
生物多样性 ›› 2021, Vol. 29 ›› Issue (4): 419-427. DOI: 10.17520/biods.2020387 cstr: 32101.14.biods.2020387
所属专题: 生物入侵
• 研究报告: 植物多样性 • 下一篇
黄河燕1,2, 朱政财4, 吴纪华2,3,1, 拉琼2,3, 周永洪2,3, 潘晓云2,3,1,*( )
)
收稿日期:2020-10-08
接受日期:2021-01-09
出版日期:2021-04-20
发布日期:2021-04-20
通讯作者:
潘晓云
基金资助:
Heyan Huang1,2, Zhengcai Zhu4, Jihua Wu2,3,1, Qiong La2,3, Yonghong Zhou2,3, Xiaoyun Pan2,3,1,*()
Received:2020-10-08
Accepted:2021-01-09
Online:2021-04-20
Published:2021-04-20
Contact:
Xiaoyun Pan
About author:* E-mail: xypan@fudan.edu.cn摘要:
植物可以通过关键功能性状的表型可塑性来适应气候变暖背景下环境温度的增加。表型可塑性增强进化假说(evolution of increased phenotypic plasticity hypothesis)认为外来植物在引入地进化出了更强的表型可塑性。以往对该假说的验证多集中于外来植物对光照、水分、养分、邻体以及天敌等的可塑性进化, 而对增温条件下植物生长和功能性状可塑性进化的研究相对较少。仅有的几项研究多集中在温带地区, 且多集中于研究植物生长相关的性状, 而对植物的抗性和草食作用对增温的响应的关注相对较少。本研究采用同质园实验比较了喜旱莲子草(Alternanthera philoxeroides)引入地(中国)和原产地(阿根廷)各8个种群的生物量、功能性状和草食作用在热带地区(广州市增城区)对模拟全天增温2℃的响应差异。结果表明: (1)模拟全天增温显著降低了喜旱莲子草总生物量(-7.8%)、贮藏根生物量(-12.8%)、分枝强度(-11.6%)和茎端取食率(-34.4%)。(2)模拟全天增温造成的引入地种群总生物量降低幅度大于原产地种群; 模拟全天增温使引入地种群的比茎长和茎端取食率降低, 而原产地种群则相反。(3)无论是否模拟全天增温, 引入地种群的贮藏根生物量(+31.5%)、分枝强度(+38.5%)、比茎长(+30.2%)、根冠比(+24.5%)和比叶面积(+20.0%)均高于原产地种群, 茎端取食率则低于原产地种群(-35.8%)。这些结果表明, 热带地区全天增温2℃对喜旱莲子草是一种胁迫; 引入地种群的生物量对模拟全天增温2℃的响应更强, 而其株形相关性状(比茎长)和草食作用(茎端取食率)对模拟全天增温的可塑性方向与原产地种群相反。由于引入地种群在热带地区模拟全天增温条件下生物量的下降和草食作用的增加明显高于原产地种群, 因此在未来全球气候变暖的背景下, 热带地区温度升高可能不利于喜旱莲子草种群多度的增加。
黄河燕, 朱政财, 吴纪华, 拉琼, 周永洪, 潘晓云 (2021) 喜旱莲子草对模拟全天增温的可塑性: 引入地和原产地种群的比较. 生物多样性, 29, 419-427. DOI: 10.17520/biods.2020387.
Heyan Huang, Zhengcai Zhu, Jihua Wu, Qiong La, Yonghong Zhou, Xiaoyun Pan (2021) Phenotypic plasticity of Alternanthera philoxeroides in response to simulated daily warming: Introduced vs. native populations. Biodiversity Science, 29, 419-427. DOI: 10.17520/biods.2020387.
