions (中国两栖动物及其分布彩色图鉴). Sichuan Publishing House of Science and Technology, Chengdu. (in Chinese) |
| [15] | .Fraser RH, Currie DJ (1996) The species richness-energy hypothesis in a system where historical factors are thought to prevail: coral reefs. The American Naturalist, 148, 138-159. |
| [16] | .Fu CZ, Hua X, Li J, Chang Z, Pu ZC, Chen JK (2006) Elevational patterns of frog species richness and endemic richness in the Hengduan Mountains, China: geometric constraints, area and climate effects. Ecography, 29, 919-927. |
| [17] | .Fu CZ, Wang JX, Pu ZC, Zhang SL, Chen HL, Zhao B, Chen JL, Wu JH (2007) Elevational gradients of diversity for lizards and snakes in the Hengduan Mountains, China. Biodiversity and Conservation, 16, 707-726. |
| [18] | .Gaston KJ (2003) The Structure and Dynamics of Geographic Ranges. Oxford University Press, New York. |
| [19] | .Gaston KJ (2000) Global patterns in biodiversity. Nature, 405, 220-227. |
| [20] | .Gaston KJ, Chown SL, Evans KL (2008) Ecogeographical rules: elements of a synthesis. Journal of Biogeography, 35, 483-500. |
| [21] | .Gouveia SF, Dobrovolski R, Lemes P, Cassemiro FAS, Diniz-Filho JAF (2013) Environmental steepness, tolerance gradient, and ecogeographical rules in glassfrogs (Anura: Centrolenidae). Biological Journal of the Linnean Society, 108, 773-783. |
| [22] | .Grytnes JA, Beaman JH (2006) Elevational species richness patterns for vascular plants on Mount Kinabalu, Borneo. Journal of Biogeography, 33, 1838-1849. |
| [23] | .Hawkins BA, Diniz-Filho JAF, Weis AE (2005) The mid- domain effect and diversity gradients: is there anything to learn. The American Naturalist, 166, E140-E143. |
| [24] | .Hawkins BA, Field R, Cornell HV, Cornell HV, Currie DJ, Guégan JF, Kaufman DM, Kerr JT, Mittelbach GG, Oberdorff T, O’Brien EM, Porter EE, Turner JRG (2003) Energy, water, and broad-scale geographic patterns of spec- ies richness. Ecology, 84, 3105-3117. |
| [25] | .Heikkinen KR, Luoto M, Kuussaari M, Pöyry J (2005) New insights into butterfly-environment relationships using partitioning methods. Proceeding of the Royal Society B: Biological Sciences, 272, 2203-2210. |
| [26] | .Hu JH, Xie F, Li C, Jiang JP (2011) Elevational patterns of species richness, range and body size for spiny frogs. PLoS ONE, 6, e19817. |
| [27] | .Hunter ML, Yonzon P (1993) Altitudinal distributions of birds, mammals, people, forests, and parks in Nepal. Conservation Biology, 7, 420-423. |
| [28] | .Letcher A, Harvey P (1994) Variation in geographical range size among mammals of the Palearctic. The American Naturalist, 144, 30-42. |
| [29] | .Li C (李成) , Li SQ (李胜全), Wang YZ (王跃招), Zhang T (张涛) (2000) Herpetological survey of southern Gansu Province, China. Sichuan Journal of Zoology(四川动物), 19(2), 74-76. (in Chinese with English abstract) |
