
生物多样性 ›› 2017, Vol. 25 ›› Issue (6): 608-614. DOI: 10.17520/biods.2017039 cstr: 32101.14.biods.2017039
所属专题: 传粉生物学; 昆虫多样性与生态功能
魏宇昆*( ), 黄艳波, 李桂彬
), 黄艳波, 李桂彬
收稿日期:2017-05-27
接受日期:2017-06-21
出版日期:2017-06-20
发布日期:2017-07-10
通讯作者:
魏宇昆
基金资助:
Yukun Wei*(), Yanbo Huang, Guibin Li
Received:2017-05-27
Accepted:2017-06-21
Online:2017-06-20
Published:2017-07-10
Contact:
Wei Yukun
摘要:
生殖隔离是生物多样性产生的重要原因之一, 不同物种间的生殖隔离形成阶段、方式和强度不完全相同。为了揭示生殖隔离在鼠尾草属(Salvia)物种多样性产生和维持过程中的作用和特点, 本文研究了浙江天目山同域分布的舌瓣鼠尾草(S. liguliloba)和南丹参(S. bowleyana)的开花、传粉和生殖隔离, 利用人工授粉和杂交实验检测它们的种间遗传相容程度。结果表明, 在花的形态结构、花及花序的生长、着生方式与数量特征上, 2种鼠尾草属植物均差异显著(P < 0.05)。2种鼠尾草属植物共享同一种有效传粉昆虫——三条熊蜂(Bombus trifasciatus), 但通过访花行为的显著差异和明显不同的花粉落置部位, 有效避免了异源花粉干扰。人工杂交授粉的结果表明2个物种间遗传相容性较高, 互为亲本的杂交结实率分别达到77.8 ± 10.7%和78.7 ± 11.2%。我们的研究表明, 同域分布、花期重叠且共享唯一传粉者的2种鼠尾草属植物, 以花部的传粉结构和传粉部位的差异, 产生了较高程度的传粉前生殖隔离, 避免了物种间的异源花粉干扰和自然杂交, 保障各自较高的繁殖成效, 以维持自然种群的多样性和遗传结构的稳定。
魏宇昆, 黄艳波, 李桂彬 (2017) 同域分布共享传粉者的鼠尾草属植物的生殖隔离. 生物多样性, 25, 608-614. DOI: 10.17520/biods.2017039.
Yukun Wei, Yanbo Huang, Guibin Li (2017) Reproductive isolation in sympatric Salvia species sharing a sole pollinator. Biodiversity Science, 25, 608-614. DOI: 10.17520/biods.2017039.

图1 舌瓣鼠尾草和南丹参的花序、花结构及昆虫传粉特征(图中标尺均为2 mm)。(a)舌瓣鼠尾草花序; (b)舌瓣鼠尾草花冠纵切(箭头所示为雄蕊花药); (c)舌瓣鼠尾草花正面观; (d)三条熊蜂访问舌瓣鼠尾草及其传粉过程(箭头所示为传粉部位); (e)南丹参花序; (f)南丹参花冠纵切(箭头所示为雄蕊花药); (g)南丹参花正面观; (h)三条熊蜂访问南丹参及其传粉过程(箭头所示为传粉部位)。
Fig. 1 Comparison of inflorescences, flower structures, and insect pollinations of Salvia liguliloba and S. bowleyana (Bar = 2 mm). (a) Inflorescences of S. liguliloba; (b) Flower lateral dissection of S. liguliloba (the arrow indicates anther); (c) A front view of S. liguliloba flower; (d) Bombus trifasciatus is visiting flowers and pollinating S. liguliloba (the arrow indicates pollen placement sites on bumblebee); (e) Inflorescences of S. bowleyana; (f) Flower lateral dissection of S. bowleyana (the arrow indicates anther); (g) A front view of S. bowleyana flower; (h) Bombus trifasciatus is visiting flowers and pollinating S. bowleyana (the arrow indicates pollen placement sites on bumblebee).