| 变异来源Source of variation | 总生物量 Total biomass | 贮藏根生物量 STB | 分枝强度 BI | 比茎长 SSL | 根冠比 RSR | 比叶面积 SLA | 相对取食面积 RFA | 茎端取食率 SFP | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| χ2 | P | χ2 | P | χ2 | P | χ2 | P | χ2 | P | χ2 | P | χ2 | P | χ2 | P | |
| 增温 Warming (W) | 5.165 | 0.023 | 3.207 | 0.073 | 4.239 | 0.040 | 0.761 | 0.383 | 0.343 | 0.558 | 0.021 | 0.884 | 0.083 | 0.773 | 10.552 | 0.001 |
| 来源地 Origin (O) | 1.523 | 0.217 | 8.756 | 0.003 | 8.023 | 0.005 | 8.092 | 0.004 | 5.758 | 0.016 | 4.665 | 0.031 | 1.837 | 0.175 | 3.680 | 0.055 |
| W × O | 2.986 | 0.084 | 0.244 | 0.621 | 1.077 | 0.299 | 3.453 | 0.063 | 0.913 | 0.339 | 0.365 | 0.546 | 3.682 | 0.055 | 1.772 | 0.183 |
表1 喜旱莲子草引入地(中国)和原产地(阿根廷)种群在两种处理(不增温、全天增温2℃)下生物量(总生物量、贮藏根生物量和生长速率)、功能性状(分枝强度、比茎长、根冠比和比叶面积)和草食作用(相对取食面积和茎端取食率)的差异。字体加粗表示效应显著(P < 0.05), 加粗斜体表示弱显著(P < 0.10)。
Table 1 Effects of origin (i.e. introduced vs. native) and simulated daily warming (i.e. control vs. warming) and their interaction on fitness traits (total biomass, storage root biomass (STB)), functional traits (branching intensity (BI), specific stem length (SSL), root to shoot ratio (RSR) and specific leaf area (SLA)) and herbivorous effect (relative feeding area (RFA) and stem-tip feeding proportion (SFP)) of Alternanthera philoxeroides. Significant effects are marked in bold and marginally significant effects are marked in italics and bold.
| 变异来源Source of variation | 总生物量 Total biomass | 贮藏根生物量 STB | 分枝强度 BI | 比茎长 SSL | 根冠比 RSR | 比叶面积 SLA | 相对取食面积 RFA | 茎端取食率 SFP | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| χ2 | P | χ2 | P | χ2 | P | χ2 | P | χ2 | P | χ2 | P | χ2 | P | χ2 | P | |
| 增温 Warming (W) | 5.165 | 0.023 | 3.207 | 0.073 | 4.239 | 0.040 | 0.761 | 0.383 | 0.343 | 0.558 | 0.021 | 0.884 | 0.083 | 0.773 | 10.552 | 0.001 |
| 来源地 Origin (O) | 1.523 | 0.217 | 8.756 | 0.003 | 8.023 | 0.005 | 8.092 | 0.004 | 5.758 | 0.016 | 4.665 | 0.031 | 1.837 | 0.175 | 3.680 | 0.055 |
