
生物多样性 ›› 2024, Vol. 32 ›› Issue (6): 24149. DOI: 10.17520/biods.2024149 cstr: 32101.14.biods.2024149
何花, 谭敦炎, 杨晓琛*( )(
)( )
)
收稿日期:2024-04-21
接受日期:2024-06-11
出版日期:2024-06-20
发布日期:2024-06-22
通讯作者:
* E-mail: xcyang@jsu.edu.cn基金资助:
Hua He, Dunyan Tan, Xiaochen Yang*()()
Received:2024-04-21
Accepted:2024-06-11
Online:2024-06-20
Published:2024-06-22
Contact:
* E-mail: xcyang@jsu.edu.cn摘要:
隐性雌雄异株指植株表型为非雌雄异株, 但形态上两性花通常雄性不育或雌性不育, 植株个体实际仅行使雌性或雄性功能。两性花植株向雌雄异株的转变在被子植物中广泛存在, 隐性雌雄异株作为潜在的中间阶段对于理解植物性系统的演化具有重要意义。据APG IV分类系统, 这些物种在被子植物的22目36科65属约221种植物中有过报道, 分别约占被子植物总目数的34.4%、总科数的8.7%、总属数的0.5%和总种数的0.1%; 多为分布于热带或亚热带地区依靠生物传粉的木本植物, 且至少存在花蜜或花粉作为访花报酬。该性系统形态学的性表达类型多样: 可分为雄花两性花异株(类型I)、两性花植株(类型II)、雌花两性花异株(类型III)及其他类型(类型IV) 4种, 分别约占已报道隐性雌雄异株物种数的48.9%、47.5%、2.7%和0.9%。从系统发育看, 该性系统在被子植物木兰分支、单子叶植物分支及真双子叶植物分支均有分布, 且主要分布于较为进化的核心真双子叶植物类群中。隐性雌雄异株物种不育性器官的进化意义存在多种假说, 包括祖先假说、遗传约束假说、促进风媒花粉落置假说、传粉者吸引假说、拟态与欺骗假说等, 但相关实证研究较少。本文对隐性雌雄异株物种不同类群的性表达形式、不育性器官败育类型进行了归纳, 并对具该性系统的类群在被子植物中的分布与系统演化进行了分析与总结, 同时对有关隐性雌雄异株物种不育性器官进化意义的5个假说进行了介绍与评价, 最后对今后的相关研究方向进行了展望。以期为深入研究被子植物隐性雌雄异株性系统的进化式样与机制研究提供理论资料。
何花, 谭敦炎, 杨晓琛 (2024) 被子植物隐性雌雄异株性系统的多样性、系统演化及进化意义. 生物多样性, 32, 24149. DOI: 10.17520/biods.2024149.
Hua He, Dunyan Tan, Xiaochen Yang (2024) Cryptic dioecy in angiosperms: Diversity, phylogeny and evolutionary significance. Biodiversity Science, 32, 24149. DOI: 10.17520/biods.2024149.
| 目 Order | 科 Family | 属 Genus | 种 Species | 参考文献 Reference |
|---|---|---|---|---|
性表达类型I Sex expression type I 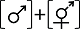 | ||||
| 木兰目 Magnoliales | 木兰科 Magnoliaceae | 拟单性木兰属 Parakmeria | 峨眉拟单性木兰 P. omeiensis | 余道平等, |
| 棕榈目 Arecales | 棕榈科 Arecaceae | 海枣属 Phoenix | 海枣 P. dactylifera | 陆静等, |
| 毛茛目 Ranunculales | 毛茛科 Ranunculaceae | 唐松草属 Thalictrum | T. macrostylum, T. polygamum, 柔毛唐松草 T. pubescens | Melampy & Hayworth, |
| 昆栏树目 Trochodendrales | 昆栏树科 Trochodendraceae | 昆栏树属 Trochodendron | 昆栏树 T. aralioides | Keng, |
| 五桠果目 Dilleniales | 五桠果科 Dilleniaceae | 锡叶藤属 Tetracera | 锡叶藤属14种 Tetracera sp. 1-14 | Kubitzki & Baretta-Kuipers, |
| 葡萄目 Vitales | 葡萄科 Vitaceae | 葡萄属 Vitis | 夏葡萄 V. aestivalis, 河岸葡萄 V. riparia, V. vinifera ssp. silvestris | Brantjes, |
| 金虎尾目 Malpighiales | 藤黄科 Clusiaceae | 藤黄属 Garcinia | G. brasiliensis | Leal et al, |
| 红厚壳科 Calophyllaceae | 南美杏属 Mammea | M. acuminata, M. africana, 马米杏 M. americana, M. sp. aff. bongo, M. eugenioides var. subsessilifolia, M. punctata, M. cf. punctata, M. siamensis | Dunthorn, | |
| 金虎尾科Malpighiaceae | 折背藤属 Madagasikaria | M. andersonii | Davis CC, | |
| 辐翅藤属 Rhynchophora | R. phillipsonii | Anderson WR, | ||
| 豆目 Fabales | 豆科 Fabaceae | 皂荚属 Gleditsia | 南美皂荚 G. amorphoides | Cerino et al, |
| 桃金娘目 Myrtales | 桃金娘科 Myrtaceae | 子楝树属 Decaspermum | 五瓣子楝树 D. parviflorum | Kevan & Lack, |
| 番樱桃属 Eugenia | E. albanensis, E. capensis, E. cf. mossambicensis, E. erythrophylla, E. natalitia, E. simii, E. umtamvunensis, E. verdoorniae, E. woodii, E. zeyheri, E.zuluensis, 番樱桃属3种 Eugenia sp. 1-3 | van Wyk & Dedekind, van der Merwe et al, | ||
| 无患子目 Sapindales | 无患子科 Sapindaceae | 干果木属 Xerospermum | X. intermedium | Appanah, |
| 石竹目 Caryophyllales | 石竹科 Caryophyllaceae | 冰漆姑属 Honckenya | H. peploides var. major | Tsukui & Sugawara, |
| 山茱萸目 Cornales | 蓝果树科 Nyssaceae | 蓝果树属 Nyssa | 云南蓝果树 N. yunnanensis | Sun et al, |
| 杜鹃花目 Ericales | 柿科 Ebenaceae | 柿属 Diospyros | 君迁子 D. lotus | 陆静等, |
| 山矾科 Symplocaceae | 山矾属 Symplocos | S. celastrinea, S. crenata, S. itatiaiae, S. lanceolata, S. mosenii, S. rhamnifolia, S. saxatilis, S. tetrandra, S. variabilis, S. inopinata, S. kleinii, S. occulta, 微毛山矾 S. wikstroemiifolia | Aranha Filho et al, | |
| 猕猴桃科 Actinidiaceae | 猕猴桃属 Actinidia | 软枣猕猴桃 A. arguta, 中华猕猴桃 A. chinensis, 美味猕猴桃 A. deliciosa var. deliciosa, 毛花猕猴桃 A. eriantha, 绵毛猕猴桃 A. fulvicoma var. lanata, 革叶猕猴桃 A. rubricaulis var. coriacea, 对萼猕猴桃 A. valvata, 葛枣猕猴桃 A. polygama | unpublished | |
| 水东哥属 Saurauia | S. bullosa, S. brachybotrys, S. excelsa, S. humboldtiana, S. tomentosa, S. putumayonis, S. ursina, S. veraguensis | Soejarto, | ||
| 伞形目 Apiales | 毛柴木科 Pennantiaceae | 毛柴木属 Pennantia | 毛柴木属3种 Pennantia sp. 1-3 | Baylis, |
| 五加科 Araliaceae | 南洋参属 Polyscias | P. dzumacensis, P. microbotrys, P. pancheri, P. subincisa | Schlessman et al, | |
| 紫草目 Boraginales | 紫草科 Boraginaceae | 破布木属 Cordia | C. crenata, C. cf. grandicalyx, C. sinensis | Pätzold et al, |
| 龙胆目 Gentianales | 茜草科 Rubiaceae | 木巴戟属 Morinda | 鸡眼藤 M. parvifolia | Liu et al, |
| 唇形目 Lamiales | 木樨科 Oleaceae | 梣属 Fraxinus | 维斯谷归化种群中的花梣 Fraxinus ornus in naturalized Vis Valley populations | Vernet et al, |