| 花冠长 Corolla length | 花冠宽 Corolla width | 花冠高 Corolla height | 冠筒长 Tube length | 冠口高 Entrance height | 冠筒口高 Tube entrance height | 冠筒口宽 Tube entrance width | 花丝长 Filament length | 药隔长 Connective length | 雌蕊长 Pistil length | |
|---|---|---|---|---|---|---|---|---|---|---|
| 舌瓣鼠尾草 S. liguliloba | 23.7 ± 0.74a | 5.31 ± 0.34a | 7.30 ± 0.67b | 19.5 ± 0.8a | 4.87 ± 0.61b | 4.87 ± 0.61a | 4.41 ± 0.52a | 2.39 ± 0.14b | 5.19 ± 0.33b | 23.7 ± 0.43b |
| 南丹参 S. bowleyana | 22.8 ± 1.10b | 4.47 ± 0.26b | 16.6 ± 0.94a | 12.4 ± 0.76b | 11.2 ± 0.80a | 4.47 ± 0.28a | 3.21 ± 0.23b | 4.30 ± 0.26a | 14.3 ± 0.68a | 26.1 ± 1.85a |
| P | 0.0407 | 0.000 | 0.000 | 0.000 | 0.000 | 0.0585 | 0.000 | 0.000 | 0.000 | 0.0011 |
表1 舌瓣鼠尾草与南丹参的花部结构比较(平均值 ± 标准差)
Table 1 Comparison of flower structures between Salvia liguliloba and S. bowleyana (mean ± SD)
| 花冠长 Corolla length | 花冠宽 Corolla width | 花冠高 Corolla height | 冠筒长 Tube length | 冠口高 Entrance height | 冠筒口高 Tube entrance height | 冠筒口宽 Tube entrance width | 花丝长 Filament length | 药隔长 Connective length | 雌蕊长 Pistil length | |
|---|---|---|---|---|---|---|---|---|---|---|
| 舌瓣鼠尾草 S. liguliloba | 23.7 ± 0.74a | 5.31 ± 0.34a | 7.30 ± 0.67b | 19.5 ± 0.8a | 4.87 ± 0.61b | 4.87 ± 0.61a | 4.41 ± 0.52a | 2.39 ± 0.14b | 5.19 ± 0.33b | 23.7 ± 0.43b |
| 南丹参 S. bowleyana | 22.8 ± 1.10b | 4.47 ± 0.26b | 16.6 ± 0.94a | 12.4 ± 0.76b | 11.2 ± 0.80a | 4.47 ± 0.28a | 3.21 ± 0.23b | 4.30 ± 0.26a | 14.3 ± 0.68a | 26.1 ± 1.85a |
| P | 0.0407 | 0.000 | 0.000 | 0.000 | 0.000 | 0.0585 | 0.000 | 0.000 | 0.000 | 0.0011 |
| 单株花序总数 No. of infloresce- nce in individual plant | 假圆锥花序 分枝数 No. of branch in a false panicle | 假总状花序的 轮伞花序数 No. of verticillaster in a false raceme | 轮伞花序小 花总数 No. of flower in a verticillaster | 轮伞花序同时 开放小花数 No. of flowering in a verticillaster | 单株总花数 Total no. of flower in individual plant | 单株总开花数 Total no. of flowering in individual plant | |
|---|---|---|---|---|---|---|---|
| 舌瓣鼠尾草 S. liguliloba | 2.0±2.1a (n = 35) | 2.0±1.9b (n = 39) | 9.3±2.4a (n = 83) | 5.6±1.1b (n = 93) | 1.4±1.7b (n = 234) | 262±299b (n = 26) | 21±30a (n = 36) |