| W × O | 2.986 | 0.084 | 0.244 | 0.621 | 1.077 | 0.299 | 3.453 | 0.063 | 0.913 | 0.339 | 0.365 | 0.546 | 3.682 | 0.055 | 1.772 | 0.183 |
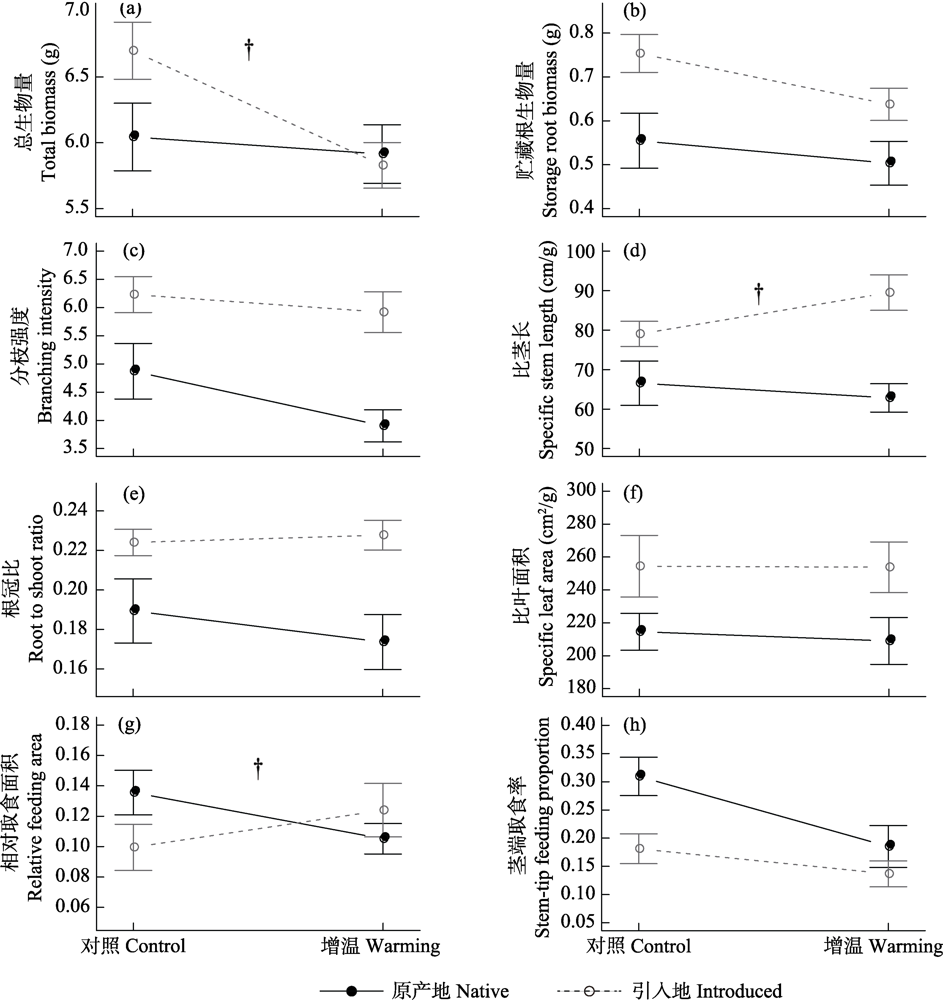
图1 喜旱莲子草引入地(中国)和原产地(阿根廷)种群对模拟全天增温的响应。误差棒代表95%的置信区间; ?表示全天增温处理与来源地的交互作用弱显著。
Fig. 1 Responses of introduced (China) and native (Argentina) populations of Alternanthera philoxeroides to simulated daily warming. Error bars indicate 95% confidence intervals, and ? indicate marginally significant differences in responses between introduced and native populations.
| [1] | Beckmann M, Bruelheide H, Erfmeier A (2011) Germination responses of three grassland species differ between native and invasive origins. Ecological Research, 26,763-771. |
| [2] |
Blumenthal DM, Kray JA, Ortmans W, Ziska LH, Pendall E (2016) Cheatgrass is favored by warming but not CO2 enrichment in a semi-arid grassland. Global Change Biology, 22,3026-3038.
DOI URL PMID |
| [3] | Buckland SM, Thompson K, Hodgson JG, Grime JP (2001) Grassland invasions: Effects of manipulations of climate and management. Journal of Applied Ecology, 38,301-309. |
| [4] | Burgin S, Norris A (2008) Alligator weed (Alternanthera philoxeroides) in New South Wales, Australia: A status report. Weed Biology and Management, 8,284-290. |
| [5] | Chu YM, Yang J, Li JJ, Peng PH (2014) Three warming scenarios differentially affect the morphological plasticity of an invasive herb Alternanthera philoxeroides. Acta Ecologica Sinica, 34,1411-1417. (in Chinese with English abstract) |
| [ 褚延梅, 杨健, 李景吉, 彭培好 (2014) 三种增温情景对入侵植物空心莲子草形态可塑性的影响. 生态学报, 34,1411-1417. ] | |
| [6] | Daehler CC (2003) Performance comparisons of co-occurring native and alien invasive plants: Implications for conservation and restoration. Annual Review of Ecology, Evolution, and Systematics, 34,183-211. |
| [7] |
Dong BC, Alpert P, Zhang Q, Yu FH (2015) Clonal integration in homogeneous environments increases performance of Alternanthera philoxeroides. Oecologia, 179,393-403.