| 茄目 Solanales | 茄科 Solanaceae | 茄属 Solanum | S. asymmetriphyllum, S. carduiforme, S. cataphractum, S. cowiei, S. cunninghamii, S. dioicum, S. leopoldense, S. ossicruentum, S. petraeum, S. polygamum, S. tudununggae, S. vansittartensis, S. sejunctum | Anderson & Symon, |
性表达类型II Sex expression type II 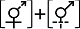 | ||||
| 蔷薇目 Rosales | 蔷薇科 Rosaceae | 蔷薇属 Rosa | 草原玫瑰 R. setigera | Kevan et al, |
| 桃金娘目 Myrtales | 桃金娘科 Myrtaceae | 多香果属 Pimenta | 多香果 P. dioica | Boyd & Benkeblia, |
| 无患子目 Sapindales | 无患子科 Sapindaceae | 槭属 Acer | 红花槭 A. rubrum | Batra, |
| 楝科 Meliaceae | 杜楝属 Turraea | T. dargentiana | Baider & Florens, | |
| 锦葵目 Malvales | 瑞香科 Thymelaeaceae | 荛花属 Wikstroemia | 夏威夷岛和可爱岛中荛花属种群 Wikstroemia populations on Kauai, Hawaii | Mayer & Charlesworth, |
| 锦葵科 Malvaceae | 非洲芙蓉属 Dombeya | D. ciliata, D. delislei, D. elegans, D. ficulnea, D. formosa, D. pilosa, D. punctata, D. reclinata | Humeau et al, Le Péchon et al, | |
| 石竹目 Caryophyllales | 仙人掌科 Cactaceae | 旗号掌属 Consolea | C. corallicola, C. millspaughii, 鱼鳞掌 C. moniliformis, C. nashii, C. picardae, C. rubescens, C. spinosissima | Rebman & Pinkava, |
| 鹿角柱属 Echinocereus | 赤花虾 E. coccineus, E. mombergerianus, E. pacificus, 多刺虾鹿角柱 E. polyacanthus | Hernández-Cruz et al, | ||
| 仙人掌属 Opuntia | O. quitensis, O. robusta, O. stenopetala | Anderson EF, | ||
| 摩天柱属 Pachycereus | 武伦柱 P. pringlei | Fleming, | ||
| 木麒麟属 Pereskia | P. portulacifolia, P. marcanoi, P. quisqueyana, P. zinniiflora | Areces-Mallea, | ||
| 杜鹃花目 Ericales | 杜鹃花科 Ericaceae | 南白珠属 Pernettya | P. rigida | Anderson et al, |
| 菊目 Asterales | 菊科 Asteraceae | 假泽兰属 Mikania | 假泽兰属5种 Mikania 5 sp. 1-5 | Holmes, |
| 茶茱萸目 Icacinales | 茶茱萸科 Icacinaceae | 假柴龙树属 Nothapodytes | 臭味假柴龙树 N. nimmoniana | Sharma et al, |
| 龙胆目 Gentianales | 茜草科 Rubiaceae | 舒榄属 Bertiera | B. borbonica | Pailler et al, |
| 弯管花属 Chassalia | C. corallioides | Pailler et al, | ||
| 牛眼棠属 Cordiera | C. concolor | Judkevich et al, | ||
| 靛榄属 Genipa | 靛榄 G. americana | Judkevich et al, | ||
| 蓝茜树属 Randia | R. calycina, R. heteromera | Judkevich et al, | ||
| 木巴戟属 Morinda | 须弥巴戟 M. villosa | Sugawara et al, | ||
| 玉叶金花属 Mussaenda | 短裂玉叶金花 M. breviloba, 尾裂玉叶金花 M. caudatiloba, 仁昌玉叶金花 M. chingii, 展枝玉叶金花 M. divaricata, 楠藤 M. erosa, 海南玉叶金花 M. hainanensis, 粗毛玉叶金花 M. hirsutula, 红毛玉叶金花 M. hossei, 广西玉叶金花 M. kwangsiensis, 广东玉叶金花 M. kwangtungensis, 疏花玉叶金花 M. laxiflora, 长瓣玉叶金花 M. longipetala, 多毛玉叶金花 M. mollissima, 小玉叶金花 M. parviflora, M. pilosissima, 屏边玉叶金花 M. pingbianensis, 玉叶金花 M. pubescens, 白花玉叶金花 M. pubescens var. alba, 贡山玉叶金花 M. treutleri, 玉叶金花属5种 Mussaenda sp. 1-5 | 邓小芳, | ||
| 沙九节属 Pagamea | P. duckei | Terra-Araujo et al, | ||
| 九节属 Psychotria | 九节 P. rubra | Watanabe et al, | ||
| 马钱科 Loganiaceae | 髯管花属 Geniostoma | G. borbonica | Humeau et al, | |
| 顶髯管花属 Labordia | L. cyrtandrae, L. degeneri, L. fagraeoidea, L. hedyosmifolia, L. hirtella, L. hosakana, L. lydgatei, L. pumila, L. sessilis, L.venosa, L. waialealae, L. waiolani | Motley & Carr, | ||
| 夹竹桃科 Apocynaceae | 萝芙木属 Rauvolfia | R. sellowii | Souto & Koch, | |
| 唇形目 Lamiales | 马鞭草科 Verbenaceae | 琴木属 Citharexylum | C. myrianthum | Rocca & Sazima, |
| 唇形科 Lamiaceae | 紫珠属 Callicarpa | C. glabra, C. nishimurae, C. subpubescens | Kawakubo, | |
| 茄目 Solanales | 茄科 Solanaceae | 山盏茄属 Deprea | D. paneroi | Sawyer & Anderson, |
| 巫灵木属 Dunalia | D. solanacea | Hepper, | ||
| 枸杞属 Lycium | L. arenicolum, L. horridum, L. tetrandrum, L. villosum | Spies et al, | ||
| 茄目 Solanales | 茄科 Solanaceae | 茄属 Solanum | S. appendiculatum, S. campanulatum, S. confertiseriatum, S. crotonoides, S. conocarpum | Knapp et al, Anderson et al, |
| 串钟花属 Symonanthus | S. aromaticus, S. bancroftii | Haegi, | ||
| 睡茄属 Withania | W. aristata, W. coagulans | Sawyer & Anderson, | ||
性表达类型III Sex expression type III 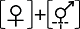 | ||||
| 金虎尾目 Malpighiales | 金虎尾科Malpighiaceae | 异金英属 Spachea | S. membranacea | Steiner, |
| 石竹目 Caryophyllales | 石竹科Caryophyllaceae | 洋漆姑属 Schiedea | S. adamantis | Mayer & Charlesworth, |
| 仙人掌科 Cactaceae | 圆柱掌属 Cylindropuntia | C. wolfii | Ramadoss et al, | |
| 裸萼球属 Gymnocalycium | 罗星丸 G. bruchii | Delbón et al, | ||
| 杜鹃花目 Ericales | 山榄科 Sapotaceae | 星苹果属 Chrysophyllum | C. gonocarpum | Judkevich et al, |
| 杜鹃花科 Ericaceae | 岩梨属 Epigaea | 岩梨 E. repens | Clay & Ellstrand, | |
性表达类型IV Sex expression type IV (1) 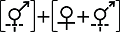 (2) (2) 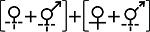 | ||||