| 南丹参 S. bowleyana | 2.2±0.45a (n = 5) | 5.8±3.0a (n = 11) | 9.5±1.9a (n = 40) | 9.8±1.9a (n = 160) | 1.8±1.0a (n = 160) | 891±472a (n = 5) | 48±56a (n = 16) |
| P | 0.681 | 0.0020 | 0.7166 | 0.000 | 0.0053 | 0.0005 | 0.089 |
表2 舌瓣鼠尾草和南丹参花及花序数量特征(平均值 ± 标准差)
Table 2 Comparisons of flowering and inflorescence between Salvia liguliloba and S. bowleyana (mean ± SD)
| 单株花序总数 No. of infloresce- nce in individual plant | 假圆锥花序 分枝数 No. of branch in a false panicle | 假总状花序的 轮伞花序数 No. of verticillaster in a false raceme | 轮伞花序小 花总数 No. of flower in a verticillaster | 轮伞花序同时 开放小花数 No. of flowering in a verticillaster | 单株总花数 Total no. of flower in individual plant | 单株总开花数 Total no. of flowering in individual plant | |
|---|---|---|---|---|---|---|---|
| 舌瓣鼠尾草 S. liguliloba | 2.0±2.1a (n = 35) | 2.0±1.9b (n = 39) | 9.3±2.4a (n = 83) | 5.6±1.1b (n = 93) | 1.4±1.7b (n = 234) | 262±299b (n = 26) | 21±30a (n = 36) |
| 南丹参 S. bowleyana | 2.2±0.45a (n = 5) | 5.8±3.0a (n = 11) | 9.5±1.9a (n = 40) | 9.8±1.9a (n = 160) | 1.8±1.0a (n = 160) | 891±472a (n = 5) | 48±56a (n = 16) |
| P | 0.681 | 0.0020 | 0.7166 | 0.000 | 0.0053 | 0.0005 | 0.089 |
| 相对频次 Relative frequency | 单花访问时间 Visit time per flower (s) | 活跃度 Activity rate | 访花频率 Visitation rate | |
|---|---|---|---|---|
| 舌瓣鼠尾草 S. liguliloba | 0.14 ± 0.017 b (n = 3) | 2.01 ± 0.85 a (n = 30) | 9.71 ± 3.15 b (n = 7) | 1.36 ± 0.17 b (n = 3) |
| 南丹参 S. bowleyana | 0.86 ± 0.017 a (n = 3) | 1.80 ± 0.63 a (n = 30) | 16.2 ± 3.4 a (n = 52) | 13.9 ± 0.28 a (n = 3) |
| P | 0.000 | 0.2758 | 0.000 | 0.000 |
表3 三条熊蜂对2种鼠尾草的访花行为比较(平均值 ± 标准差)
Table 3 Comparisons of flower visiting behavior of Bombus trifasciatus between Salvia liguliloba and S. bowleyana (mean ± SD)
| 相对频次 Relative frequency | 单花访问时间 Visit time per flower (s) | 活跃度 Activity rate | 访花频率 Visitation rate | |
|---|---|---|---|---|
| 舌瓣鼠尾草 S. liguliloba | 0.14 ± 0.017 b (n = 3) | 2.01 ± 0.85 a (n = 30) | 9.71 ± 3.15 b (n = 7) | 1.36 ± 0.17 b (n = 3) |
| 南丹参 S. bowleyana | 0.86 ± 0.017 a (n = 3) | 1.80 ± 0.63 a (n = 30) | 16.2 ± 3.4 a (n = 52) | 13.9 ± 0.28 a (n = 3) |
