
生物多样性 ›› 2018, Vol. 26 ›› Issue (9): 931-940. DOI: 10.17520/biods.2018059 cstr: 32101.14.biods.2018059
庄鸿飞1,2, 张殷波3, 王伟1,4,*( ), 任月恒1, 刘方正1, 杜金鸿1, 周越1
), 任月恒1, 刘方正1, 杜金鸿1, 周越1
收稿日期:2018-02-13
接受日期:2018-05-15
出版日期:2018-09-20
发布日期:2019-01-05
通讯作者:
王伟
作者简介:# 共同第一作者
基金资助:
Hongfei Zhuang1,2, Yinbo Zhang3, Wei Wang1,4,*(), Yueheng Ren1, Fangzheng Liu1, Jinhong Du1, Yue Zhou1
Received:2018-02-13
Accepted:2018-05-15
Online:2018-09-20
Published:2019-01-05
Contact:
Wang Wei
About author:# Co-first authors
摘要:
单一空间尺度构建的最大熵(maximum entropy, MaxEnt)模型是否具有代表性, 是MaxEnt模型应用与发展中面临的重要问题。本研究基于有效的地理分布位点数据, 利用最小凸多边形法(the minimum convex polygon method)在三江并流、云南省及全国3个空间尺度下分别识别了红色木莲(Manglietia insignis)的建模区域, 并进一步建立MaxEnt模型: 使用ROC曲线分析法与遗漏率(omission rate, OR)检验评估MaxEnt模型预测精度; 基于ArcGIS分析分布概率及其热点区域的分布趋势, 并通过分区统计工具Zonal识别潜在适宜分布区域的质心位置; 采用刀切法检验环境因子贡献率。结果表明: (1)不同尺度下红色木莲的MaxEnt模型都有良好的预测效果, 三江并流、云南省及全国尺度下的AUC值分别为0.936、0.887和0.930, OR值分别为0.18、0.15和0.20; (2)各尺度红色木莲的适生区格局呈现一致性分布趋势, 集中在独龙江、怒江和澜沧江3个流域; (3) 3个空间尺度下红色木莲的地理分布受不同环境因子影响, 存在着尺度依赖效应。由此可见, 红色木莲在不同空间尺度下的预测模型有着稳定的性能表现与良好的预测效果。此外, 我们建议在野外实地调查与野生生物资源保护中加强对普通物种的关注, 在预测物种地理分布的研究中将MaxEnt模型与热点分析结合使用。
庄鸿飞, 张殷波, 王伟, 任月恒, 刘方正, 杜金鸿, 周越 (2018) 基于最大熵模型的不同尺度物种分布概率优化热点分析: 以红色木莲为例. 生物多样性, 26, 931-940. DOI: 10.17520/biods.2018059.
Hongfei Zhuang, Yinbo Zhang, Wei Wang, Yueheng Ren, Fangzheng Liu, Jinhong Du, Yue Zhou (2018) Optimized hot spot analysis for probability of species distribution under different spatial scales based on MaxEnt model: Manglietia insignis case. Biodiversity Science, 26, 931-940. DOI: 10.17520/biods.2018059.
| 尺度 Scale | 覆盖范围 Cover range | 红色木莲分布现状 Distribution status of Manglietia insignis |
|---|---|---|
| 三江并流 Three Parallel Rivers | 泸水县、兰坪白族普米族自治县、玉龙纳西族自治县、福贡县、维西傈僳族自治县、德钦县、贡山独龙族怒族自治县和香格里拉市8个行 政县, 约4.5万km2 Including Lushui County, Lanping Bai Pumi Autonomous County, Yulong Naxi Autonomous County, Fugong County, Weixi County, Deqin County, Gongshan County, and Shangri-La City, about 45,000 km2 | 泸水县、福贡县、德钦县和贡山独龙族怒族自治县有其分布记录 Lushui County, Fugong County, Deqin County, and Gongshan County with distribution records |