| [30] | .Li QY (李巧燕), Wang XP (王襄平) (2013) Elevational pattern of species richness in the Three Gorges Region of the Yangtze River: effect of climate, geometric constraints, area and topographical heterogeneity. Biodiversity Science(生物多样性), 21, 141-152. (in Chinese with English abstract) |
| [31] | .Liang J (梁军), Shen ZH (沈泽昊) (2010) On the test of the Rapoport’s rule, algorithm comparison, and weakening of mid-domain effect: with a case study on the seed plants in Mt. Wuliang, Yunnan Province. Journal of Mountain Science(山地学报), 28, 526-533. (in Chinese with English abstract) |
| [32] | .Lu QY (卢绮妍), Shen ZH (沈泽昊) (2009) Altitudinal pattern of species range size of vascular plants in Mt. Shennongjia: a test of Rapoport’s Rule. Biodiversity Science(生物多样性), 17, 644-651. (in Chinese with English abstract) |
| [33] | .Kreft H, Jetz W (2007) Global patterns and determinants of vascular plant diversity. Proceedings of the National Academy of Sciences, USA, 104, 5925-5930. |
| [34] | .McCain CM (2010) Global analysis of reptile elevatoional diversity. Global Ecology and Biogeography, 19, 541-553. |
| [35] | .McCain CM, Grytnes JA(2010) Elevational gradients in species richness. In: Encyclopedia of Life Sciences (ELS), pp. 1-10. John Wiley & Sons, New York. |
| [36] | .O’Brien EM (1993) Climatic gradients in woody plant species richness: towards an explanation based on an analysis of southern Africa’s woody flora. Journal of Biogeography, 20, 181-198. |
| [37] | .O’Brien EM (1998) Water-energy dynamics, climate, and prediction of woody plant species richness: an interim general model. Journal of Biogeography, 25, 379-398. |
| [38] | .de Oliveira G, Diniz-Filho JAF (2010) Spatial patterns of terrestrial vertebrate richness in Brazilian semiarid, northeastern Brazil: selecting hypotheses and revealing constraints. Journal of Arid Environments, 74, 1418-1426. |
| [39] | .Orme CDL, Davies RG, Burgess M, Eigenbrod F, Pickup N, Olson VA, Webster AJ, Ding TS, Rasmussen PC, Ridgely RS, Stattersfield AJ, Bennett PM, Blackburn TM, Gaston KJ, Owens LPF (2005) Global hotpots of species richness are not congruent with endemism or threat. Nature, 436, 1016-1019. |
| [40] | .Parmesan C (2006) Ecological and evolutionary responses to recent climates change. Annual Review of Ecology, Evolution, and Systematics, 37, 637-669. |
| [41] | .Qiao XJ, Tang ZY, Shen ZH, Fang JY (2012) What causes geographical variation in the species-area relationship? A test from forests in China. Ecography, 35, 1110-1116. |
| [42] | .Rahbek C (1995) The elevational gradient of species richness: a uniform pattern?Ecography, 18, 200-205. |
| [43] | .R Development Core Team (2013) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. . |
| [44] | .Ruggiero A, Werekraut V (2007) One-dimensional analyses of Rapoport’s rule reviewed through meta-analysis. Global Ecology and Biogeography, 16, 401-414. |