| P | 0.000 | 0.2758 | 0.000 | 0.000 |
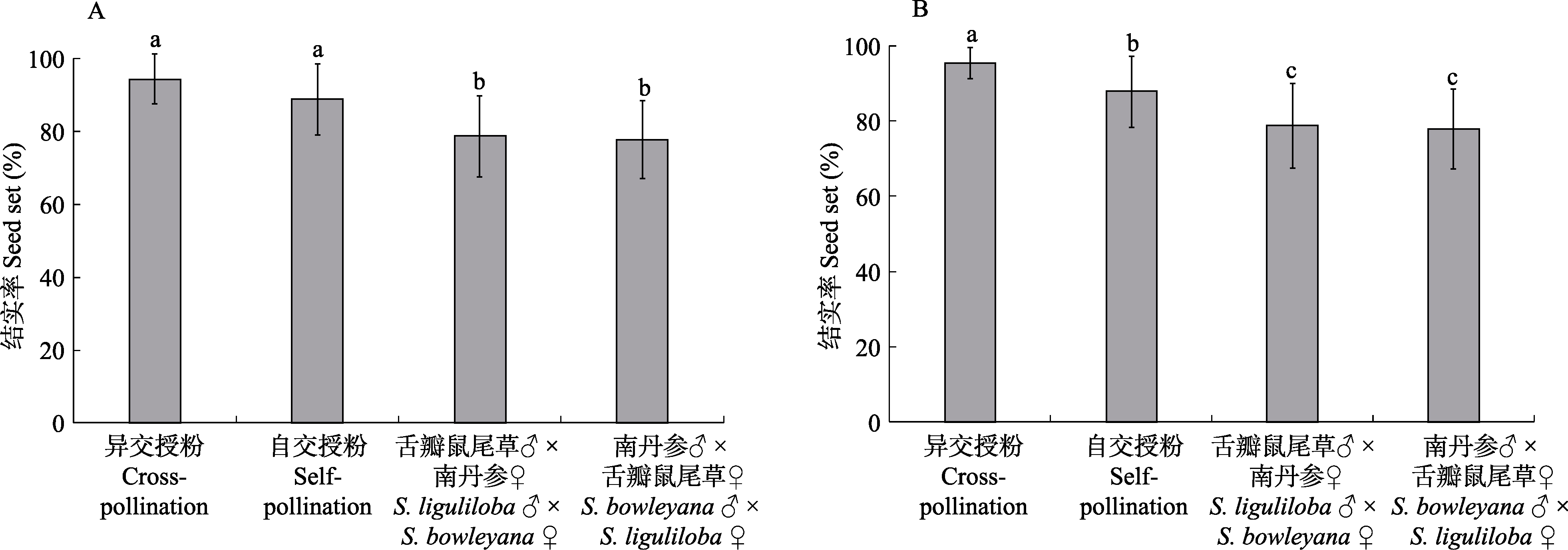
图2 2种鼠尾草的种内异交、自交和种间杂交结实率比较(平均值 ± 标准差)。(A)舌瓣鼠尾草异交、自交以及与南丹参杂交的结实率多重比较结果; (B)南丹参异交、自交以及与舌瓣鼠尾草杂交的结实率多重比较结果。统计分析采用t-检验, 不同字母代表差异显著(P < 0.05)。
Fig. 2 Comparisons of cross-pollination, self-pollination, and hybridization of Salvia liguliloba and S. bowleyana. (A) and (B) show S. liguliloba and S. bowleyana, respectively. Different letters indicate significant difference at the level of P < 0.05 by t-test.
| [1] | Baack E, Melo MC, Rieseberg LH, Ortiz-Barrientos D (2015) The origins of reproductive isolation in plants. New Phytologist, 207, 968-984. |
| [2] | Claßen-Bockhoff R, Speck T, Tweraser E, Wester P, Thimm S, Reith M (2004) The staminal lever mechanism in Salvia L. (Lamiaceae): a key innovation for adaptive radiation? Organisms Diversity & Evolution, 4, 189-205. |
| [3] | Claßen-Bockhoff R, Wester P, Tweraser E (2003) The staminal lever mechanism in Salvia L. (Lamiaceae): a review. Plant Biology, 5, 33-41. |
| [4] | Epling C (1947) Natural hybridization of Salvia apiana and S. mellifera. Evolution, 1, 69-78. |
| [5] | Hopkins R (2013) Reinforcement in plants. New Phytologist, 197, 1095-1103. |
| [6] | Huang S, Shi X (2013) Floral isolation in Pedicularis: how do congeners with shared pollinators minimize reproductive interference? New Phytologist, 199, 858-865. |
| [7] | Huang YB, Wei YK, Ge BJ, Wang Q (2014) Research progress in pollination biology of genus Salvia (Lamiaceae) and their pollination mechanisms in East Asia (China). Acta Ecologica Sinica, 34, 2282-2289. (in Chinese with English abstract) |
| [黄艳波, 魏宇昆, 葛斌杰, 王琦 (2014) 鼠尾草属东亚分支的传粉模式. 生态学报, 34, 2282-2289.] | |
| [8] | Huang YB, Wei YK, Wang Q, Xiao YE, Ye XY (2015) Floral morphology and pollination mechanism of Salvia liguliloba, a narrow endemic species with degraded lever-like stamens. Chinese Journal of Plant Ecology, 39, 753-761. (in Chinese with English abstract) |
| [黄艳波, 魏宇昆, 王琦, 肖月娥, 叶喜阳 (2015) 舌瓣鼠尾草退化杠杆雄蕊的相关花部特征及传粉机制. 植物生态学报, 39, 753-761.] | |