| 云南省 Yunnan Province | 包含昆明市、玉溪市和迪庆藏族自治州等16个州市, 约39万km2 Containing 16 cities, such as Kunming, Yuxi, and Dêqên Tibetan Autonomous Prefecture, about 390,000 km2 | 临沧市、普洱市和玉溪市等12个州市有其分布 记录 Twelve cities including Lincang City, Puer City, and Yuxi with distribution records |
| 全国 Country | 包含省、自治区、直辖市及特别行政区在内的34个省级行政区域, 约 960万km2 Containing 34 provincial administrative regions, including provinces, autonomous regions, municipalities, and special administrative regions, about 9,600,000 km2 | 湖北、湖南、福建、广西、四川、贵州、云南和西藏8省有其分布记录 Hubei, Hunan, Fujian, Guangxi, Sichuan, Guizhou, Yunnan, Tibet with distribution records |
表1 研究区域的尺度设置与红色木莲的分布现状
Table 1 Scale setting of the study areas and the distribution status of Manglietia insignis
| 尺度 Scale | 覆盖范围 Cover range | 红色木莲分布现状 Distribution status of Manglietia insignis |
|---|---|---|
| 三江并流 Three Parallel Rivers | 泸水县、兰坪白族普米族自治县、玉龙纳西族自治县、福贡县、维西傈僳族自治县、德钦县、贡山独龙族怒族自治县和香格里拉市8个行 政县, 约4.5万km2 Including Lushui County, Lanping Bai Pumi Autonomous County, Yulong Naxi Autonomous County, Fugong County, Weixi County, Deqin County, Gongshan County, and Shangri-La City, about 45,000 km2 | 泸水县、福贡县、德钦县和贡山独龙族怒族自治县有其分布记录 Lushui County, Fugong County, Deqin County, and Gongshan County with distribution records |
| 云南省 Yunnan Province | 包含昆明市、玉溪市和迪庆藏族自治州等16个州市, 约39万km2 Containing 16 cities, such as Kunming, Yuxi, and Dêqên Tibetan Autonomous Prefecture, about 390,000 km2 | 临沧市、普洱市和玉溪市等12个州市有其分布 记录 Twelve cities including Lincang City, Puer City, and Yuxi with distribution records |
| 全国 Country | 包含省、自治区、直辖市及特别行政区在内的34个省级行政区域, 约 960万km2 Containing 34 provincial administrative regions, including provinces, autonomous regions, municipalities, and special administrative regions, about 9,600,000 km2 | 湖北、湖南、福建、广西、四川、贵州、云南和西藏8省有其分布记录 Hubei, Hunan, Fujian, Guangxi, Sichuan, Guizhou, Yunnan, Tibet with distribution records |
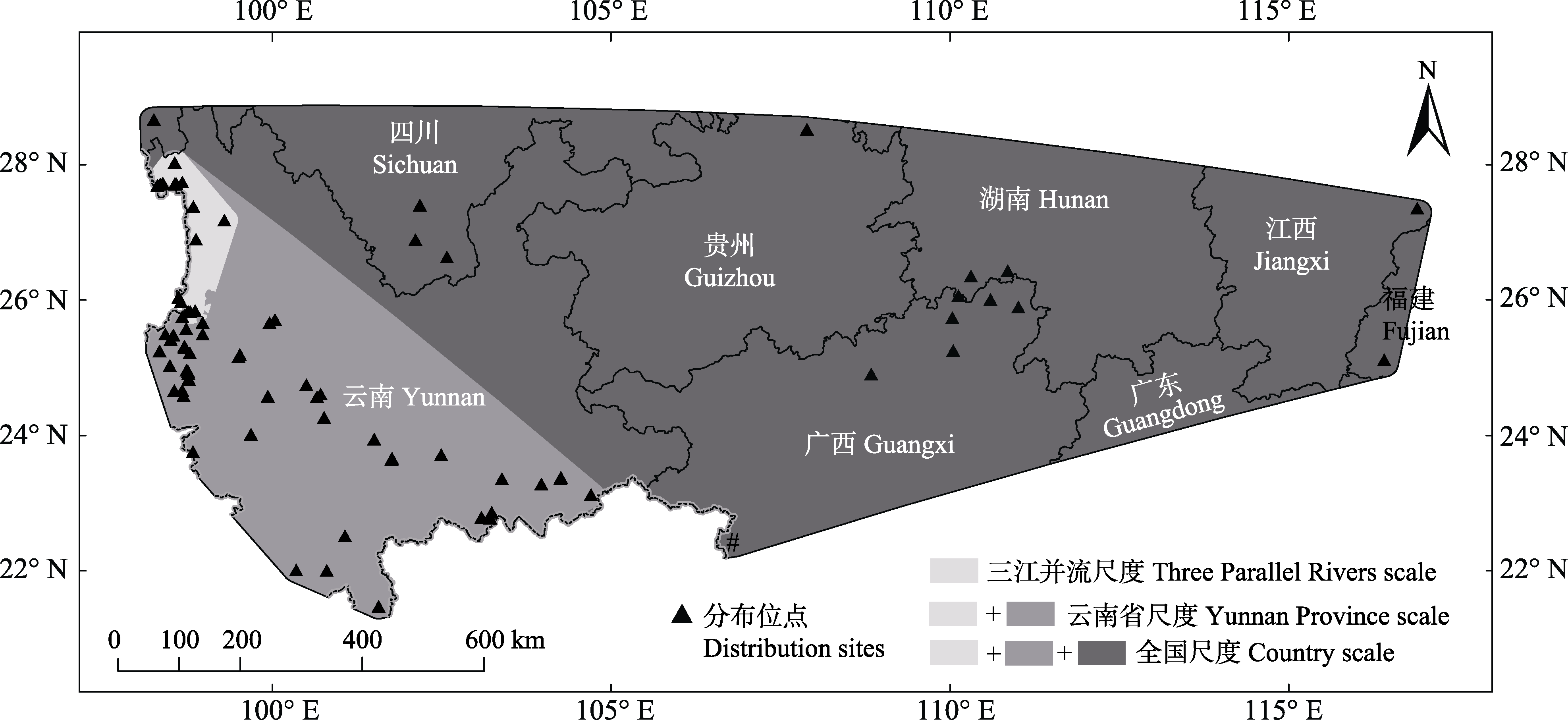
图1 3个空间尺度的建模区域与红色木莲分布位点