| 菊目 Asterales | 菊科 Asteraceae | 蜂斗菜属 Petasites | (1)蜂斗菜 P. japonicas; (2)毛裂蜂斗菜 P. tricholobus | Sakai et al, |
表1 隐性雌雄异株的性表达类型及其在被子植物中的分布
Table 1 Sex expression types of cryptic dioecy and their distribution in angiosperms
| 目 Order | 科 Family | 属 Genus | 种 Species | 参考文献 Reference |
|---|---|---|---|---|
| 性表达类型I Sex expression type I | ||||
| 木兰目 Magnoliales | 木兰科 Magnoliaceae | 拟单性木兰属 Parakmeria | 峨眉拟单性木兰 P. omeiensis | 余道平等, |
| 棕榈目 Arecales | 棕榈科 Arecaceae | 海枣属 Phoenix | 海枣 P. dactylifera | 陆静等, |
| 毛茛目 Ranunculales | 毛茛科 Ranunculaceae | 唐松草属 Thalictrum | T. macrostylum, T. polygamum, 柔毛唐松草 T. pubescens | Melampy & Hayworth, |
| 昆栏树目 Trochodendrales | 昆栏树科 Trochodendraceae | 昆栏树属 Trochodendron | 昆栏树 T. aralioides | Keng, |
| 五桠果目 Dilleniales | 五桠果科 Dilleniaceae | 锡叶藤属 Tetracera | 锡叶藤属14种 Tetracera sp. 1-14 | Kubitzki & Baretta-Kuipers, |
| 葡萄目 Vitales | 葡萄科 Vitaceae | 葡萄属 Vitis | 夏葡萄 V. aestivalis, 河岸葡萄 V. riparia, V. vinifera ssp. silvestris | Brantjes, |
| 金虎尾目 Malpighiales | 藤黄科 Clusiaceae | 藤黄属 Garcinia | G. brasiliensis | Leal et al, |
| 红厚壳科 Calophyllaceae | 南美杏属 Mammea | M. acuminata, M. africana, 马米杏 M. americana, M. sp. aff. bongo, M. eugenioides var. subsessilifolia, M. punctata, M. cf. punctata, M. siamensis | Dunthorn, | |
| 金虎尾科Malpighiaceae | 折背藤属 Madagasikaria | M. andersonii | Davis CC, | |
| 辐翅藤属 Rhynchophora | R. phillipsonii | Anderson WR, | ||
| 豆目 Fabales | 豆科 Fabaceae | 皂荚属 Gleditsia | 南美皂荚 G. amorphoides | Cerino et al, |
| 桃金娘目 Myrtales | 桃金娘科 Myrtaceae | 子楝树属 Decaspermum | 五瓣子楝树 D. parviflorum | Kevan & Lack, |
| 番樱桃属 Eugenia | E. albanensis, E. capensis, E. cf. mossambicensis, E. erythrophylla, E. natalitia, E. simii, E. umtamvunensis, E. verdoorniae, E. woodii, E. zeyheri, E.zuluensis, 番樱桃属3种 Eugenia sp. 1-3 | van Wyk & Dedekind, van der Merwe et al, | ||
| 无患子目 Sapindales | 无患子科 Sapindaceae | 干果木属 Xerospermum | X. intermedium | Appanah, |
| 石竹目 Caryophyllales | 石竹科 Caryophyllaceae | 冰漆姑属 Honckenya | H. peploides var. major | Tsukui & Sugawara, |
| 山茱萸目 Cornales | 蓝果树科 Nyssaceae | 蓝果树属 Nyssa | 云南蓝果树 N. yunnanensis | Sun et al, |
| 杜鹃花目 Ericales | 柿科 Ebenaceae | 柿属 Diospyros | 君迁子 D. lotus | 陆静等, |
| 山矾科 Symplocaceae | 山矾属 Symplocos | S. celastrinea, S. crenata, S. itatiaiae, S. lanceolata, S. mosenii, S. rhamnifolia, S. saxatilis, S. tetrandra, S. variabilis, S. inopinata, S. kleinii, S. occulta, 微毛山矾 S. wikstroemiifolia | Aranha Filho et al, | |
| 猕猴桃科 Actinidiaceae | 猕猴桃属 Actinidia | 软枣猕猴桃 A. arguta, 中华猕猴桃 A. chinensis, 美味猕猴桃 A. deliciosa var. deliciosa, 毛花猕猴桃 A. eriantha, 绵毛猕猴桃 A. fulvicoma var. lanata, 革叶猕猴桃 A. rubricaulis var. coriacea, 对萼猕猴桃 A. valvata, 葛枣猕猴桃 A. polygama | unpublished | |
| 水东哥属 Saurauia | S. bullosa, S. brachybotrys, S. excelsa, S. humboldtiana, S. tomentosa, S. putumayonis, S. ursina, S. veraguensis | Soejarto, | ||
| 伞形目 Apiales | 毛柴木科 Pennantiaceae | 毛柴木属 Pennantia | 毛柴木属3种 Pennantia sp. 1-3 | Baylis, |
| 五加科 Araliaceae | 南洋参属 Polyscias | P. dzumacensis, P. microbotrys, P. pancheri, P. subincisa | Schlessman et al, | |
| 紫草目 Boraginales | 紫草科 Boraginaceae | 破布木属 Cordia | C. crenata, C. cf. grandicalyx, C. sinensis | Pätzold et al, |
| 龙胆目 Gentianales | 茜草科 Rubiaceae | 木巴戟属 Morinda | 鸡眼藤 M. parvifolia | Liu et al, |
| 唇形目 Lamiales | 木樨科 Oleaceae | 梣属 Fraxinus | 维斯谷归化种群中的花梣 Fraxinus ornus in naturalized Vis Valley populations | Vernet et al, |
| 茄目 Solanales | 茄科 Solanaceae | 茄属 Solanum | S. asymmetriphyllum, S. carduiforme, S. cataphractum, S. cowiei, S. cunninghamii, S. dioicum, S. leopoldense, S. ossicruentum, S. petraeum, S. polygamum, S. tudununggae, S. vansittartensis, S. sejunctum | Anderson & Symon, |
| 性表达类型II Sex expression type II | ||||
| 蔷薇目 Rosales | 蔷薇科 Rosaceae | 蔷薇属 Rosa | 草原玫瑰 R. setigera | Kevan et al, |
| 桃金娘目 Myrtales | 桃金娘科 Myrtaceae | 多香果属 Pimenta | 多香果 P. dioica | Boyd & Benkeblia, |
| 无患子目 Sapindales | 无患子科 Sapindaceae | 槭属 Acer | 红花槭 A. rubrum | Batra, |
| 楝科 Meliaceae | 杜楝属 Turraea | T. dargentiana | Baider & Florens, | |
| 锦葵目 Malvales | 瑞香科 Thymelaeaceae | 荛花属 Wikstroemia | 夏威夷岛和可爱岛中荛花属种群 Wikstroemia populations on Kauai, Hawaii | Mayer & Charlesworth, |
| 锦葵科 Malvaceae | 非洲芙蓉属 Dombeya | D. ciliata, D. delislei, D. elegans, D. ficulnea, D. formosa, D. pilosa, D. punctata, D. reclinata | Humeau et al, Le Péchon et al, | |
| 石竹目 Caryophyllales | 仙人掌科 Cactaceae | 旗号掌属 Consolea | C. corallicola, C. millspaughii, 鱼鳞掌 C. moniliformis, C. nashii, C. picardae, C. rubescens, C. spinosissima | Rebman & Pinkava, |