| [9] | Huang Z, Liu H, Huang S (2015) Interspecific pollen transfer between two coflowering species was minimized by bumblebee fidelity and differential pollen placement on the bumblebee body. Journal of Plant Ecology, 8, 109-115. |
| [10] | Kipling R, Warren J (2013) How generalists coexist: the role of floral phenotype and spatial factors in the pollination systems of two Ranunculus species. Journal of Plant Ecology, 7, 480-489. |
| [11] | Ma YP, Xie WJ, Sun WB, Marczewski T (2016a) Strong reproductive isolation despite occasional hybridization between a widely distributed and a narrow endemic Rhododendron species. Scientific Reports, 6, 19146. |
| [12] | Ma YP, Zhou RC, Milne R (2016b) Pollinator-mediated isolation may be an underestimated factor in promoting homoploid hybrid speciation. Frontiers in Plant Science, 7, 1183. |
| [13] | Melo MC, Grealy A, Brittain B, Walter GM, Ortiz-Barrientos D (2014) Strong extrinsic reproductive isolation between parapatric populations of an Australian groundsel. New Phytologist, 203, 323-334. |
| [14] | Meyn O, Emboden WA (1987) Parameters and consequences of introgression in Salvia apiana × S. mellifera (Lamiaceae). Systematic Botany, 12, 390-399. |
| [15] | Pauw A (2013) Can pollination niches facilitate plant coexistence? Trends in Ecology & Evolution, 28, 30-37. |
| [16] | Pedron M, Buzatto CR, Singer RB, Batista JAN, Moser A (2012) Pollination biology of four sympatric species of Habenaria (Orchidaceae: Orchidinae) from southern Brazil. Botanical Journal of the Linnean Society, 170, 141-156. |
| [17] | Ruchisansakun S, Tangtorwongsakul P, Cozien RJ, Smets EF, Niet TVD (2016) Floral specialization for different pollinators and divergent use of the same pollinator among co-occuring Impatiens species (Balsaminaceae) from Southeast Asia. Botanical Journal of the Linnean Society, 181, 651-666. |
| [18] | Wei YK, Wang Q, Huang YB (2015) Species diversity and distribution of Salvia (Lamiaceae). Biodiversity Science, 23, 3-10. (in Chinese with English abstract) |
| [魏宇昆, 王琦, 黄艳波 (2015) 唇形科鼠尾草属的物种多样性与分布. 生物多样性, 23, 3-10.] | |