Fig. 1 Modeling areas of three spatial scales and distribution sites of Manglietia insignis
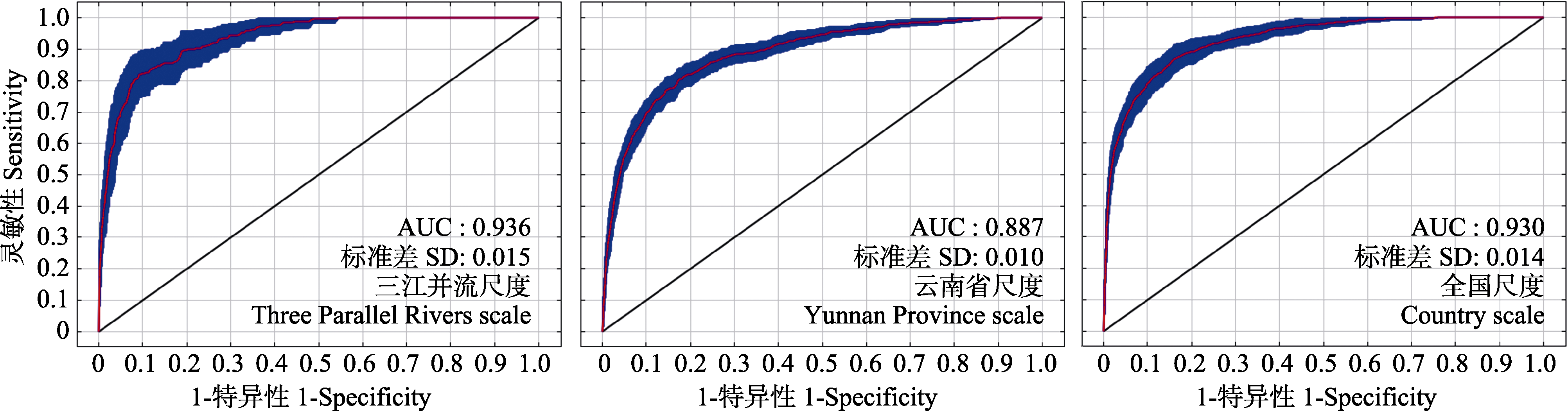
图2 3个空间尺度的受试者工作曲线与曲线下面积
Fig. 2 Receiver operating characteristic curve of three spatial scales and area under the curve (AUC)
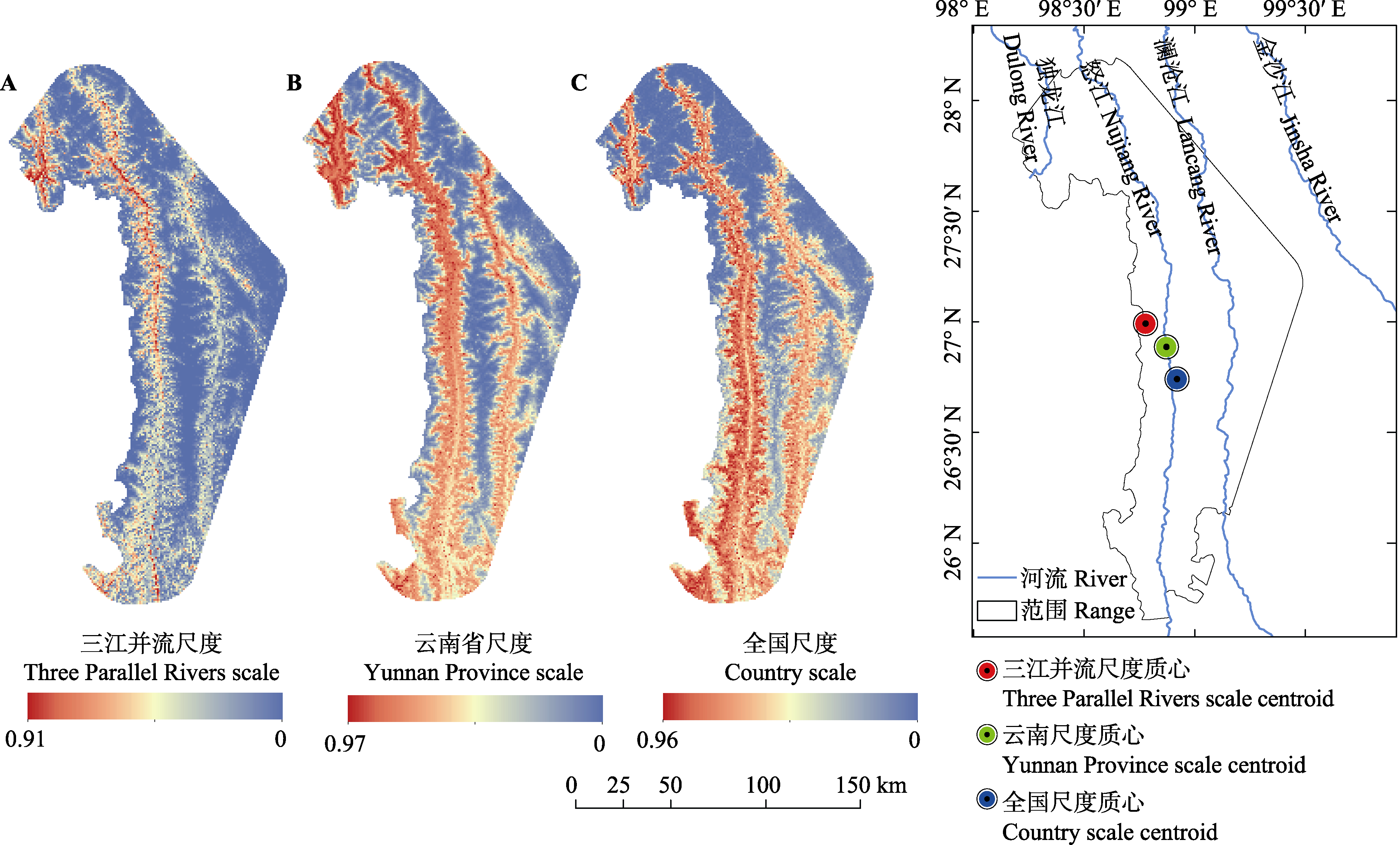
图3 3个空间尺度下红色木莲的潜在适宜性及其质心分布
Fig. 3 Potential suitable distributions and its centroids of Manglietia insignis under three spatial scales
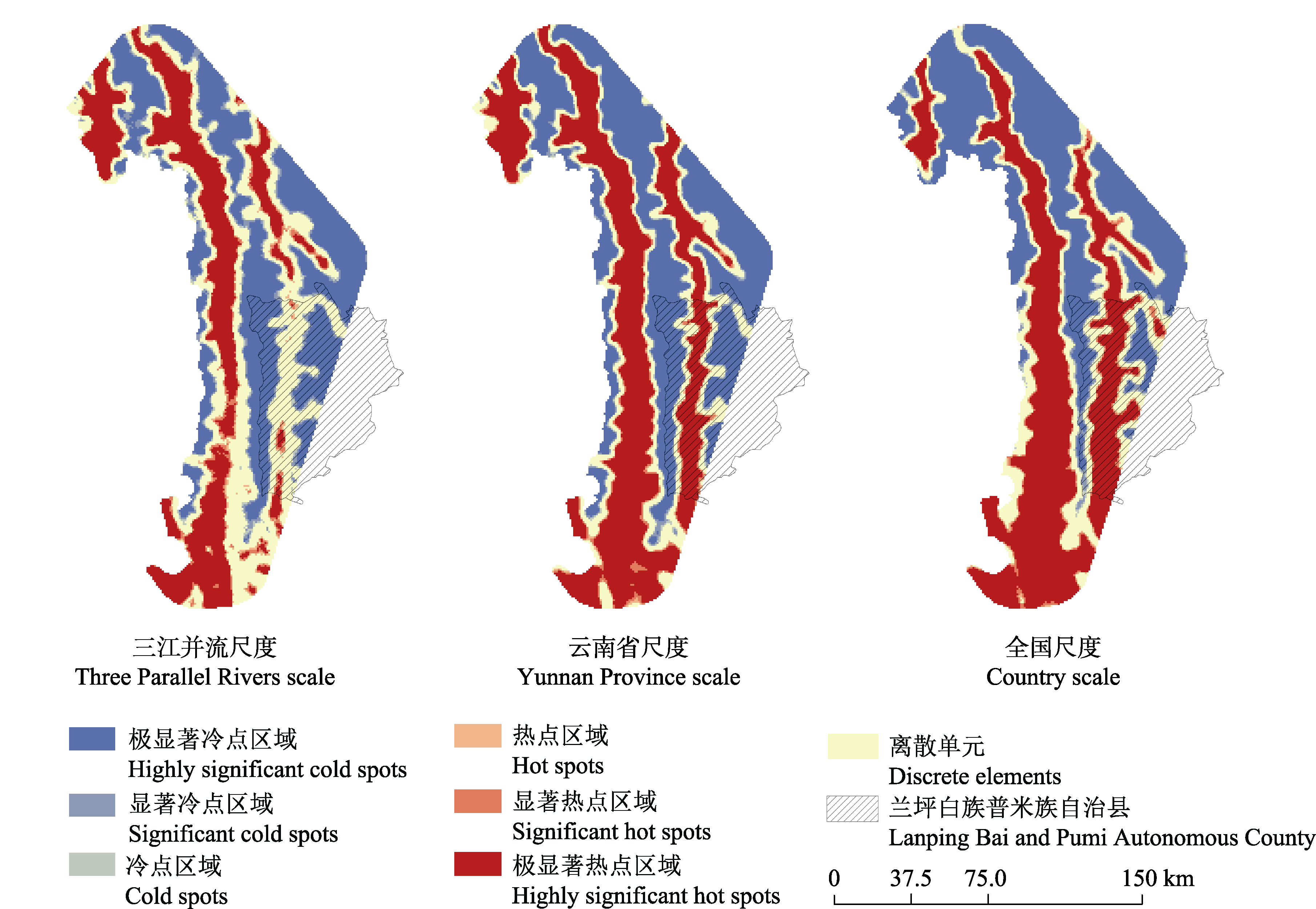
图4 3个空间尺度下红色木莲分布概率的优化热点分析
Fig. 4 Optimized hot spot analysis of probability distribution of Manglietia insignis under three spatial scales
| 尺度 Scale | 变量 Variable | 环境因子贡献率 Contribution (%) | 尺度 Scale | 变量 Variable | 环境因子贡献率 Contribution (%) |
|---|---|---|---|---|---|
| 三江并流尺度 TPRS | b1 | 3.4 | b15 | 13.7 | |
| b2 | 9.8 | alt | 2.9 | ||
| b4 | 6.2 | slo | 10.1 | ||
| b15 | 11 | asp | 3.5 | ||
| alt | 39.4* | 全国尺度 CS | b1 | 5.9 | |
| slo | 8.1 | b2 | 24.8* | ||
| asp | 22.1* | b7 | 21.7* | ||
| 云南省尺度 YPS | b1 | 5.1 | b12 | 11.4 | |
| b2 | 8.9 | b18 | 14.1 | ||
| b4 | 18.8 | alt | 7.1 | ||
| b12 | 35.5* | slo | 10.4 | ||
| b14 | 1.6 | asp | 4.7 |
表2 三江并流、云南省、全国3个尺度下红色木莲MaxEnt模型的环境因子贡献率
Table 2 Contribution of environmental variables for Manglietia insignis under Three Parallel Rivers scale (TPRS), Yunnan Province scale (YPS), and country scale (CS)
| 尺度 Scale | 变量 Variable | 环境因子贡献率 Contribution (%) | 尺度 Scale | 变量 Variable | 环境因子贡献率 Contribution (%) |
|---|---|---|---|---|---|
| 三江并流尺度 TPRS | b1 | 3.4 | b15 | 13.7 | |
| b2 | 9.8 | alt | 2.9 | ||
| b4 | 6.2 | slo | 10.1 | ||
| b15 | 11 | asp | 3.5 | ||
| alt | 39.4* | 全国尺度 CS | b1 | 5.9 | |
| slo | 8.1 | b2 | 24.8* | ||
| asp | 22.1* | b7 | 21.7* | ||
| 云南省尺度 YPS | b1 | 5.1 | b12 | 11.4 | |
| b2 | 8.9 | b18 | 14.1 | ||
| b4 | 18.8 | alt | 7.1 | ||
| b12 | 35.5* | slo | 10.4 | ||
| b14 | 1.6 | asp | 4.7 |
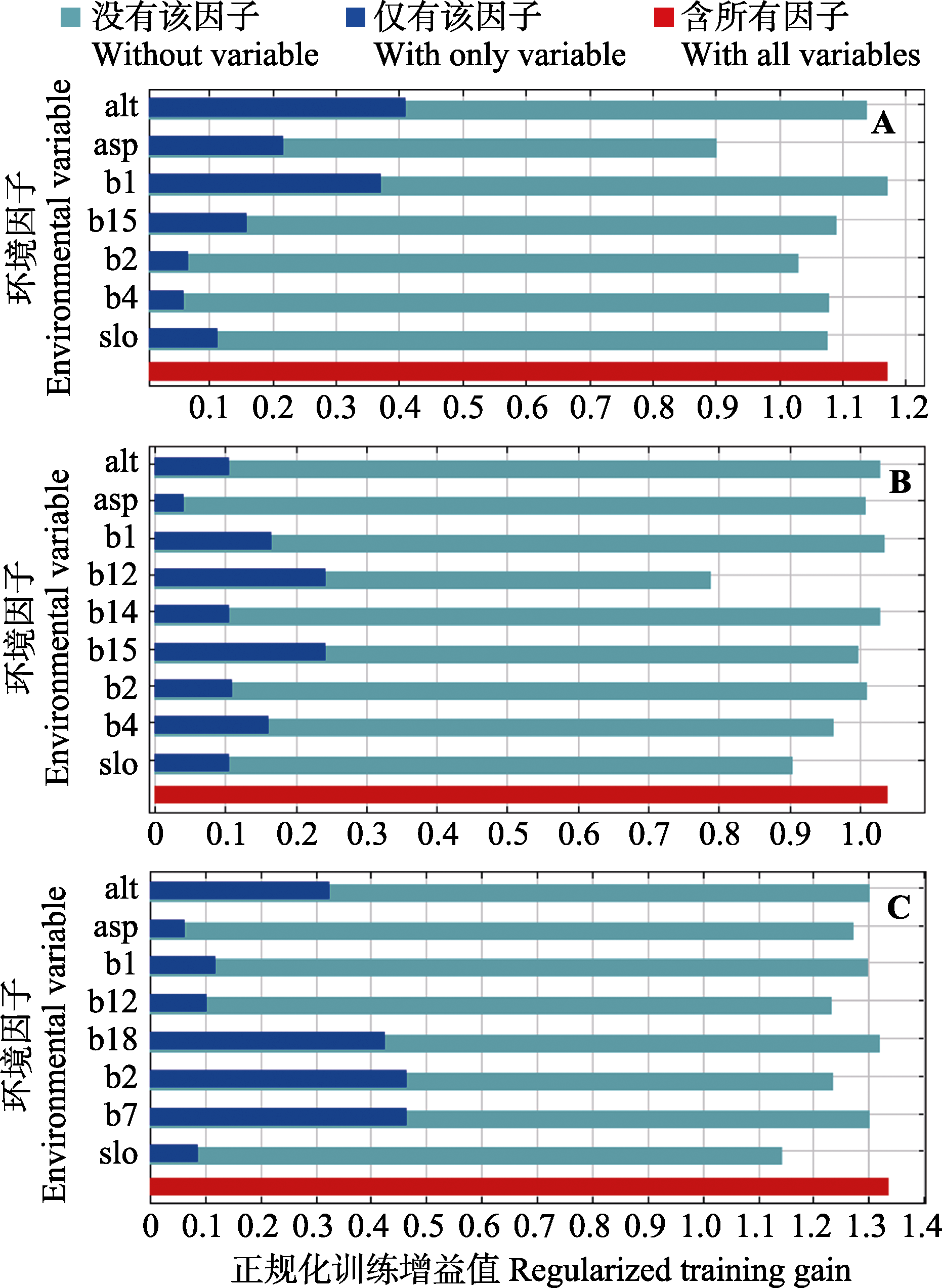
图5 三江并流尺度(A)、云南省尺度(B)与全国尺度(C)下环境因子正规化训练增益值的刀切法评估