| 鹿角柱属 Echinocereus | 赤花虾 E. coccineus, E. mombergerianus, E. pacificus, 多刺虾鹿角柱 E. polyacanthus | Hernández-Cruz et al, | ||
| 仙人掌属 Opuntia | O. quitensis, O. robusta, O. stenopetala | Anderson EF, | ||
| 摩天柱属 Pachycereus | 武伦柱 P. pringlei | Fleming, | ||
| 木麒麟属 Pereskia | P. portulacifolia, P. marcanoi, P. quisqueyana, P. zinniiflora | Areces-Mallea, | ||
| 杜鹃花目 Ericales | 杜鹃花科 Ericaceae | 南白珠属 Pernettya | P. rigida | Anderson et al, |
| 菊目 Asterales | 菊科 Asteraceae | 假泽兰属 Mikania | 假泽兰属5种 Mikania 5 sp. 1-5 | Holmes, |
| 茶茱萸目 Icacinales | 茶茱萸科 Icacinaceae | 假柴龙树属 Nothapodytes | 臭味假柴龙树 N. nimmoniana | Sharma et al, |
| 龙胆目 Gentianales | 茜草科 Rubiaceae | 舒榄属 Bertiera | B. borbonica | Pailler et al, |
| 弯管花属 Chassalia | C. corallioides | Pailler et al, | ||
| 牛眼棠属 Cordiera | C. concolor | Judkevich et al, | ||
| 靛榄属 Genipa | 靛榄 G. americana | Judkevich et al, | ||
| 蓝茜树属 Randia | R. calycina, R. heteromera | Judkevich et al, | ||
| 木巴戟属 Morinda | 须弥巴戟 M. villosa | Sugawara et al, | ||
| 玉叶金花属 Mussaenda | 短裂玉叶金花 M. breviloba, 尾裂玉叶金花 M. caudatiloba, 仁昌玉叶金花 M. chingii, 展枝玉叶金花 M. divaricata, 楠藤 M. erosa, 海南玉叶金花 M. hainanensis, 粗毛玉叶金花 M. hirsutula, 红毛玉叶金花 M. hossei, 广西玉叶金花 M. kwangsiensis, 广东玉叶金花 M. kwangtungensis, 疏花玉叶金花 M. laxiflora, 长瓣玉叶金花 M. longipetala, 多毛玉叶金花 M. mollissima, 小玉叶金花 M. parviflora, M. pilosissima, 屏边玉叶金花 M. pingbianensis, 玉叶金花 M. pubescens, 白花玉叶金花 M. pubescens var. alba, 贡山玉叶金花 M. treutleri, 玉叶金花属5种 Mussaenda sp. 1-5 | 邓小芳, | ||
| 沙九节属 Pagamea | P. duckei | Terra-Araujo et al, | ||
| 九节属 Psychotria | 九节 P. rubra | Watanabe et al, | ||
| 马钱科 Loganiaceae | 髯管花属 Geniostoma | G. borbonica | Humeau et al, | |
| 顶髯管花属 Labordia | L. cyrtandrae, L. degeneri, L. fagraeoidea, L. hedyosmifolia, L. hirtella, L. hosakana, L. lydgatei, L. pumila, L. sessilis, L.venosa, L. waialealae, L. waiolani | Motley & Carr, | ||
| 夹竹桃科 Apocynaceae | 萝芙木属 Rauvolfia | R. sellowii | Souto & Koch, | |
| 唇形目 Lamiales | 马鞭草科 Verbenaceae | 琴木属 Citharexylum | C. myrianthum | Rocca & Sazima, |
| 唇形科 Lamiaceae | 紫珠属 Callicarpa | C. glabra, C. nishimurae, C. subpubescens | Kawakubo, | |
| 茄目 Solanales | 茄科 Solanaceae | 山盏茄属 Deprea | D. paneroi | Sawyer & Anderson, |
| 巫灵木属 Dunalia | D. solanacea | Hepper, | ||
| 枸杞属 Lycium | L. arenicolum, L. horridum, L. tetrandrum, L. villosum | Spies et al, | ||
| 茄目 Solanales | 茄科 Solanaceae | 茄属 Solanum | S. appendiculatum, S. campanulatum, S. confertiseriatum, S. crotonoides, S. conocarpum | Knapp et al, Anderson et al, |
| 串钟花属 Symonanthus | S. aromaticus, S. bancroftii | Haegi, | ||
| 睡茄属 Withania | W. aristata, W. coagulans | Sawyer & Anderson, | ||
| 性表达类型III Sex expression type III | ||||
| 金虎尾目 Malpighiales | 金虎尾科Malpighiaceae | 异金英属 Spachea | S. membranacea | Steiner, |
| 石竹目 Caryophyllales | 石竹科Caryophyllaceae | 洋漆姑属 Schiedea | S. adamantis | Mayer & Charlesworth, |
| 仙人掌科 Cactaceae | 圆柱掌属 Cylindropuntia | C. wolfii | Ramadoss et al, | |
| 裸萼球属 Gymnocalycium | 罗星丸 G. bruchii | Delbón et al, | ||
| 杜鹃花目 Ericales | 山榄科 Sapotaceae | 星苹果属 Chrysophyllum | C. gonocarpum | Judkevich et al, |
| 杜鹃花科 Ericaceae | 岩梨属 Epigaea | 岩梨 E. repens | Clay & Ellstrand, | |
| 性表达类型IV Sex expression type IV (1) (2) | ||||
| 菊目 Asterales | 菊科 Asteraceae | 蜂斗菜属 Petasites | (1)蜂斗菜 P. japonicas; (2)毛裂蜂斗菜 P. tricholobus | Sakai et al, |

图1 隐性雌雄异株2种常见形态学性表达类型的花部特征。(A)以葛枣猕猴桃为例示性表达类型I (形态上雄花两性花异株); (B)以玉叶金花为例示性表达类型II (形态上两性花植株)。蓝色箭头和红色箭头所指分别为可育性器官和不育性器官。
Fig. 1 Floral traits of two common sex expression types of cryptic dioecious species. (A) Actinidia polygama, indicating sex expression type I (morphologically androdioecy). (B) Mussaenda pubescens, indicating sex expression type II (morphologically hermaphrodite). The blue and red arrows indicate fertile and sterile sex organ, respectively.
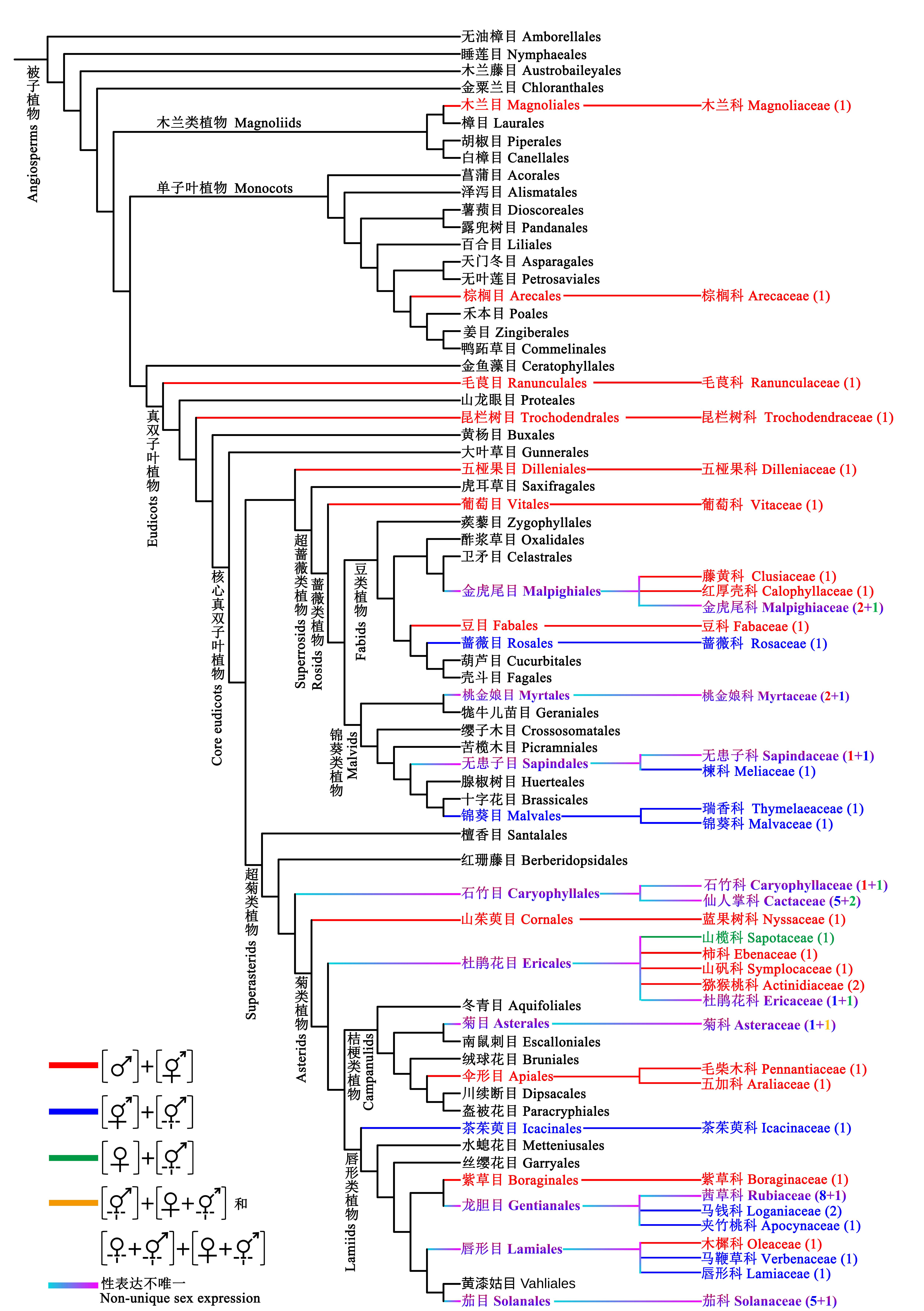
图2 具隐性雌雄异株性系统的分类群在被子植物中的系统分布。系统发育树参照APG IV (2016)和Li等(2019), 括号中数字表示具隐性雌雄异株性系统物种的属数。支长不代表真实时间尺度, 仅为美观考虑。♂表示雄花, ♀表示雌花, 表示两性花, 虚线表示该性别无功能, [ ]表示植株个体。
Fig. 2 The systematic distribution of taxa with cryptic dioecy species across angiosperm. Phylogenetic diagram adapted from APG IV (2016) and Li et al (2019). The value in parentheses represents the number of genera with cryptic dioecy species. Clade lengths are not indicative of true time scales and are for aesthetic purposes only. ♂ indicates male flowers, ♀ indicates female flowers, indicates hermaphrodite flowers, dashed line denotes a nonfunctional sex, [ ] indicates individuals.