URL PMID |
| [8] | Geng YP, Pan XY, Xu CY, Zhang WJ, Li B, Chen JK (2006) Phenotypic plasticity of invasive Alternanthera philoxeroides in relation to different water availability, compared to its native congener. Acta Oecologica, 30,380-385. |
| [9] | Griffith AB, Andonian K, Weiss CP, Loik ME (2014) Variation in phenotypic plasticity for native and invasive populations of Bromus tectorum. Biological Invasions, 16,2627-2638. |
| [10] |
He WM, Li JJ, Peng PH (2012) Simulated warming differentially affects the growth and competitive ability of Centaurea maculosa populations from home and introduced ranges. PLoS ONE, 7,e31170.
DOI URL PMID |
| [11] | Hyldgaard B, Brix H (2012) Intraspecies differences in phenotypic plasticity: Invasive versus non-invasive populations of Ceratophyllum demersum. Aquatic Botany, 97,49-56. |
| [12] | Julien MH, Skarratt B, Maywald GF (1995) Potential geographical-distribution of alligator weed and its biological-control Agasicles hygrophila. Journal of Aquatic Plant Management, 33,55-60. |
| [13] |
Kim YU, Lee BW (2019) Differential mechanisms of potato yield loss induced by high day and night temperatures during tuber initiation and bulking: Photosynthesis and tuber growth. Frontiers in Plant Science, 10,300.
URL PMID |
| [14] | Lemoine NP, Drews WA, Burkepile DE, Parker JD (2013) Increased temperature alters feeding behavior of a generalist herbivore. Oikos, 122,1669-1678. |
| [15] | Lin DL, Xia JY, Wan SQ (2010) Climate warming and biomass accumulation of terrestrial plants: A meta-analysis. New Phytologist, 188,187-198. |
| [16] | Liu M, Pan XY, Zhang ZJ, van Kleunen M, Li B (2020) Testing the shifting defense hypothesis for constitutive and induced resistance and tolerance. Journal of Pest Science, 93,355-364. |
| [17] | Liu M, Zhou F, Pan XY, Zhang ZJ, Traw MB, Li B (2018) Specificity of herbivore-induced responses in an invasive species, Alternanthera philoxeroides (Alligator weed). Ecology and Evolution, 8,59-70. |
| [18] | Luo YQ (2007) Terrestrial carbon-cycle feedback to climate warming. Annual Review of Ecology Evolution and Systematics, 38,683-712. |
| [19] | McKinney ML (2006) Urbanization as a major cause of biotic homogenization. Biological Conservation, 127,247-260. |
| [20] | Nguyen HM, Yadav NS, Barak S, Lima FP, Sapir Y, Winters G (2020) Responses of invasive and native populations of the seagrass Halophila stipulacea to simulated climate change. Frontiers in Marine Science, 6,812. |
| [21] |
Nicotra AB, Atkin OK, Bonser SP, Davidson AM, Finnegan EJ, Mathesius U, Poot P, Purugganan MD, Richards CL, Valladares F, van Kleunen M (2010) Plant phenotypic plasticity in a changing climate. Trends in Plant Science, 15,684-692.
DOI URL PMID |
| [22] | Pan XY, Geng YP, Sosa A, Zhang WJ, Li B, Chen JK (2007) Invasive Alternanthera philoxeroides: Biology, ecology and management. Acta Phytotaxonomica Sinica, 45,884-900. (in Chinese with English abstract) |
| [ 潘晓云, 耿宇鹏, Alejandro Sosa, 张文驹, 李博, 陈家宽 (2007) 入侵植物喜旱莲子草——生物学、生态学及管理. 植物分类学报, 45,884-900. ] | |
| [23] | Pan XY, Jia X, Fu DJ, Li B (2013) Geographical diversification of growth-defense strategies in an invasive plant. Journal of Systematics and Evolution, 51,308-317. |
| [24] | Paudel S, Lin PA, Hoover K, Felton GW, Rajotte EG (2020) Asymmetric responses to climate change: Temperature differentially alters herbivore salivary elicitor and host plant responses to herbivory. Journal of Chemical Ecology, 46,891-905. |
| [25] |
Peng SS, Piao SL, Ciais P, Myneni RB, Chen AP, Chevallier F, Dolman AJ, Janssens IA, Peñuelas J, Zhang GX, Vicca S, Wan SQ, Wang SP, Zeng H (2013) Asymmetric effects of daytime and night-time warming on Northern Hemisphere vegetation. Nature, 501,88-92.