Fig. 5 Results of Jackknife evaluation of the environmental variables with respect to regularized training gain for Three Parallel Rivers scale (A), Yunnan Province scale (B), and country scale (C)
| [1] | Ahmed SE, McInerny G, O’Hara K, Harper R, Salido L, Emmott S, Joppa LN (2015) Scientists and software—surveying the species distribution modelling community. Diversity and Distributions, 21, 258-267. |
| [2] | Barbosa FG, Schneck F (2015) Characteristics of the top-cited papers in species distribution predictive models. Ecological Modelling, 313, 77-83. |
| [3] | Catherineh G, Jane E, Robertj H, Antoine G, Townsend PA, Bettea L (2008) The influence of spatial errors in species occurrence data used in distribution models. Journal of Applied Ecology, 45, 239-247. |
| [4] | Chai Y, Zhu H, Meng GT, Shi JP, Yang GB (2011) Population structure and distribution pattern of dominant tree species in ancient tea tree community in Ailao Mountains of Yunnan Province, China. Forest Research, 24, 277-284. (in Chinese with English abstract) |
| [柴勇, 朱华, 孟广涛, 施济普, 杨国平 (2011) 云南哀牢山古茶树群落优势树种的种群结构与分布格局. 林业科学研究, 24, 277-284.] | |
| [5] | Chen F, Wang JM, Sun BG, Chen XM, Yang ZX, Duan ZY (2012) Relationships of plant species distribution in different strata of Pinus yunnanensis forest with landform and climatic factors. Chinese Journal of Ecology, 31, 1070-1076. (in Chinese with English abstract) |
| [陈飞, 王健敏, 孙宝刚, 陈晓鸣, 杨子祥, 段兆尧 (2012) 云南松林不同层植物分布与地形、气候因子的关系. 生态学杂志, 31, 1070-1076.] | |
| [6] | Chen XM, Lei YC, Zhang XQ, Jia HY (2012) Effects of sample sizes on accuracy and stability of maximum entropy model in predicting species distribution. Scientia Silvae Sinicae, 48(1), 53-59. (in Chinese with English abstract) |
| [陈新美, 雷渊才, 张雄清, 贾宏炎 (2012) 样本量对MaxEnt模型预测物种分布精度和稳定性的影响. 林业科学, 48(1), 53-59.] | |
| [7] | Dong XF (2017) Effect of topographic factors on the distribution of Manglietia insignis. Journal of Anhui Agricultural Sciences, 45(10), 162-163. (in Chinese with English abstract) |
| [董学芬 (2017) 地形因素对红花木莲分布的影响. 安徽农业科学, 45(10), 162-163.] | |