| [45] | .Shaanxi Institute of Zoology (陕西省动物研究所), Institute of Hydrobiology of Chinese Academy of Sciences (中国科学院水生生物研究所), Biological Department of Lanzhou University (兰州大学生物系) (1987) The Fishes of Qinling Range (秦岭鱼类志). Science Press, Beijing. (in Chinese) |
| [46] | .Shen ZH (沈泽昊), Lu QY (卢绮妍) (2009) The Rapoport’s rule for the geographic patterns of species range size. Biodiversity Science(生物多样性), 17, 560-567. (in Chinese with English abstract) |
| [47] | .Stevens GC (1989) The latitudinal gradient in geographical range: How so many species coexist in the tropics?The American Naturalist, 133, 240-256. |
| [48] | .Stevens GC (1992) The elevational gradient in altitudinal range, an extension of Rapoport’s latitudinal rule to altitude. The American Naturalist, 140, 893-911. |
| [49] | .Stuart SN, Chanson JS, Cox NA, Young BE, Rodrigues ASL, Fishchman DL, Waller RW (2004) Status and trends of amphibian declines and extinctions worldwide. Science, 306, 1783-1786. |
| [50] | .Tang ZY (唐志尧), Fang JY (方精云) (2004) A review on the elevational patterns of plant species diversity. Biodiversity Science(生物多样性), 12, 20-28. (in Chinese with English abstract) |
| [51] | .Tang ZY (唐志尧), Ke JH (柯金虎) (2004) Altitudinal patterns of plant species diversity in Mt. Niubeiliang, Qinling Mountains. Biodiversity Science(生物多样性), 12, 108-114. (in Chinese with English abstract) |
| [52] | .Triantis KA, Mylonas M, Lika K, Vardinoyannis K (2003) A model for the species-area-habitat relationship. Journal of Biogeography, 30, 19-27. |
| [53] | .Vetaas OR, Grytnes JA (2002) Distribution of vascular plant species richness and endemic richness along the Himalayan elevation gradient in Nepal. Global Ecology and Biogeography, 11, 291-301. |
| [54] | .Vitt LJ, Caldwell JP (2009) Herpetology: An Introductory Biology of Amphibians and Reptiles. Science Press, New York. |
| [55] | .Wang XP, Fang JY (2012) Constraining null models with environmental gradients: a new method for evaluating the effects of environmental factors and geometric constraints on geographic diversity patterns. Ecography, 35, 1147-1159. |
| [56] | .Wang XP (王襄平), Fang JY (方精云), Tang ZY (唐志尧) (2009) The mid-domain effect hypothesis: models, evidence and limitations. Biodiversity Science(生物多样性), 17, 568-578. (in Chinese with English abstract) |
| [57] | .Wang ZH, Tang ZY, Fang JY (2007) Altitudinal patterns of seed plant richness in the Gaoligong Mountains, south-east Tibet, in China’s mountains. Diversity and Distributions, 13, 845-854. |
| [58] | .Whitton FJS, Purvis A, Orme CDL, Olalla-Tárraga MA (2012) Understanding global patterns in amphibian geographic range size: does Rapoport rule?Global Ecology and Biogeography, 21, 179-190. |
| [59] | .Xie F (谢锋), Liu HN (刘惠宁), Stuart SN, Chanson JS, Cox NA, Fischman DL (2006) Review on protective demands of Chinese amphibians. Science in China Series C: Life Sciences(中国科学C辑), 36, 570-581. (in Chinese) |