| [19] | Wester P, Claßen-Bockhoff R (2007) Floral diversity and pollen transfer mechanisms in bird-pollinated Salvia species. Annals of Botany, 100, 401-421. |
| [20] | Widmer A, Lexer C, Cozzolino S (2009) Evolution of reproductive isolation in plants. Heredity, 102, 31-38. |
| [21] | Yang FC (1992) Comprehensive Investigation Report on Natural Resource of Tianmu Mountain Nature Reserve. Zhejiang Science and Technology Press, Hangzhou. (in Chinese) |
| [杨逢春 (1992) 天目山自然保护区自然资源综合考察报告. 浙江科学技术出版社, 杭州.] |
| [1] | 吴晓晴 张美惠 葛苏婷 李漫淑 宋坤 沈国春 达良俊 张健. 上海近自然林重建过程中木本植物物种多样性与地上生物量的时空动态——以闵行区生态岛为例[J]. 生物多样性, 2025, 33(5): 24444-. |
| [2] | 王太, 宋福俊, 张永胜, 娄忠玉, 张艳萍, 杜岩岩. 河西走廊内陆河水系鱼类多样性及资源现状[J]. 生物多样性, 2025, 33(4): 24387-. |
| [3] | 张晶晶, 黄文彬, 陈奕廷, 杨泽鹏, 柯伟业, 彭昭杰, 魏世超, 张志伟, 胡怡思, 余文华, 周文良. 广东南澎列岛海洋生态国家级自然保护区造礁石珊瑚多样性及分布特征[J]. 生物多样性, 2025, 33(4): 24424-. |
| [4] | 尚华丹, 张楚晴, 王梅, 裴文娅, 李国宏, 王鸿斌. 中国杨树害虫物种多样性及其地理分布[J]. 生物多样性, 2025, 33(2): 24370-. |
| [5] | 吴昱萱, 王平, 胡晓生, 丁一, 彭甜恬, 植秋滢, 巴德木其其格, 李文杰, 关潇, 李俊生. 呼伦贝尔草地退化现状评估与植被特征变化[J]. 生物多样性, 2025, 33(2): 24118-. |
| [6] | 陈自宏, 张翼飞, 陈凯, 陈见影, 徐玲. 高黎贡山南段昆虫病原真菌物种多样性及影响因素[J]. 生物多样性, 2025, 33(1): 24228-. |
| [7] | 谭珂, 宁瑶, 王仁芬, 王晴, 梁丹萍, 辛子兵, 温放. 中国苦苣苔科植物名录与地理分布数据集[J]. 生物多样性, 2025, 33(1): 23275-. |
| [8] | 韩佳楠, 苏杨, 李霏, 刘君妍, 赵依林, 李琳, 赵建成, 梁红柱, 李敏. 河北省苔藓植物多样性[J]. 生物多样性, 2024, 32(9): 24096-. |
| [9] | 李东红, 郝媛媛, 甘辉林, 张航, 刘耀猛, 他富源, 胡桂馨. 祁连山北麓中段不同类型草地蝗虫种类及分布[J]. 生物多样性, 2024, 32(9): 24119-. |
| [10] | 牛红玉, 陈璐, 赵恒月, 古丽扎尔·阿不都克力木, 张洪茂. 城市化对动物的影响: 从群落到个体[J]. 生物多样性, 2024, 32(8): 23489-. |
| [11] | 白雪, 李正飞, 刘洋, 张君倩, 张多鹏, 罗鑫, 杨佳莉, 杜丽娜, 蒋玄空, 武瑞文, 谢志才. 西江流域大型底栖无脊椎动物物种多样性及维持机制[J]. 生物多样性, 2024, 32(7): 23499-. |
| [12] | 许佳, 崔小娟, 张翼飞, 吴昌, 孙远东. 南岭地区鱼类多样性及其地理分布[J]. 生物多样性, 2024, 32(7): 23482-. |
| [13] | 邝起宇, 胡亮. 广东东海岛与硇洲岛海域底栖贝类物种多样性及其地理分布[J]. 生物多样性, 2024, 32(5): 24065-. |
| [14] | 赵勇强, 阎玺羽, 谢加琪, 侯梦婷, 陈丹梅, 臧丽鹏, 刘庆福, 隋明浈, 张广奇. 退化喀斯特森林自然恢复中不同生活史阶段木本植物物种多样性与群落构建[J]. 生物多样性, 2024, 32(5): 23462-. |
| [15] | 徐伟强, 苏强. 分形模型与一般性物种多度分布关系的检验解析:以贝类和昆虫群落为例[J]. 生物多样性, 2024, 32(4): 23410-. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
备案号:京ICP备16067583号-7
Copyright © 2026 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn
![]()