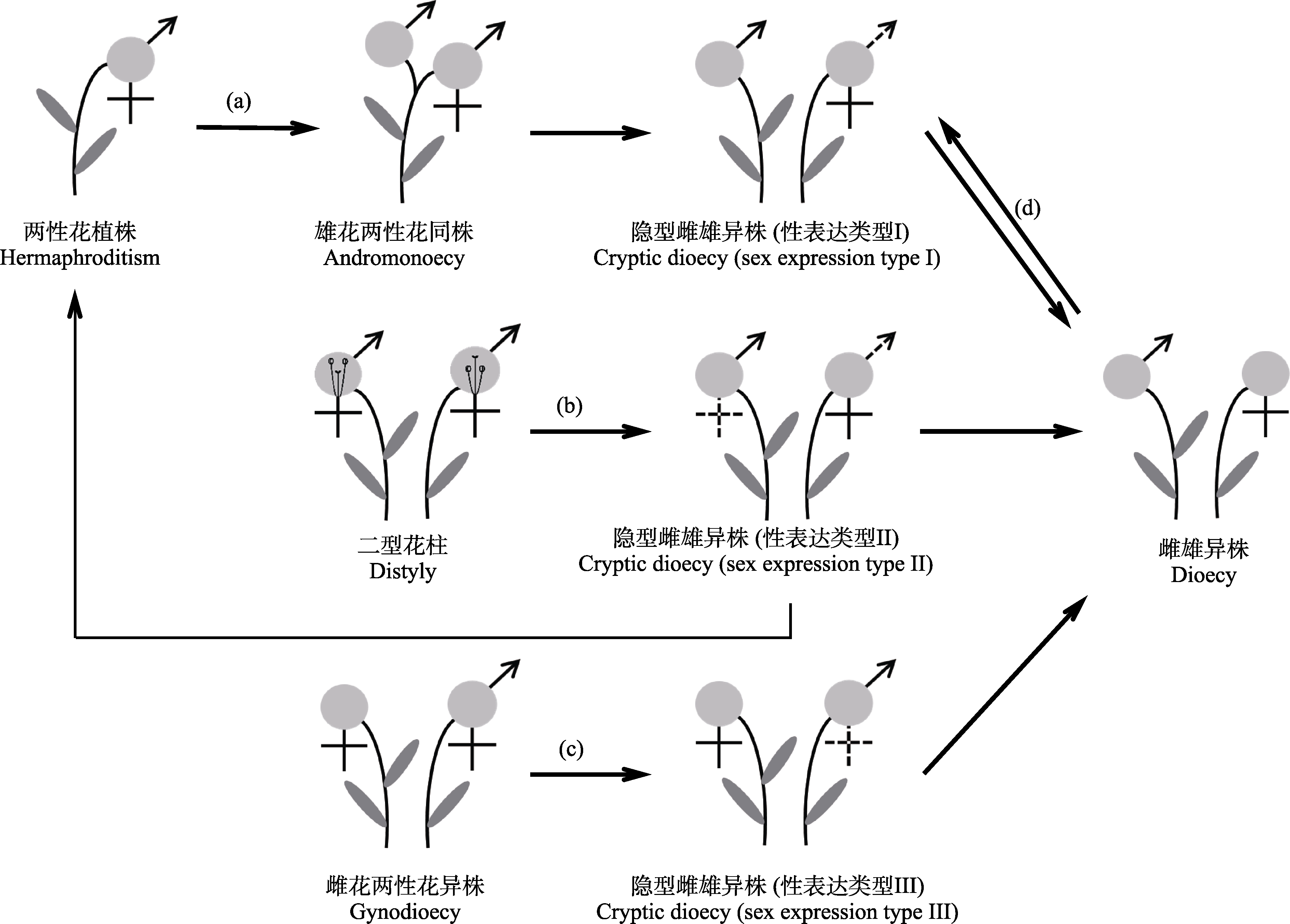
图3 被子植物隐性雌雄异株可能的主要进化途径。(a)经雄花两性花同株进化为隐性雌雄异株性表达类型I; (b)由二型花柱直接进化为隐性雌雄异株性表达类型II; (c)由雌花两性花异株直接进化为隐性雌雄异株性表达类型III; (d)由雌雄异株直接进化为隐性雌雄异株性表达类型I。花及其性别用浅灰色圆圈表示。
Fig. 3 Evolutionary pathways to cryptic dioecy in angiosperms. (a) Cryptic dioecy sex expression type I evolved via andromonoecy; (b) Cryptic dioecy sex expression type II evolved directly from distyly; (c) Cryptic dioecy sex expression type III evolved directly from gynodioecy; (d) Cryptic dioecy sex expression type Ⅰ evolved directly from dioecy. Flowers and their gender are depicted as light grey circles.
| [1] | Agren J, Schemske DW (1991) Pollination by deceit in a Neotropical monoecious herb, Begonia involucrata. Biotropica, 23, 235-241. |
| [2] |
Akagi T, Henry IM, Ohtani H, Morimoto T, Beppu K, Kataoka I, Tao R (2018) A Y-encoded suppressor of feminization arose via lineage-specific duplication of a cytokinin response regulator in kiwifruit. The Plant Cell, 30, 780-795.
DOI PMID |
| [3] | Akagi T, Henry IM, Tao R, Comai L (2014) A Y-chromosome- encoded small RNA acts as a sex determinant in persimmons. Science, 346, 646-650. |
| [4] | Anderson EF (2001) The Cactus Family. Timber Press, Portland. |
| [5] |
Anderson GJ, Anderson MK, Patel N (2015) The ecology, evolution, and biogeography of dioecy in the genus Solanum: With paradigms from the strong dioecy in Solanum polygamum, to the unsuspected and cryptic dioecy in Solanum conocarpum. American Journal of Botany, 102, 471-486.
DOI PMID |
| [6] | Anderson GJ, Bernardello G, Lopez P, Stuessy TF, Crawford DJ (2000) Dioecy and wind pollination in Pernettya rigida (Ericaceae) of the Juan Fernández Islands. Botanical Journal of the Linnean Society, 132, 121-141. |
| [7] |
Anderson GJ, Bernardello G, Opel MR, Santos-Guerra A, Anderson M (2006) Reproductive biology of the dioecious Canary Islands endemic Withania aristata (Solanaceae). American Journal of Botany, 93, 1295-1305.
DOI PMID |
| [8] |
Anderson GJ, Symon DE (1989) Functional dioecy and andromonoecy in Solanum. Evolution, 43, 204-219.
DOI PMID |
| [9] | Anderson WR (2001) Observations on the Malagasy genus Rhynchophora (Malpighiaceae). Contributions from the University of Michigan Herbarium, 23, 53-58. |
| [10] | APG Ⅳ (2016) An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Botanical Journal of the Linnean Society, 181, 1-20. |
| [11] | Appanah S (1982) Pollination of androdioecious Xerospermum intermedium Radlk. (Sapindaceae) in a rain forest. Biological Journal of the Linnean Society, 18, 11-34. |
| [12] | Aranha Filho JLM, Fritsch PW, Almeda F, Martins AB (2009) Cryptic dioecy is widespread in South American species of Symplocos section Barberina (Symplocaceae). Plant Systematics and Evolution, 277, 99-104. |
| [13] | Aranha Filho JLM, Fritsch PW, Almeda F, Martins AB (2013) Three new species of Symplocos sect. Hopea (Symplocaceae) from Brazil. Kew Bulletin, 68, 625-633. |
| [14] | Areces-Mallea AE (1992) Pereskia marcanoi, a new species of Cactaceae from Hispaniola. Brittonia, 44, 423-428. |
| [15] | Atlan A, Gouyon PH, Fournial T, Pomente D, Couvet D (1992) Sex allocation in an hermaphroditic plant: The case of gynodioecy in Thymus vulgaris L. Journal of Evolutionary Biology, 5, 189-203. |
| [16] | Baider C, Florens FBV (2016) A new and critically endangered species of Turraea (Meliaceae) endemic to the island of Mauritius. Phytotaxa, 247, 219-228. |
| [17] | Barrett SCH (2002a) Sexual interference of the floral kind. Heredity, 88, 154-159. |
| [18] | Barrett SCH (2002b) The evolution of plant sexual diversity. Nature Reviews Genetics, 3, 274-284. |
| [19] | Barrett SCH (2013) The evolution of plant reproductive systems:How often are transitions irreversible? Proceedings of the Royal Society B: Biological Sciences, 280, 20130913. |
| [20] | Batra SWT (1985) Red maple (Acer rubrum L.), an important early spring food resource for honey bees and other insects. Journal of the Kansas Entomological Society, 58, 169-172. |
| [21] | Bawa KS (1980a) Evolution of dioecy in flowering plants. Annual Review of Ecology and Systematics, 11, 15-39. |
| [22] | Bawa KS (1980b) Mimicry of male by female flowers and intrasexual competition for pollinators in Jacaratia dolichaula (D. Smith) Woodson (Caricaceae). Evolution, 34, 467-474. |
| [23] | Baylis GT (1977) Pennantia baylisiana (Oliver) Baylis comb. nov. New Zealand Journal of Botany, 15, 511-512. |
| [24] | Boyd FAH, Benkeblia N (2011) A description of the floral biology operative within Pimenta dioica (L.) Merr. Acta Horticulturae, 894, 83-88. |
| [25] | Brantjes NBM (1978) Pollinator attraction of Vitis vinifera subsp. silvestris. Vitis, 17, 229-233. |
| [26] | Callejas-Chavero A, Vargas-Mendoza CF, Gomez-Hinostrosa C, Arriola-Padilla VJ, Cornejo-Romero A (2021) Breeding system in a population of the globose cactus Mammillaria magnimamma at Valle del Mezquital, Mexico. Botanical Sciences, 99, 229-241. |
| [27] | Cane J (1993) Reproductive role of sterile pollen in cryptically dioecious species of flowering plants. Pollination Biology in Tropics, 65, 223-225. |
| [28] | Cerino MC, Castro DC, Richard GA, Exner E, Pensiero JF (2018) Functional dioecy in Gleditsia amorphoides (Fabaceae). Australian Journal of Botany, 66, 85-93. |
| [29] | Charlesworth B, Charlesworth D (1978) A model for the evolution of dioecy and gynodioecy. The American Naturalist, 112, 975-997. |
| [30] | Chen S (2013) The Evolutionary Breakdown of Distyly in Mussaenda (Rubiaceae). PhD dissertation, South China Botanical Garden, Chinese Academy of Sciences, Guangzhou, (in Chinese with English abstract) |
| [陈湜 (2013) 玉叶金花属(茜草科)二型花柱系统破损的进化途径. 博士学位论文. 中国科学院华南植物园, 广州.] | |
| [31] | Clark CJ, Lintas C (1992) Chemical composition of pollen from kiwifruit vines. New Zealand Journal of Crop and Horticultural Science, 20, 337-344. |
| [32] | Clay K, Ellstrand NC (1981) Stylar polymorphism in Epigaea repens, a dioecious species. Bulletin of the Torrey Botanical Club, 108, 305-310. |
| [33] | Cox PA (1982) Vertebrate pollination and the maintenance of dioecism in Freycinetia. The American Naturalist, 120, 65-80. |
| [34] | Davis CC (2002) Madagasikaria (Malpighiaceae): A new genus from Madagascar with implications for floral evolution in Malpighiaceae. American Journal of Botany, 89, 699-706. |
| [35] | Davis SL (1997) Stamens are not essential as an attractant for pollinators in females of cryptically dioecious Thalictrum pubescens Pursch. (Ranunculaceae). Sexual Plant Reproduction, 10, 293-299. |
| [36] | Davis SL (2002) Allocation to floral structures in Thalictrum pubescens (Ranunculaceae), a cryptically dioecious species. Annals of Botany, 90, 119-126. |
| [37] | Delbón NE, Aliscioni NL, Lorenzati M, García S, Singer RB, Gurvich DE (2024) Looking for non-hermaphrodite cacti: Multidisciplinary studies in Gymnocalycium bruchii endemic to central Argentina. Plant Reproduction, 37, 201-214. |
| [38] | Delph LF (2009) Sex allocation: Evolution to and from dioecy. Current Biology, 19, R249-R251. |
| [39] | Deng XF (2007) Taxonomic Revision and Phylogenetic Analysis of Breeding System Evolution of Mussaenda L. (Rubiaceae) in China. PhD dissertation, South China Botanical Garden, Chinese Academy of Sciences, Guangzhou. (in Chinese with English abstract) |
| [邓小芳 (2007) 中国玉叶金花属(茜草科)分类修订及繁育系统进化的研究. 博士学位论文. 中国科学院华南植物园, 广州.] | |
| [40] | Duan TT (2016) Evolutionary Pathways of Sexual Systems in Mussaenda and Phylogeography of M. pubescens. PhD dissertation, South China Botanical Garden, Chinese Academy of Sciences, Guangzhou. (in Chinese with English abstract) |
| [段婷婷 (2016) 玉叶金花属性系统进化及玉叶金花谱系地理学研究. 博士学位论文. 中国科学院华南植物园, 广州.] | |
| [41] | Dunthorn M (2004) Cryptic dioecy in Mammea (Clusiaceae). Plant Systematics and Evolution, 249, 191-196. |
| [42] | Fleming TH (1989) Climb every cactus-BCI’s Sonoran Desert bat-cactus project concludes its first field season in Mexico. Bats, 7, 3-6. |
| [43] | Haegi L (1981) A conspectus of Solanaceae tribe Anthocercideae. Telopea, 2, 173-180. |
| [44] | Hepper FN (1991) Old world Withania (Solanaceae): A taxonomic review and key to the species. In: Solanaceae Ⅲ: Taxonomy, Chemistry, Evolution (eds Hakes JG, Lester RN, Nee M, Estrada N), pp. 211-228. Royal Botanic Gardens, Kew, Richmond. |
| [45] | Hernández-Cruz R, Barrón-Pacheco F, Sánchez D, Arias S, Vázquez-Santana S (2018) Functional dioecy in Echinocereus: Ontogenetic patterns, programmed cell death, and evolutionary significance. International Journal of Plant Sciences, 179, 257-274. |
| [46] |
Hernández-Cruz R, Silva-Martínez J, García-Campusano F, Cruz-García F, Orozco-Arroyo G, Alfaro I, Vázquez- Santana S (2019) Comparative development of staminate and pistillate flowers in the dioecious cactus Opuntia robusta. Plant Reproduction, 32, 257-273.
DOI PMID |
| [47] | Holmes WC (1991) Dioecy in Mikania (Compositae: Eupatorieae). Plant Systematics and Evolution, 175, 87-92. |
| [48] |
Humeau L, Pailler T, Thompson JD (1999) Cryptic dioecy and leaky dioecy in endemic species of Dombeya (Sterculiaceae) on La Réunion. American Journal of Botany, 86, 1437-1447.
PMID |
| [49] | Humeau L, Pailler T, Thompson JD (2000) Variation in gender and flower-size dimorphism in the dioecious tree Dombeya ciliata, an endemic to La Réunion Island. Biotropica, 32, 463-472. |
| [50] | Humeau L, Strasberg D, Pailler T (2003) Dioécie cryptique chez Geniostoma borbonica, espèce pionnière endémique de La Réunion. Canadian Journal of Botany, 81, 897-904. |
| [51] |
Judkevich MD, Luaces PA, Gonzalez AM (2023) Flower structure, anatomy, and sexuality of Chrysophyllum gonocarpum (Sapotaceae). Protoplasma, 260, 1271-1285.
DOI PMID |
| [52] | Judkevich MD, Salas RM, Gonzalez AM (2022) Anther structure and pollen development in species of Rubiaceae and anatomical evidence of pathway to morphological dioecy. Anais da Academia Brasileira de Ciências, 94, e20191362. |
| [53] |
Käfer J, Marais GAB, Pannell JR (2017) On the rarity of dioecy in flowering plants. Molecular Ecology, 26, 1225-1241.
DOI PMID |
| [54] | Kawagoe T, Suzuki N (2004) Cryptic dioecy in Actinidia polygama: A test of the pollinator attraction hypothesis. Canadian Journal of Botany, 82, 214-218. |
| [55] | Kawakubo N (1998) Evolution of cryptic dioecy in Callicarpa (Verbenaceae) on the Bonin Islands. In: Evolution and Speciation of Island Plants (eds Stuessy T, Ono M), pp. 155-168. Cambridge University Press, Cambridge. |
| [56] | Keng H (1959) Androdioecism in the flowers of Trochodendron aralioides. Journal of the Arnold Arboretum, 40, 158-160. |
| [57] | Kevan PG, Blades DCA, Posluszny U, Ambrose JD (1988) Pollen dimorphism and dioecy in Vitis aestivalis. Vitis, 27, 143-146. |
| [58] | Kevan PG, Eisikowitch D, Ambrose JD, Kemp JR (1990) Cryptic dioecy and insect pollination in Rosa setigera Michx. (Rosaceae), a rare plant of Carolinian Canada. Biological Journal of the Linnean Society, 40, 229-243. |
| [59] | Kevan PG, Lack AJ (1985) Pollination in a cryptically dioecious plant Decaspermum parviflorum (Lam.) A. J. Scott (Myrtaceae) by pollen-collecting bees in Sulawesi, Indonesia. Biological Journal of the Linnean Society, 25, 319-330. |
| [60] | Kevan PG, Longair RW, Gadawski RM (1985) Dioecy and pollen dimorphism in Vitis riparia (Vitaceae). Canadian Journal of Botany, 63, 2263-2267. |
| [61] | Knapp S, Persson V, Blackmore S (1998) Pollen morphology and functional dioecy in Solanum (Solanaceae). Plant Systematics and Evolution, 210, 113-139. |
| [62] | Kubitzki K, Baretta-Kuipers T (1969) Pollendimorphie und androdiözie bei Tetracera (Dilleniaceae). Naturwissenschaften, 56, 219-220. |
| [63] | Le Péchon T, Pause JB, Dubuisson JY, Gigord L, Haevermans A, Haevermans T, Humeau L (2013) Dombeya formosa (Malvaceae s.l.): A new species endemic to La Réunion (Indian Ocean) based on morphological and molecular evidence. Systematic Botany, 38, 424-433. |
| [64] | Leal DO, Malafaia C, Cesar R, Pimentel RR, Santiago- Fernandes LDR, Lima HA, Sá-Haiad B (2012) Floral structure of Garcinia brasiliensis in relation to flower biology and evolution. International Journal of Plant Sciences, 173, 172-183. |
| [65] | Li AM, Wu XQ, Zhang DX, Barrett SCH (2010) Cryptic dioecy in Mussaenda pubescens (Rubiaceae): A species with stigma-height dimorphism. Annals of Botany, 106, 521-531. |
| [66] | Li HT, Yi TS, Gao LM, Ma PF, Zhang T, Yang JB, Gitzendanner MA, Fritsch PW, Cai J, Luo Y, Wang H, van der Bank M, Zhang SD, Wang QF, Wang J, Zhang ZR, Fu CN, Yang J, Hollingsworth PM, Chase MW, Soltis DE, Soltis PS, Li DZ (2019) Origin of angiosperms and the puzzle of the Jurassic gap. Nature Plants, 5, 461-470. |
| [67] | Liu Y, Luo ZL, Wu XQ, Bai XF, Zhang DX (2012) Functional dioecy in Morinda parvifolia (Rubiaceae), a species with stigma-height dimorphism. Plant Systematics and Evolution, 298, 775-785. |
| [68] | Lloyd DG (1979) Parental strategies of angiosperms. New Zealand Journal of Botany, 17, 595-606. |
| [69] | Lu J, Chen YN, Yin TM (2021) Research progress on sex determination genes of woody plants. Chinese Bulletin of Botany, 56, 90-103. (in Chinese with English abstract) |
| [陆静, 陈赢男, 尹佟明 (2021) 木本植物性别决定基因研究进展. 植物学报, 56, 90-103.] | |
| [70] | Martine CT, Anderson GJ, Les DH (2009) Gender-bending aubergines: Molecular phylogenetics of cryptically dioecious Solanum in Australia. Australian Systematic Botany, 22, 107-120. |
| [71] | Martine CT, Symon DE, Evans EC (2013) A new cryptically dioecious species of bush tomato (Solanum) from the Northern Territory, Australia. PhytoKeys, 30, 23-32. |
| [72] | Martine CT, Vanderpool D, Anderson GJ, Les DH (2006) Phylogenetic relationships of andromonoecious and dioecious Australian species of Solanum subgenus Leptostemonum section Melongena: Inferences from ITS sequence data. Systematic Botany, 31, 410-420. |
| [73] | Mayer SS, Charlesworth D (1991) Cryptic dioecy in flowering plants. Trends in Ecology & Evolution, 6, 320-325. |
| [74] |
Meagher TR (1992) The quantitative genetics of sexual dimorphism in Silene latifolia (Caryophyllaceae). I. Genetic variation. Evolution, 46, 445-457.
DOI PMID |
| [75] |
Melampy MN, Hayworth AM (1980) Seed production and pollen vectors in several nectarless plants. Evolution, 34, 1144-1154.
DOI PMID |
| [76] |
Motley TJ, Carr GD (1998) Artificial hybridization in the Hawaiian endemic genus Labordia (Loganiaceae). American Journal of Botany, 85, 654-660.
PMID |
| [77] |
Ndem-Galbert JR, Hall JE, McDonnell AJ, Martine CT (2021) Differential reward in “male” versus “female” pollen of functionally dioecious Solanum (Solanaceae). American Journal of Botany, 108, 2282-2293.
DOI PMID |
| [78] | Pailler T, Humeau L, Figier J, Thompson JD (1998a) Reproductive trait variation in the functionally dioecious and morphologically heterostylous island endemic Chassalia corallioides (Rubiaceae). Biological Journal of the Linnean Society, 64, 297-313. |
| [79] | Pailler T, Humeau L, Langlade F, Figier J (1998b) Cryptic dioecy in Bertiera borbonica var. borbonica (Rubiaceae), an endemic species of La Réunion Island. Acta Botanica Gallica, 145, 29-38. |
| [80] | Pätzold J, Feyrer B, Saumweber J, Hilger HH, Gottschling M (2023) Observations on flower and fruit anatomy in dioecious species of Cordia (Cordiaceae, Boraginales) with evolutionary interpretations. Organisms Diversity & Evolution, 23, 77-90. |
| [81] | Penny RH (2014) Sexual dimorphism in cryptically dioecious Thalictrum macrostylum. International Journal of Plant Sciences, 175, 794-802. |
| [82] | Penny RH (2015) Cryptic Dioecy and Evolution of Pollen Morphology in Thalictrum. PhD dissertation, Indiana University, Bloomington. |
| [83] | Penny RH (2018) Sterile stamens do not enhance seed set in females of the cryptically dioecious Thalictrum macrostylum (Ranunculaceae). Journal of the Torrey Botanical Society, 145, 82-90. |
| [84] | Ramadoss N, Orduño-Baez A, Portillo C, Steele S, Rebman J, Flores-Rentería L (2022) Unraveling the development behind unisexual flowers in Cylindropuntia wolfii (Cactaceae). BMC Plant Biology, 22, 94-108. |
| [85] | Rebman JP, Pinkava DJ (2001) Opuntia cacti of north America: An overview. Florida Entomologist, 84, 474-485. |
| [86] |
Renner SS (2014) The relative and absolute frequencies of angiosperm sexual systems: Dioecy, monoecy, gynodioecy, and an updated online database. American Journal of Botany, 101, 1588-1596.
DOI PMID |
| [87] | Renner SS, Ricklefs RE (1995) Dioecy and its correlates in the flowering plants. American Journal of Botany, 82, 596-606. |
| [88] | Rocca MA, Sazima M (2006) The dioecious, sphingophilous species Citharexylum myrianthum (Verbenaceae): Pollination and visitor diversity. Flora, 201, 440-450. |
| [89] | Sakai AK, Weller SG (1999) Gender and sexual dimorphism in flowering plants: A review of terminology, biogeographic patterns, ecological correlates, and phylogenetic approaches. In: Gender and Sexual Dimorphism in Flowering Plants (eds Geber MA, Dawson TE, Delph LF), pp.1-31. Springer, Berlin. |
| [90] |
Sakai AK, Weller SG, Chen ML, Chou SY, Tasanont C (1997) Evolution of gynodioecy and maintenance of females: The role of inbreeding depression, outcrossing rates, and resource allocation in Schiedea adamantis (Caryophyllaceae). Evolution, 51, 724-736.
DOI PMID |
| [91] | Sakai S, Suzuki Y, Itagaki T, Tsujisawa H, Makino TT (2008) On the function of hermaphrodite florets in female inflorescences of Petasites japonicus (Asteraceae). Botany, 86, 179-184. |
| [92] | Sawyer NW, Anderson GJ (2000) Dioecy in South American Deprea (Solanaceae). Biotropica, 32, 291-298. |
| [93] | Schlessman M, Lloyd D, Lowry PP (1990) Evolution of sexual systems in New Caledonian Araliaceae. Memoires of the New York Botanical Garden, 55, 105-117. |
| [94] | Seger J, Eckhart VM (1996) Evolution of sexual systems and sex allocation in plants when growth and reproduction overlap. Proceedings of the Royal Society of London B: Biological Sciences, 263, 833-841. |
| [95] | Sharma MV, Shaanker RU, Vasudeva R, Shivanna K (2010) Functional dioecy in Nothapodytes nimmoniana, a distylous species in the Western Ghats. Current Science, 99, 1444-1448. |
| [96] | Soejarto DD (1969) Aspects of reproduction in Saurauia. Journal of the Arnold Arboretum, 50, 180-196. |
| [97] | Souto LS, Koch I (2018) Explaining ovule sterility in Rauvolfia L. (Apocynaceae). International Journal of Plant Sciences, 179, 368-376. |
| [98] | Spies JJ, Minne L, Venter HJ, Venter AM (1993) A cytogenetic study of the functionally dioecious species in the genus Lycium (Solanaceae). South African Journal of Botany, 59, 535-540. |
| [99] | Stasiak A, Łata B, Bieniasz M, Latocha P (2020) Morphological variation of male A. arguta plants affects their flowering potential and pollen efficiency. Horticultural Science, 47, 100-109. |
| [100] | Steiner KE (1985) Functional dioecism in the Malpighiaceae: The breeding system of Spachea membranacea Cuatr. American Journal of Botany, 72, 1537-1543. |
| [101] | Strittmatter LI, Hickey RJ, Negrón-Ortiz V (2008) Heterochrony and its role in sex determination of cryptically dioecious Consolea (Cactaceae) staminate flowers. Botanical Journal of the Linnean Society, 156, 305-326. |
| [102] | Strittmatter LI, Negrón-Ortiz V, Hickey RJ (2006) Comparative microsporangium development in male-fertile and male-sterile flowers of Consolea (Cactaceae): When and how does pollen abortion occur. Grana, 45, 81-100. |
| [103] | Sugawara T, Tanaka N, Murata J (2011) Dioecy derived from distyly in Morinda villosa Hook. f. (Rubiaceae) occurring in Hukaung Valley, Kachin State, Myanmar. Journal of Japanese Botany, 86, 9-14. |
| [104] | Sun BL, Zhang CQ, Lowry PP, Wen J (2009) Cryptic dioecy in Nyssa yunnanensis (Nyssaceae), a critically endangered species from tropical eastern Asia. Annals of the Missouri Botanical Garden, 96, 672-684. |
| [105] | Terra-Araujo MH, Webber AC, Vicentini A (2012) Pollination of Pagamea duckei Standl. (Rubiaceae): A functionally dioecious species. Biota Neotropica, 12, 98-104. |
| [106] |
Timerman D, Barrett SCH (2020) Influence of local density and sex ratio on pollination in an ambophilous flowering plant. American Journal of Botany, 107, 587-598.
DOI PMID |
| [107] |
Torres MF, Mathew LS, Ahmed I, Al-Azwani IK, Krueger R, Rivera-Nuñez D, Mohamoud YA, Clark AG, Suhre K, Malek JA (2018) Genus-wide sequencing supports a two-locus model for sex-determination in Phoenix. Nature Communications, 9, 3969.
DOI PMID |
| [108] | Tsukui T, Sugawara T (1992) Dioecy in Honkenya peploides var. major (Caryophyllaceae). Journal of Plant Research, 105, 615-624. |
| [109] |
Vamosi JC, Otto SP, Barrett SCH (2003) Phylogenetic analysis of the ecological correlates of dioecy in angiosperms. Journal of Evolutionary Biology, 16, 1006-1018.
PMID |
| [110] | van Wyk AE, Dedekind I (1985) The genus Eugenia (Myrtaceae) in southern Africa: Morphology and taxonomic value of pollen. South African Journal of Botany, 51, 371-378. |
| [111] | van der Merwe MM, van Wyk AE, Botha AM (2005) Molecular phylogenetic analysis of Eugenia L. (Myrtaceae), with emphasis on southern African taxa. Plant Systematics and Evolution, 251, 21-34. |
| [112] |
Vernet P, Lepercq P, Billiard S, Bourceaux A, Lepart J, Dommée B, Saumitou-Laprade P (2016) Evidence for the long-term maintenance of a rare self-incompatibility system in Oleaceae. New Phytologist, 210, 1408-1417.
DOI PMID |
| [113] | Wang YC, Hu JM (2011) Cryptic dioecy of Symplocos wikstroemiifolia Hayata (Symplocaceae) in Taiwan. Botanical Studies, 52, 479-491. |
| [114] | Watanabe K, ShimizuA, Sugawara T (2013) Dioecy derived from distyly and pollination in Psychotria rubra (Rubiaceae) occurring in the Ryukyu Islands, Japan. Plant Species Biology, 29, 181-191. |
| [115] | Willmer P (2011) Pollination and Floral Ecology. Princeton University Press, Princeton. |
| [116] | Yu DP, Li CH, Wen XY, Li XJ, Peng QX, Xie KP (2019) Flowering biological characteristics and breeding system of Parakmeria omeiensis. Guihaia, 39, 600-607. (in Chinese with English abstract) |
| [余道平, 李策宏, 文香英, 李小杰, 彭启新, 谢孔平 (2019) 峨眉拟单性木兰的开花生物学特性与繁育系统. 广西植物, 39, 600-607.] | |
| [117] | Yu Q, Li DX, Luo W, Guo YH (2011) Function and evolution of sterile sex organs in cryptically dioecious Petasites tricholobus (Asteraceae). Annals of Botany, 108, 65-71. |
| [118] |
Yu Q, Zhang YW, Guo YH (2008) Translation and elucidation of common terms in pollination biology. Journal of Systematics and Evolution, 46, 96-102. (in Chinese with English abstract)
DOI |
| [予茜, 张彦文, 郭友好 (2008) 传粉生物学常用术语释译. 植物分类学报, 46, 96-102.] | |
| [119] | Zhang DY (2004) Plant Life-History Evolution and Reproductive Ecology. Science Press, Beijing. (in Chinese) |
| [张大勇 (2004) 植物生活史进化与繁殖生态学. 科学出版社, 北京.] |
| [1] | 王凤英, 吴增源, 崔涵, 李垠蕾, 邓莉娟, 王红, 刘杰. 第三极荨麻属麻叶荨麻分支的物种界限[J]. 生物多样性, 2025, 33(8): 25138-. |
| [2] | 徐凯伦, 陈小荣, 张敏华, 于婉婉, 吴素美, 朱志成, 陈定云, 兰荣光, 董舒, 刘宇. 演替和地形共同影响浙江百山祖森林群落的性系统多样性[J]. 生物多样性, 2024, 32(12): 24338-. |
| [3] | 张飞飞, 杨天凤, 陈莉荣, 刘冬梅, 杨柳园, 杨杜宇, 鞠鹏, 陆露. 被子植物花粉颜色多样性及应用研究进展[J]. 生物多样性, 2024, 32(1): 23346-. |
| [4] | 钱宏, 张健, 赵静超. 世界上已知维管植物有多少种? 基于多个全球植物数据库的整合[J]. 生物多样性, 2022, 30(7): 22254-. |
| [5] | 王芸芸, 郝占庆. 被子植物性系统的多样性、生态功能及分布规律[J]. 生物多样性, 2022, 30(7): 22065-. |
| [6] | 寄玲, 谢宜飞, 李中阳, 许廷晨, 杨波, 李波. 江西省野生维管植物名录[J]. 生物多样性, 2022, 30(6): 22057-. |
| [7] | 张淑梅, 李微, 李丁男. 辽宁省高等植物多样性编目[J]. 生物多样性, 2022, 30(6): 22038-. |
| [8] | 吴墨栩, 安明态, 田力, 刘锋. 茂兰喀斯特森林木本植物性系统数量特征及其与环境因子的关系[J]. 生物多样性, 2022, 30(11): 22025-. |
| [9] | 扈凡斌, 辛玥, 郭柯, 赵利清. 采自西藏和新疆的7种中国新记录植物[J]. 生物多样性, 2021, 29(9): 1265-1270. |
| [10] | 都业勤, 张迪, 王赛, 王磊, 闫兴富, 唐占辉. 湿地植物大花百合种群的性系统特征[J]. 生物多样性, 2021, 29(10): 1321-1335. |
| [11] | 孙远, 胡维刚, 姚树冉, 孙颖, 邓建明. 黄河流域被子植物和陆栖脊椎动物丰富度格局及其影响因子[J]. 生物多样性, 2020, 28(12): 1523-1532. |
| [12] | 葛美玲, 徐勤增, 范士亮, 王宗兴, 张学雷. 中国近海多毛纲底栖类群目与科水平的分类[J]. 生物多样性, 2018, 26(9): 998-1003. |
| [13] | 覃海宁, 赵莉娜, 于胜祥, 刘慧圆, 刘博, 夏念和, 彭华, 李振宇, 张志翔, 何兴金, 尹林克, 林余霖, 刘全儒, 侯元同, 刘演, 刘启新, 曹伟, 李建强, 陈世龙, 金效华, 高天刚, 陈文俐, 马海英, 耿玉英, 金孝锋, 常朝阳, 蒋宏, 蔡蕾, 臧春鑫, 武建勇, 叶建飞, 赖阳均, 刘冰, 林秦文, 薛纳新. 中国被子植物濒危等级的评估[J]. 生物多样性, 2017, 25(7): 745-757. |
| [14] | 王伟, 张晓霞, 陈之端, 路安民. 被子植物APG分类系统评论[J]. 生物多样性, 2017, 25(4): 418-426. |
| [15] | 王家坚, 彭智邦, 孙航, 聂泽龙, 孟盈. 青藏高原与横断山被子植物区系演化的 细胞地理学特征[J]. 生物多样性, 2017, 25(2): 218-225. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
备案号:京ICP备16067583号-7
Copyright © 2026 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn
![]()