| [60] | .Zhang MW (张孟闻), Zong Y (宗愉), Ma JF (马积藩) (1998) Fauna Sinica, Reptile, Vol. 1 (中国动物志·爬行纲·第一卷). Science Press, Beijing. (in Chinese) |
| [61] | .Zhang RZ (张荣祖) (2011) Zoogeography of China (中国动物地理). Science Press, Beijing. (in Chinese) |
| [62] | .Zhang WJ (张婉君), Lu QY (卢绮妍), Liang J (梁军), Shen ZH (沈泽昊) (2010) Altitudinal gradients of species richness and range size of vascular plants in Taiwan: a test of Rapoport’s rule. Biodiversity Science(生物多样性), 18, 312-322. (in Chinese with English abstract) |
| [63] | .Zhao EM (赵尔宓) (2006) Snakes of China (中国蛇类). Anhui Science and Technology Publishing House, Hefei. (in Chinese) |
| [64] | .Zhao EM (赵尔宓), Jiang YM (江耀明), Huang QY (黄庆云), Zhao H (赵惠), Zhao KT (赵肯堂), Zhou KY (周开亚), Liu YZ (刘月珍), Liu MY (刘明玉), Li DJ (李德俊), Zhang YX (张玉霞) (1999) Fauna Sinica Reptile Vol. 2 (中国动物志·爬行纲·第二卷). Science Press, Beijing. (in Chinese) |
| [65] | .Zheng Z (郑智), Gong DJ (龚大洁), Sun CX (孙呈祥) (2014) Elevational pattern of species richness and species range size of herpetofauna in Baishuijiang Nature Reserve: a test of Rapoport’s rule. Chinese Journal of Ecology(生态学杂志), 33, 537-546. (in Chinese with English abstract) |
| [66] | .Zhu ZC (朱志诚) (1991) Features of vegetation zones on the Qinling Mountain and its loess plateau. Scientia Geographica Sinica(地理科学), 11, 157-164. (in Chinese) |
| [1] | 顾婧婧, 刘宜卓, 苏杨. 基层地方政府在完成《昆蒙框架》中的作用和难点: 基于《联合国气候变化框架公约》任务的比较[J]. 生物多样性, 2025, 33(3): 24585-. |
| [2] | 贾贞妮, 张意岑, 杜彦君, 任海保. 干扰对中亚热带森林群落物种多样性演替动态的影响[J]. 生物多样性, 2025, 33(2): 24078-. |
| [3] | 魏诗雨, 宋天骄, 罗佳宜, 张燕, 赵子萱, 茹靖雯, 易华, 林雁冰. 秦岭火地塘针叶林土壤细菌群落的海拔分布格局[J]. 生物多样性, 2024, 32(9): 24180-. |
| [4] | 吴琪, 张晓青, 杨雨婷, 周艺博, 马毅, 许大明, 斯幸峰, 王健. 浙江钱江源-百山祖国家公园庆元片区叶附生苔多样性及其时空变化[J]. 生物多样性, 2024, 32(4): 24010-. |
| [5] | 曹可欣, 王敬雯, 郑国, 武鹏峰, 李英滨, 崔淑艳. 降水格局改变及氮沉降对北方典型草原土壤线虫多样性的影响[J]. 生物多样性, 2024, 32(3): 23491-. |
| [6] | 孟敬慈, 王国栋, 曹光兰, 胡楠林, 赵美玲, 赵延彤, 薛振山, 刘波, 朴文华, 姜明. 中国芦苇沼泽植物物种丰富度分布格局及其驱动因素[J]. 生物多样性, 2024, 32(2): 23194-. |
| [7] | 施国杉, 刘峰, 曹光宏, 陈典, 夏尚文, 邓云, 王彬, 杨效东, 林露湘. 西双版纳热带季节雨林木本植物的beta多样性: 空间、环境与林分结构的作用[J]. 生物多样性, 2024, 32(12): 24285-. |
| [8] | 吴芳芳, 刘娜, 何春梅, 原作强, 郝占庆, 尹秋龙. 秦岭山地木本植物群落结构及多样性的海拔梯度格局[J]. 生物多样性, 2024, 32(12): 24239-. |
| [9] | 杜聪聪, 冯学宇, 陈志林. 桥头堡效应中气候生态位差异的缩小促进了红火蚁的入侵[J]. 生物多样性, 2024, 32(11): 24276-. |
| [10] | 原雪姣, 张渊媛, 张衍亮, 胡璐祎, 桑卫国, 杨峥, 陈颀. 基于飞机草历史分布数据拟合的物种分布模型及其预测能力[J]. 生物多样性, 2024, 32(11): 24288-. |
| [11] | 王丽媛, 胡慧建, 姜杰, 胡一鸣. 南岭哺乳类和鸟类物种丰富度空间分布格局及其影响因子[J]. 生物多样性, 2024, 32(1): 23026-. |
| [12] | 刘志发, 王新财, 龚粤宁, 陈道剑, 张强. 基于红外相机监测的广东南岭国家级自然保护区鸟兽多样性及其垂直分布特征[J]. 生物多样性, 2023, 31(8): 22689-. |
| [13] | 冯莉. 国际法视野下生物多样性和气候变化的协同治理[J]. 生物多样性, 2023, 31(7): 23110-. |
| [14] | 陈声文, 任海保, 童光蓉, 王宁宁, 蓝文超, 薛建华, 米湘成. 钱江源国家公园木本植物物种多样性空间分布格局[J]. 生物多样性, 2023, 31(7): 22587-. |
| [15] | 谢艳秋, 黄晖, 王春晓, 何雅琴, 江怡萱, 刘子琳, 邓传远, 郑郁善. 福建海岛滨海特有植物种-面积关系及物种丰富度决定因素[J]. 生物多样性, 2023, 31(5): 22345-. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
备案号:京ICP备16067583号-7
Copyright © 2026 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn
![]()