| [8] | Elith J, Phillips SJ, Hastie T, Dudík M, Chee YE, Yates CJ (2011) A statistical explanation of MaxEnt for ecologists. Diversity and Distributions, 17, 43-57. |
| [9] | Escalante T, Rodríguez-Tapia G, Linaje M, Illoldi-Rangel P, González-López R (2013) Identification of areas of endemism from species distribution models: Threshold selection and Nearctic mammals. Transaction Image Processing, 16, 5-17. |
| [10] | Getis A, Ord JK (1992) The analysis of spatial association by use of distance statistics. Geographical Analysis, 24, 189-206. |
| [11] | Hu X, Wu FC, Guo W, Liu N (2014) Identification of potential cultivation region for Santalum album in China by the MaxEnt ecologic niche model. Scientia Silvae Sinicae, 50(5), 27-33. (in Chinese with English abstract) |
| [胡秀, 吴福川, 郭微, 刘念 (2014) 基于MaxEnt生态学模型的檀香在中国的潜在种植区预测. 林业科学, 50(5), 27-33.] | |
| [12] | Jiang ZG (1996) Hoarding behavior of animal and its ecological functions. Chinese Journal of Zoology, (3), 47-49. (in Chinese) |
| [蒋志刚 (1996) 动物贮食行为及其生态意义. 动物学杂志, (3), 47-49.] | |
| [13] | Kunming Institute of Botany, Chinese Academy of Sciences(2006) Flora of Yunnan. Science Press, Beijing. (in Chinese) |
| [中国科学院昆明植物研究所(2006) 云南植物志. 科学出版社, 北京.] | |
| [14] | Levin SA (1992) The problem of pattern and scale in ecology: The Robert H. MacArthur Award lecture. Ecology, 73, 1943-1967. |
| [15] | Li HJ, Zhang ZB (2001) Relationship between animals and plant regeneration by seed. II. Seed predation, dispersal and burial by animals and relationship between animals and seedling establishment. Biodiversity Science, 9, 25-37. (in Chinese with English abstract) |
| [李宏俊, 张知彬 (2001) 动物与植物种子更新的关系. II. 动物对种子的捕食、扩散、贮藏及与幼苗建成的关系. 生物多样性, 9, 25-37.] | |
| [16] | Lin YP, Deng D, Lin WC, Lemmens R, Crossman ND, Henle K, Schmeller DS (2015) Uncertainty analysis of crowd- sourced and professionally collected field data used in species distribution models of Taiwanese moths. Biological Conservation, 181, 102-110. |