URL PMID |
| [26] | Peng Y, Yang JX, Zhou XH, Peng PH, Li JJ, Zhang SM, He WM (2019) An invasive population of Solidago canadensis is less sensitive to warming and nitrogen-addition than its native population in an invaded range. Biological Invasions, 21,151-162. |
| [27] | Pereira FMV, Rosa E, Fahey JW, Stephenson KK, Carvalho R, Aires A (2002) Influence of temperature and ontogeny on the levels of glucosinolates in broccoli (Brassica oleracea var. italica) sprouts and their effect on the induction of mammalian phase 2 enzymes. Journal of Agricultural and Food Chemistry, 50,6239-6244. |
| [28] | Portela R, Dong BC, Yu FH, Barreiro R, Roiloa SR, Silva Matos DM (2020) Trans-generational effects in the clonal invader Alternanthera philoxeroides. Journal of Plant Ecology, 13,122-129. |
| [29] | Prange RK, McRae KB, Midmore DJ, Deng RB (1990) Reduction in potato growth at high temperature: Role of photosynthesis and dark respiration. American Potato Journal, 67,357-369. |
| [30] | R Development Core Team (2020) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna. |
| [31] |
Richards CL, Bossdorf O, Muth NZ, Gurevitch J, Pigliucci M (2006) Jack of all trades, master of some? On the role of phenotypic plasticity in plant invasions. Ecology Letters, 9,981-993.
URL PMID |
| [32] | Shipley B, Meziane D (2002) The balanced-growth hypothesis and the allometry of leaf and root biomass allocation. Functional Ecology, 16,326-331. |
| [33] | Wang Q, Tang Y, Xie T, Wang H (2017) Distinctive responses of photosynthetic characteristics to warming of invasive Alternanthera philoxeroides and native Sambucus chinensis. Acta Ecologica Sinica, 37,770-777. (in Chinese with English abstract) |
| [ 王琼, 唐娅, 谢涛, 王辉 (2017) 入侵植物喜旱莲子草和本地种接骨草光合生理特征对增温响应的差异. 生态学报, 37,770-777. ] | |
| [34] | Wu H, Zhang C, Dai WK (2020) Interactive effects of climate warming and species diversity on the invasiveness of the alien weed Alternanthera philoxeroides. Acta Prataculturae Sinica, 29,38-48. (in Chinese with English abstract) |
| [ 吴昊, 张辰, 代文魁 (2020) 气候变暖和物种多样性交互效应对空心莲子草入侵的影响. 草业学报, 29,38-48. ] | |
| [35] | Yu LR, Zhu ZC, Pan XY (2020) Phenotypic plasticity of Alternanthera philoxeroides in response to root neighbors of kin: Introduced vs. native genotypes. Biodiversity Science, 28,651-657. (in Chinese with English abstract) |
| [ 于良瑞, 朱政财, 潘晓云 (2020) 喜旱莲子草对同基因型邻体根系的表型可塑性: 入侵地和原产地的比较. 生物多样性, 28,651-657. ] | |
| [36] |
Zhang R, Leshak A, Shea K (2012) Decreased structural defence of an invasive thistle under warming. Plant Biology, 14,249-252.