| [17] | Manhães AP, Loyola R, Mazzochini GG, Ganade G, Oliveira-Filho AT, Carvalho AR (2018) Low-cost strategies for protecting ecosystem services and biodiversity. Biological Conservation, 217, 187-194. |
| [18] | Morales NS, Fernández IC, Bacagonzález V (2017) MaxEnt’s parameter configuration and small samples: Are we paying attention to recommendations? A systematic review. PeerJ, 5, e3093. |
| [19] | Myers N (1990) The biodiversity challenge: Expanded hot-spots analysis. Environmentalist, 10, 243-256. |
| [20] | Nüchel J, Bøcher PK, Xiao W, Zhu AX, Svenning JC (2018) Snub-nosed monkeys (Rhinopithecus): Potential distribution and its implication for conservation. Biodiversity and Conservation, 27, 1517-1538. |
| [21] | Ord JK, Getis A (1995) Local spatial autocorrelation statistics: Distributional issues and an application. Geographical Analysis, 27, 286-306. |
| [22] | Phillips SJ, Anderson RP, Schapire RE (2006) Maximum entropy modeling of species geographic distributions. Ecological Modelling, 190, 231-259. |
| [23] | Phillips SJ, Dudík M, Schapire RE (2004) A Maximum Entropy Approach to Species Distribution Modeling. p. 83. Association for Computing Machinery, Banff. |
| [24] | Raes N, Roos MC, Slik JWF, van Loon EE, ter Steege H (2009) Botanical richness and endemicity patterns of Borneo derived from species distribution models. Ecography, 32, 180-192. |
| [25] | Raes N, Steege HT (2007) A null-model for significance testing of presence-only species distribution models. Ecography, 30, 727-736. |
| [26] | Scott JM, Csuti B, Jacobi JD, Estes JE (1987) Species richness. BioScience, 37, 782-788. |
| [27] | Sierra R, Campos F, Chamberlin J (2002) Assessing biodiversity conservation priorities: Ecosystem risk and representativeness in continental Ecuador. Landscape and Urban Planning, 59, 95-110. |
| [28] | Song W, Kim E, Lee D, Lee M, Jeon SW (2013) The sensitivity of species distribution modeling to scale differences. Ecological Modelling, 248, 113-118. |
| [29] | Tang JH, Cheng YX, Luo LZ, Zhang L, Jiang XF (2017) MaxEnt-based prediction of overwintering areas of Loxostege sticticalis in China under different climate change scenarios. Acta Ecologica Sinica, 37, 4852-4863. (in Chinese with English abstract) |
| [唐继洪, 程云霞, 罗礼智, 张蕾, 江幸福 (2017) 基于MaxEnt模型的不同气候变化情景下我国草地螟越冬区预测. 生态学报, 37, 4852-4863.] | |
| [30] | Tang ZY, Fang JY (2004) A review on the elevational patterns of plant species diversity. Biodiversity Science, 12, 20-28. (in Chinese with English abstract) |
| [唐志尧, 方精云 (2004) 植物物种多样性的垂直分布格局. 生物多样性, 12, 20-28.] | |
| [31] | Vaz ÚL, Cunha HF, Nabout JC (2015) Trends and biases in global scientific literature about ecological niche models. Brazilian Journal of Biology, 75(4), S17-S24. |
| [32] | Wang HL (2017) Protection and utilization of wild animals and plants in Lanping County. Journal of Green Science and Technology, (3), 90-91. (in Chinese) |
| [汪海林 (2017) 兰坪县野生动植物保护与利用对策. 绿色科技, (3), 90-91.] | |
| [33] | Wiegand T, Gunatilleke S, Gunatilleke N (2007) Species associations in a heterogeneous Sri Lankan dipterocarp forest. The American Naturalist, 170, E77. |
| [34] | Yang GB (2011) Ecological quality assessment of Yunling Nature Reserve in Lanping County of Yunnan Province. Journal of West China Forestry Science, 40(4), 48-53. (in Chinese with English abstract) |
| [杨国斌 (2011) 兰坪云岭省级自然保护区生态质量评价研究. 西部林业科学, 40(4), 48-53.] | |
| [35] | Yang GB (2013) Lanping Yunling Nature Reserve. Yunnan Science and Technology Press, Kunming. (in Chinese) |
| [杨国斌 (2013) 兰坪云岭自然保护区. 云南科技出版社, 昆明.] | |
| [36] | Zhang L (2015a) Application of MaxEnt model in predicting the potential distribution of species. Bulletin of Biology, 50(11), 9-12. (in Chinese) |
| [张路 (2015a) MaxEnt最大熵模型在预测物种潜在分布范围方面的应用. 生物学通报, 50(11), 9-12.] | |
| [37] | Zhang L (2015b) Prediction of potential distribution area of Euphorbia dentata in China based on MaxEnt model. Journal of Biosafety, 24, 194-200. (in Chinese with English abstract) |
| [张路 (2015b) 基于MaxEnt模型预测齿裂大戟在中国的潜在分布区. 生物安全学报, 24, 194-200.] | |