URL PMID |
| [37] | Zhang ZJ, Zhou F, Pan XY, Kleunen M, Liu M, Li B (2019) Evolution of increased intraspecific competitive ability following introduction: The importance of relatedness among genotypes. Journal of Ecology, 107,387-395. |
| [38] | Zhang ZY, Zhang ZJ, Pan XY (2015) Phenotypic plasticity of Alternanthera philoxeroides in response to shading: Intro- duced vs. native populations. Biodiversity Science, 23,18-22. (in Chinese with English abstract) |
| [ 张紫妍, 张致杰, 潘晓云 (2015) 喜旱莲子草对遮荫的可塑性反应: 入侵地与原产地种群的比较. 生物多样性, 23,18-22. ] | |
| [39] | Zhou F, Zhang ZJ, Liu M, Pan XY (2017) Effects of nutrient levels on defense against specialist insects in an invasive alligator weed. Biodiversity Science, 25,1276-1284. (in Chinese with English abstract) |
| [ 周方, 张致杰, 刘木, 潘晓云 (2017) 养分影响入侵种喜旱莲子草对专食性天敌的防御. 生物多样性, 25,1276-1284. ] | |
| [40] | Zhou XH, He WM (2020) Climate warming facilitates seed germination in native but not invasive solidago canadensis populations. Frontiers in Ecology and Evolution, 8,595214. |
| [1] | 夏东坡, 李静, 田军东, 黄中豪, 伍乘风, 陈仕望, 李进华. 中国野生猕猴属动物的研究进展[J]. 生物多样性, 2026, 34(3): 25460-. |
| [2] | 曾文敏, 杨霞, 邹成玉, 李彦欣, 王贤楠, 魏依婷, 徐艳琴, 周银. 植物复合群的形成机制与分类学研究进展[J]. 生物多样性, 2026, 34(2): 25321-. |
| [3] | 孙婷婷, 张彦文. 花粉二型性研究进展[J]. 生物多样性, 2025, 33(7): 25031-. |
| [4] | 张浩斌, 肖路, 刘艳杰. 夜间灯光对外来入侵植物和本地植物群落多样性和生长的影响[J]. 生物多样性, 2025, 33(4): 24553-. |
| [5] | 李燚, 赵艳超, 陈立同. 高寒草甸α与β多样性对增温的响应受海拔和土壤养分水平调控[J]. 生物多样性, 2025, 33(12): 25062-. |
| [6] | 陈楠, 张全国. 实验进化研究途径[J]. 生物多样性, 2024, 32(9): 24171-. |
| [7] | 何花, 谭敦炎, 杨晓琛. 被子植物隐性雌雄异株性系统的多样性、系统演化及进化意义[J]. 生物多样性, 2024, 32(6): 24149-. |
| [8] | 姚祝, 魏雪, 马金豪, 任晓, 王玉英, 胡雷, 吴鹏飞. 气候暖湿化对高寒草甸土壤线虫群落的短期影响[J]. 生物多样性, 2024, 32(5): 23483-. |
| [9] | 曲锐, 左振君, 王有鑫, 张良键, 吴志刚, 乔秀娟, 王忠. 基于元素组的生物地球化学生态位及其在不同生态系统中的应用[J]. 生物多样性, 2024, 32(4): 23378-. |
| [10] | 李庆多, 栗冬梅. 全球蝙蝠巴尔通体流行状况分析[J]. 生物多样性, 2023, 31(9): 23166-. |
| [11] | 陈哲涵, 尹进, 叶吉, 刘冬伟, 毛子昆, 房帅, 蔺菲, 王绪高. 增温对东北温带次生林草本群落季节动态的影响[J]. 生物多样性, 2023, 31(5): 23059-. |
| [12] | 邵雯雯, 范国祯, 何知舟, 宋志平. 多地同质园实验揭示普通野生稻的表型可塑性与本地适应性[J]. 生物多样性, 2023, 31(3): 22311-. |
| [13] | 沈诗韵, 潘远飞, 陈丽茹, 土艳丽, 潘晓云. 喜旱莲子草原产地和入侵地种群的植物-土壤反馈差异[J]. 生物多样性, 2023, 31(3): 22436-. |
| [14] | 戚海迪, 张定海, 单立山, 陈国鹏, 张勃. 昆虫病原真菌感染昆虫宿主的机制和宿主昆虫的防御策略研究进展[J]. 生物多样性, 2023, 31(11): 23273-. |
| [15] | 俄广旭, 白天天, 朱振宇, 郭雪峰. 动物消化道微生物多样性与宿主协同进化关系的研究进展[J]. 生物多样性, 2023, 31(11): 23214-. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
备案号:京ICP备16067583号-7
Copyright © 2026 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn
![]()