| [38] | Zhao SQ, Fang JY, Lei GC (2000) Global 200: An approach to setting large-scale biodiversity conservation priorities. Chinese Biodiversity, 8, 435-440. (in Chinese with English abstract) |
| [赵淑清, 方精云, 雷光春 (2000) 全球200: 确定大尺度生物多样性优先保护的一种方法. 生物多样性, 8, 435-440.] | |
| [39] | Zhu GP, Qiao HJ (2016) Effect of the MaxEnt model’s complexity on the prediction of species potential distributions. Biodiversity Science, 24, 1189-1196. (in Chinese with English abstract) |
| [朱耿平, 乔慧捷 (2016) MaxEnt模型复杂度对物种潜在分布区预测的影响. 生物多样性, 24, 1189-1196.] |
| [1] | 付梦娣, 朱彦鹏, 任月恒, 李爽, 秦乐, 谢正君, 王清春, 张立博. 新疆野生动物通道空间布局优化[J]. 生物多样性, 2025, 33(3): 24346-. |
| [2] | 张琼悦, 邓卓迪, 胡学斌, 丁志锋, 肖荣波, 修晨, 吴政浩, 汪光, 韩东晖, 张语克, 梁健超, 胡慧建. 粤港澳大湾区城市化进程对区域内鸟类分布及栖息地连通性的影响[J]. 生物多样性, 2023, 31(3): 22161-. |
| [3] | 杨华林, 程跃红, 周天祥, 冯茜, 胡强, 张贵权, 杨建, 张晋东, 王彬, 周材权. 四川卧龙国家级自然保护区多空间尺度下绿尾虹雉的生境选择[J]. 生物多样性, 2022, 30(7): 21535-. |
| [4] | 滕继荣, 刘兴明, 何礼文, 王钧亮, 黄建, 冯杰, 王放, 翁悦. 甘肃白水江国家级自然保护区林缘社区饲养犬只对大熊猫时空节律的影响[J]. 生物多样性, 2022, 30(1): 21204-. |
| [5] | 张晨, 马伟, 陈晨, 汪沐阳, 徐文轩, 杨维康. 重大工程影响下新疆卡拉麦里山有蹄类野生动物自然保护区鹅喉羚的生境格局变化[J]. 生物多样性, 2022, 30(1): 21176-. |
| [6] | 马星, 王浩, 余蔚, 杜勇, 梁健超, 胡慧建, 邱胜荣, 刘璐. 基于MaxEnt模型分析广东省鸟类多样性热点分布及保护空缺[J]. 生物多样性, 2021, 29(8): 1097-1107. |
| [7] | 周润, 慈秀芹, 肖建华, 曹关龙, 李捷. 气候变化对亚热带常绿阔叶林优势类群樟属植物的影响及保护评估[J]. 生物多样性, 2021, 29(6): 697-711. |
| [8] | 高梅香, 林琳, 常亮, 孙新, 刘冬, 吴东辉. 土壤动物群落空间格局和构建机制研究进展[J]. 生物多样性, 2018, 26(10): 1034-1050. |
| [9] | 陈彦清, 曹永生, 吴彦澎, 陈丽娜, 方沩. 基于空间统计学分析农作物种质资源的空间分布特征[J]. 生物多样性, 2017, 25(11): 1213-1222. |
| [10] | 谭珊珊, 王忍忍, 龚筱羚, 蔡佳瑶, 沈国春. 群落物种及结构多样性对森林地上生物量的影响及其尺度效应: 以巴拿马BCI样地为例[J]. 生物多样性, 2017, 25(10): 1054-1064. |
| [11] | 朱耿平, 乔慧捷. Maxent模型复杂度对物种潜在分布区预测的影响[J]. 生物多样性, 2016, 24(10): 1189-1196. |
| [12] | 任思远, 王婷, 祝燕, 叶永忠, 李聪, 潘娜, 叶永忠. 暖温带-北亚热带过渡带落叶阔叶林群落不同径级系统发育结构的变化[J]. 生物多样性, 2014, 22(5): 574-582. |
| [13] | 高程, 郭良栋. 外生菌根真菌多样性的分布格局与维持机制研究进展[J]. 生物多样性, 2013, 21(4): 488-498. |
| [14] | 朱耿平, 刘国卿, 卜文俊, 高玉葆. 生态位模型的基本原理及其在生物多样性保护中的应用[J]. 生物多样性, 2013, 21(1): 90-98. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
备案号:京ICP备16067583号-7
Copyright © 2022 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn