
生物多样性 ›› 2018, Vol. 26 ›› Issue (6): 601-610. DOI: 10.17520/biods.2017339 cstr: 32101.14.biods.2017339
徐开达, 卢衎尔*( ), 卢占晖, 戴乾
), 卢占晖, 戴乾
收稿日期:2017-12-28
接受日期:2018-03-07
出版日期:2018-06-20
发布日期:2018-09-11
通讯作者:
卢衎尔
作者简介:# 共同第一作者
基金资助:
Kaida Xu, Kaner Lu*(), Zhanhui Lu, Qian Dai
Received:2017-12-28
Accepted:2018-03-07
Online:2018-06-20
Published:2018-09-11
Contact:
Lu Kaner
About author:# Co-first authors
摘要:
根据韭山列岛自然保护区2015年11月, 2016年2月、5月、8月的调查资料, 运用相对重要性指数(index of relative importance, IRI)、生态位宽度以及生态位重叠指数等生态指标, 对调查海域的优势种虾类进行生态位特征分析, 以了解该海域内虾类组成及优势种间的关系。结果表明, 保护区共调查到虾类16种, 隶属8科12属, 均属暖温种或暖水种, 其中优势种(IRI > 500)有7种, 隶属于4科5属; 虾类相对资源量呈春秋季高, 夏冬季低的季节性分布特征。除脊尾白虾(Exopalaemon carinicauda)外, 优势种的时空生态位宽度值(时间生态位宽度与空间生态位宽度的乘积)均大于非优势种。日本鼓虾(Alpheus japonicus)和细巧仿对虾(Parapenaeopsis tenella)的时空二维生态位重叠值最大, 表明其对资源的利用具有相似性; 安氏白虾(Exopalaemon annandalei)和哈氏仿对虾(Parapenaeopsis hardwickii)的时空生态位重叠值最小, 说明二者相关性低。通过计算得知, 相对重要性指数(IRI)与生态位宽度对物种重要性排序结果有异: 时空生态位宽度值最大的为葛氏长臂虾(Palaemon gravieri), 而IRI值最高的为日本鼓虾。与相对重要性指数(IRI)相比, 生态位宽度指数不仅能反映物种对资源的利用能力, 还能反映物种分布是否均匀, 但这两者均不能反映生物量的多少。
徐开达, 卢衎尔, 卢占晖, 戴乾 (2018) 韭山列岛自然保护区虾类优势种生态位. 生物多样性, 26, 601-610. DOI: 10.17520/biods.2017339.
Kaida Xu, Kaner Lu, Zhanhui Lu, Qian Dai (2018) Ecological niche analysis of dominant shrimp species in the Jiushan Islands Marine Nature Reserve. Biodiversity Science, 26, 601-610. DOI: 10.17520/biods.2017339.
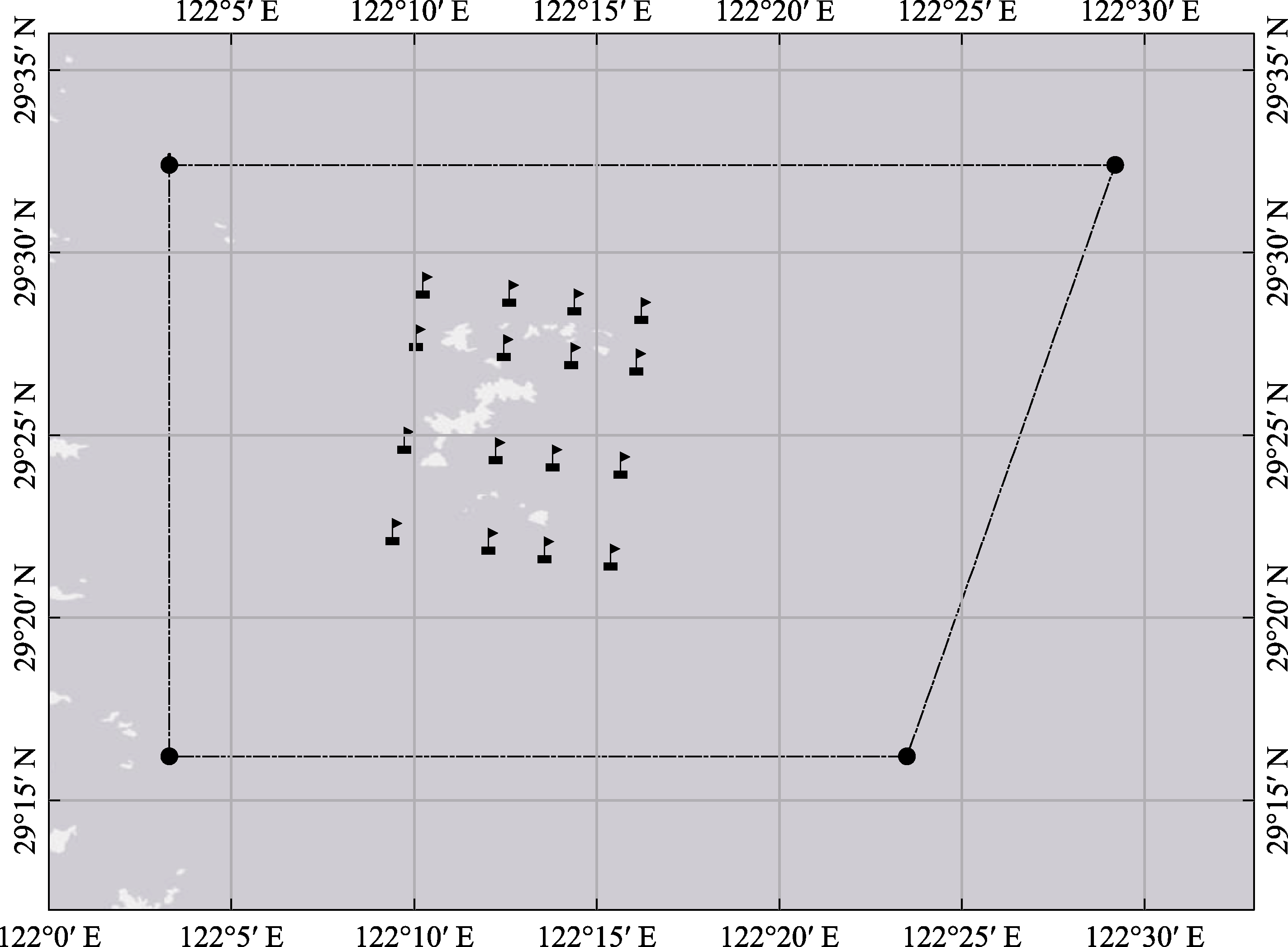
图1 研究海域及采样站位示意图(采样站位设置在保护区核心范围附近)
Fig. 1 Illustration of the investigated area and sampling sites (sampling sites were set around the core area of the Jiushan Islands Marine Nature Reserve)
| 科 Family | 属 Genus | 种 Species | 适温性 Optimum temperature | 出现月份 Occurrence | 相对重要性指数 Index of relative importance (IRI) |
|---|---|---|---|---|---|
| 对虾科 Penaeidae | 新对虾属 Metapenaeus | 周氏新对虾 Metapenaeus joyneri | WW | ○△▲ | 99.33 |
| 明对虾属 Fennerpenaeus | 长毛明对虾 Fenneropenaeus penicillatus | WW | ○△▲ | 7.75 | |
| 赤虾属 Metapenaeopsis | 戴氏赤虾 Metapenaeopsis dalei | WT | ○●△▲ | 4.62 | |
| 仿对虾属 Parapenaeopsis | 细巧仿对虾 Parapenaeopsis tenella | WT | ○●△▲ | 1,002.58 | |
| 哈氏仿对虾 Parapenaeopsis hardwickii | WW | ○●△▲ | 2,204.78 | ||
| 长臂虾科 Palaemonidae | 白虾属 Exopalaemon | 安氏白虾 Exopalaemon annandalei | WT | ○●△▲ | 3,944.06 |
| 脊尾白虾 Exopalaemon carinicauda | WT | ○△▲ | 752.25 | ||
| 长臂虾属 Palaemon | 葛氏长臂虾 Palaemon gravieri | WT | ○●△▲ | 3,008.04 | |
| 巨指长臂虾 Palaemon macrodactylus | WT | ●△▲ | 4.15 | ||
| 鼓虾科 Alpheidae | 鼓虾属 Alpheus | 日本鼓虾 Alpheus japonicus | WT | △▲ | 5,370.43 |
| 鲜明鼓虾 Alpheus distinguendus | WW | ○●△▲ | 374.41 | ||
| 藻虾科 Hippolytidae | 鞭腕虾属 Lysmata Risso | 鞭腕虾 Lysmata vittata | WW | ●▲ | 4.4 |
| 樱虾科 Sergestidae | 毛虾属 Acetes | 中国毛虾 Acetes chinensis | WW | ▲ | 123.28 |
| 异指虾科 Processidae | 异指虾属 Processa | 日本异指虾 Processa japonica | WW | △▲ | 0.89 |
| 管鞭虾科 Solenoceridae | 管鞭虾属 Solenocera | 中华管鞭虾 Solenocera crassicornis | WW | ●▲ | 2,183.67 |
| 玻璃虾科 Pasiphaeidae | 细螯虾属 Leptochela | 细螯虾 Leptochela gracilis | WW | ●▲ | 104.97 |
表1 2015-2016年韭山列岛自然保护区调查海域虾类组成
Table 1 Shrimp composition of Jiushan Islands Marine Nature Reserve in 2015-2016
| 科 Family | 属 Genus | 种 Species | 适温性 Optimum temperature | 出现月份 Occurrence | 相对重要性指数 Index of relative importance (IRI) |
|---|---|---|---|---|---|
| 对虾科 Penaeidae | 新对虾属 Metapenaeus | 周氏新对虾 Metapenaeus joyneri | WW | ○△▲ | 99.33 |
| 明对虾属 Fennerpenaeus | 长毛明对虾 Fenneropenaeus penicillatus | WW | ○△▲ | 7.75 | |
| 赤虾属 Metapenaeopsis | 戴氏赤虾 Metapenaeopsis dalei | WT | ○●△▲ | 4.62 | |
| 仿对虾属 Parapenaeopsis | 细巧仿对虾 Parapenaeopsis tenella | WT | ○●△▲ | 1,002.58 | |
| 哈氏仿对虾 Parapenaeopsis hardwickii | WW | ○●△▲ | 2,204.78 | ||
| 长臂虾科 Palaemonidae | 白虾属 Exopalaemon | 安氏白虾 Exopalaemon annandalei | WT | ○●△▲ | 3,944.06 |
| 脊尾白虾 Exopalaemon carinicauda | WT | ○△▲ | 752.25 | ||
| 长臂虾属 Palaemon | 葛氏长臂虾 Palaemon gravieri | WT | ○●△▲ | 3,008.04 | |
| 巨指长臂虾 Palaemon macrodactylus | WT | ●△▲ | 4.15 | ||
| 鼓虾科 Alpheidae | 鼓虾属 Alpheus | 日本鼓虾 Alpheus japonicus | WT | △▲ | 5,370.43 |
| 鲜明鼓虾 Alpheus distinguendus | WW | ○●△▲ | 374.41 | ||
| 藻虾科 Hippolytidae | 鞭腕虾属 Lysmata Risso | 鞭腕虾 Lysmata vittata | WW | ●▲ | 4.4 |
| 樱虾科 Sergestidae | 毛虾属 Acetes | 中国毛虾 Acetes chinensis | WW | ▲ | 123.28 |
| 异指虾科 Processidae | 异指虾属 Processa | 日本异指虾 Processa japonica | WW | △▲ | 0.89 |
| 管鞭虾科 Solenoceridae | 管鞭虾属 Solenocera | 中华管鞭虾 Solenocera crassicornis | WW | ●▲ | 2,183.67 |
| 玻璃虾科 Pasiphaeidae | 细螯虾属 Leptochela | 细螯虾 Leptochela gracilis | WW | ●▲ | 104.97 |
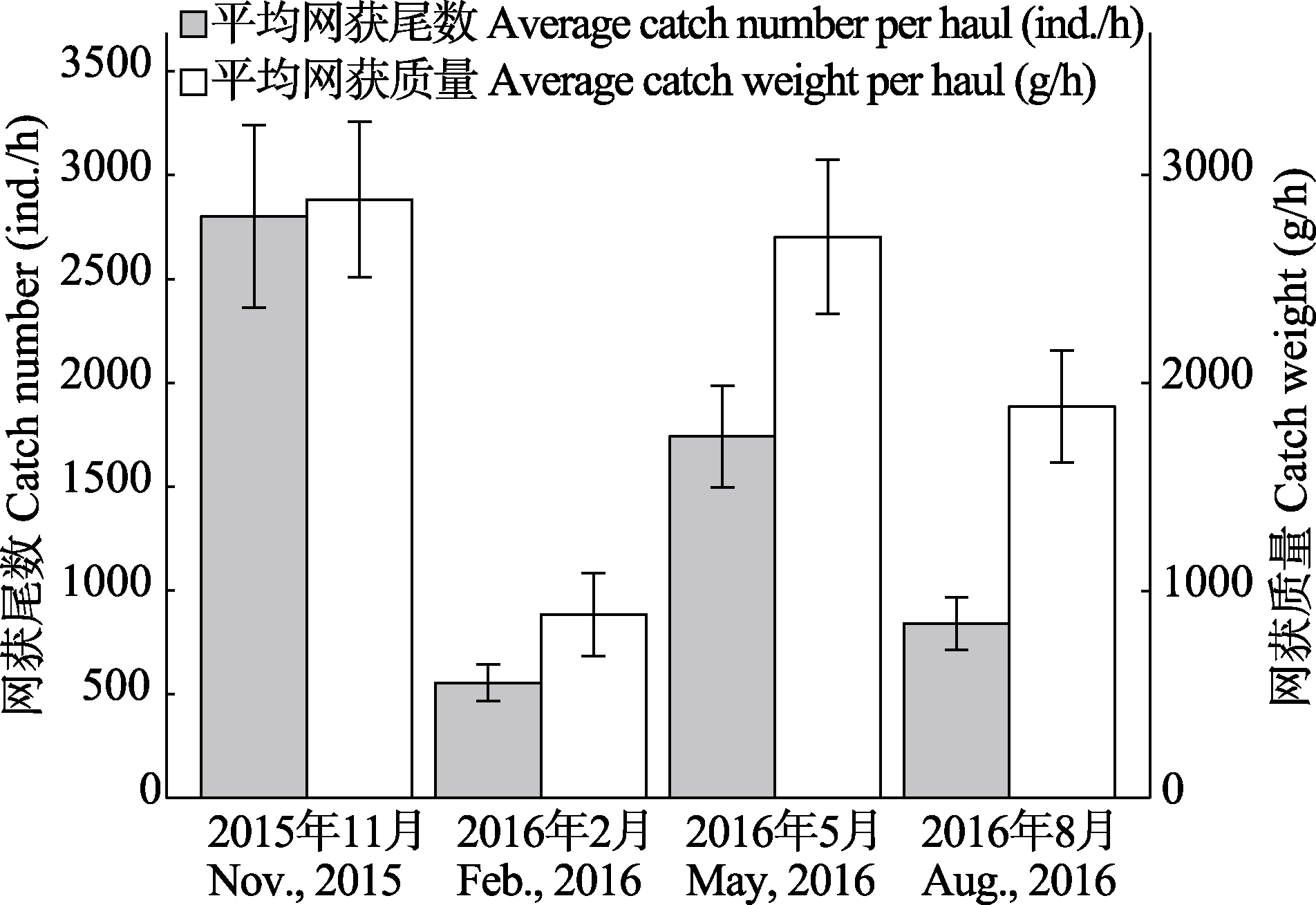
图2 调查海域虾类相对资源量随季节的变化(95%置信区间)
Fig. 2 The change of relative biomass resources of shrimps on a seasonal basis (95% confidence interval)
| 物种 Species | 空间生态位宽度 Spatial niche breadth (Bi-space) | 时间生态位宽度 Temporal niche breadth (Bi-time) | 时空二维生态位宽度 Spatio-temporal niche breadth (Bi) |
|---|---|---|---|
| 安氏白虾 Exopalaemon annandalei | 4.49 | 0.67 | 3.03 |
| 葛氏长臂虾 Palaemon gravieri | 4.54 | 1.04 | 4.71 |
| 哈氏仿对虾 Parapenaeopsis hardwickii | 2.42 | 0.60 | 1.46 |
| 脊尾白虾 Exopalaemon carinicauda | 1.40 | 0.31 | 0.43 |
| 日本鼓虾 Alpheus japonicus | 5.18 | 0.73 | 3.76 |
| 细巧仿对虾 Parapenaeopsis tenella | 2.51 | 0.64 | 1.61 |
| 中华管鞭虾 Solenocera crassicornis | 3.14 | 0.75 | 2.35 |
表2 优势种的时间、空间以及时空二维生态位宽度对比
Table 2 Comparisons of dominant shrimps among spatial, temporal and spatio-temporal niche breadths
| 物种 Species | 空间生态位宽度 Spatial niche breadth (Bi-space) | 时间生态位宽度 Temporal niche breadth (Bi-time) | 时空二维生态位宽度 Spatio-temporal niche breadth (Bi) |
|---|---|---|---|
| 安氏白虾 Exopalaemon annandalei | 4.49 | 0.67 | 3.03 |
| 葛氏长臂虾 Palaemon gravieri | 4.54 | 1.04 | 4.71 |
| 哈氏仿对虾 Parapenaeopsis hardwickii | 2.42 | 0.60 | 1.46 |
| 脊尾白虾 Exopalaemon carinicauda | 1.40 | 0.31 | 0.43 |
| 日本鼓虾 Alpheus japonicus | 5.18 | 0.73 | 3.76 |
| 细巧仿对虾 Parapenaeopsis tenella | 2.51 | 0.64 | 1.61 |
| 中华管鞭虾 Solenocera crassicornis | 3.14 | 0.75 | 2.35 |
| 安氏白虾 Exopalaemon annandalei | 葛氏长臂虾 Palaemon gravieri | 哈氏仿对虾 Parapenaeopsis hardwickii | 脊尾白虾 Exopalaemon carinicauda | 日本鼓虾 Alpheus japonicus | 细巧仿对虾 Parapenaeopsis tenella | 中华管鞭虾 Solenocera crassicornis | |
|---|---|---|---|---|---|---|---|
| 安氏白虾 Exopalaemon annandalei | 0.65 | 0.03 | 0.72 | 0.24 | 0.38 | 0.14 | |
| 葛氏长臂虾 Palaemon gravieri | 0.67 | 0.75 | 0.69 | 0.24 | 0.52 | 0.82 | |
| 哈氏仿对虾 Parapenaeopsis hardwickii | 0.50 | 0.80 | 0.13 | 0.11 | 0.39 | 0.99 | |
| 脊尾白虾 Exopalaemon carinicauda | 0.54 | 0.85 | 0.53 | 0.37 | 0.46 | 0.21 | |
| 日本鼓虾 Alpheus japonicus | 0.61 | 0.90 | 0.58 | 0.89 | 0.95 | 0.08 | |
| 细巧仿对虾 Parapenaeopsis tenella | 0.65 | 0.72 | 0.58 | 0.51 | 0.77 | 0.37 | |
| 中华管鞭虾 Solenocera crassicornis | 0.58 | 0.83 | 0.90 | 0.65 | 0.71 | 0.56 |
表3 韭山列岛自然保护区优势种间空间生态位(对角线以下)和时间生态位(对角线以上)重叠值
Table 3 Spatial niche overlaps (under the diagonal) and temporal niche overlaps (above the diagonal) between dominant shrimp species in the Jiushan Islands Marine Nature Reserve
| 安氏白虾 Exopalaemon annandalei | 葛氏长臂虾 Palaemon gravieri | 哈氏仿对虾 Parapenaeopsis hardwickii | 脊尾白虾 Exopalaemon carinicauda | 日本鼓虾 Alpheus japonicus | 细巧仿对虾 Parapenaeopsis tenella | 中华管鞭虾 Solenocera crassicornis | |
|---|---|---|---|---|---|---|---|
| 安氏白虾 Exopalaemon annandalei | 0.65 | 0.03 | 0.72 | 0.24 | 0.38 | 0.14 | |
| 葛氏长臂虾 Palaemon gravieri | 0.67 | 0.75 | 0.69 | 0.24 | 0.52 | 0.82 | |
| 哈氏仿对虾 Parapenaeopsis hardwickii | 0.50 | 0.80 | 0.13 | 0.11 | 0.39 | 0.99 | |
| 脊尾白虾 Exopalaemon carinicauda | 0.54 | 0.85 | 0.53 | 0.37 | 0.46 | 0.21 | |
| 日本鼓虾 Alpheus japonicus | 0.61 | 0.90 | 0.58 | 0.89 | 0.95 | 0.08 | |
| 细巧仿对虾 Parapenaeopsis tenella | 0.65 | 0.72 | 0.58 | 0.51 | 0.77 | 0.37 | |
| 中华管鞭虾 Solenocera crassicornis | 0.58 | 0.83 | 0.90 | 0.65 | 0.71 | 0.56 |
| 安氏白虾 Exopalaemon annandalei | 葛氏长臂虾 Palaemon gravieri | 哈氏仿对虾 Parapenaeopsis hardwickii | 脊尾白虾 Exopalaemon carinicauda | 日本鼓虾 Alpheus japonicus | 细巧仿对虾 Parapenaeopsis tenella | |
|---|---|---|---|---|---|---|
| 葛氏长臂虾 Palaemon gravieri | 0.43 | |||||
| 哈氏仿对虾 Parapenaeopsis hardwickii | 0.02 | 0.60 | ||||
| 脊尾白虾 Exopalaemon carinicauda | 0.38 | 0.58 | 0.07 | |||
| 日本鼓虾 Alpheus japonicus | 0.14 | 0.22 | 0.07 | 0.33 | ||
| 细巧仿对虾 Parapenaeopsis tenella | 0.25 | 0.38 | 0.22 | 0.24 | 0.73 | |
| 中华管鞭虾 Solenocera crassicornis | 0.08 | 0.68 | 0.89 | 0.14 | 0.06 | 0.21 |
表4 优势种间时空二维生态位重叠值
Table 4 Spatio-temporal niche overlap between dominant shrimp species
| 安氏白虾 Exopalaemon annandalei | 葛氏长臂虾 Palaemon gravieri | 哈氏仿对虾 Parapenaeopsis hardwickii | 脊尾白虾 Exopalaemon carinicauda | 日本鼓虾 Alpheus japonicus | 细巧仿对虾 Parapenaeopsis tenella | |
|---|---|---|---|---|---|---|
| 葛氏长臂虾 Palaemon gravieri | 0.43 | |||||
| 哈氏仿对虾 Parapenaeopsis hardwickii | 0.02 | 0.60 | ||||
| 脊尾白虾 Exopalaemon carinicauda | 0.38 | 0.58 | 0.07 | |||
| 日本鼓虾 Alpheus japonicus | 0.14 | 0.22 | 0.07 | 0.33 | ||
| 细巧仿对虾 Parapenaeopsis tenella | 0.25 | 0.38 | 0.22 | 0.24 | 0.73 | |
| 中华管鞭虾 Solenocera crassicornis | 0.08 | 0.68 | 0.89 | 0.14 | 0.06 | 0.21 |
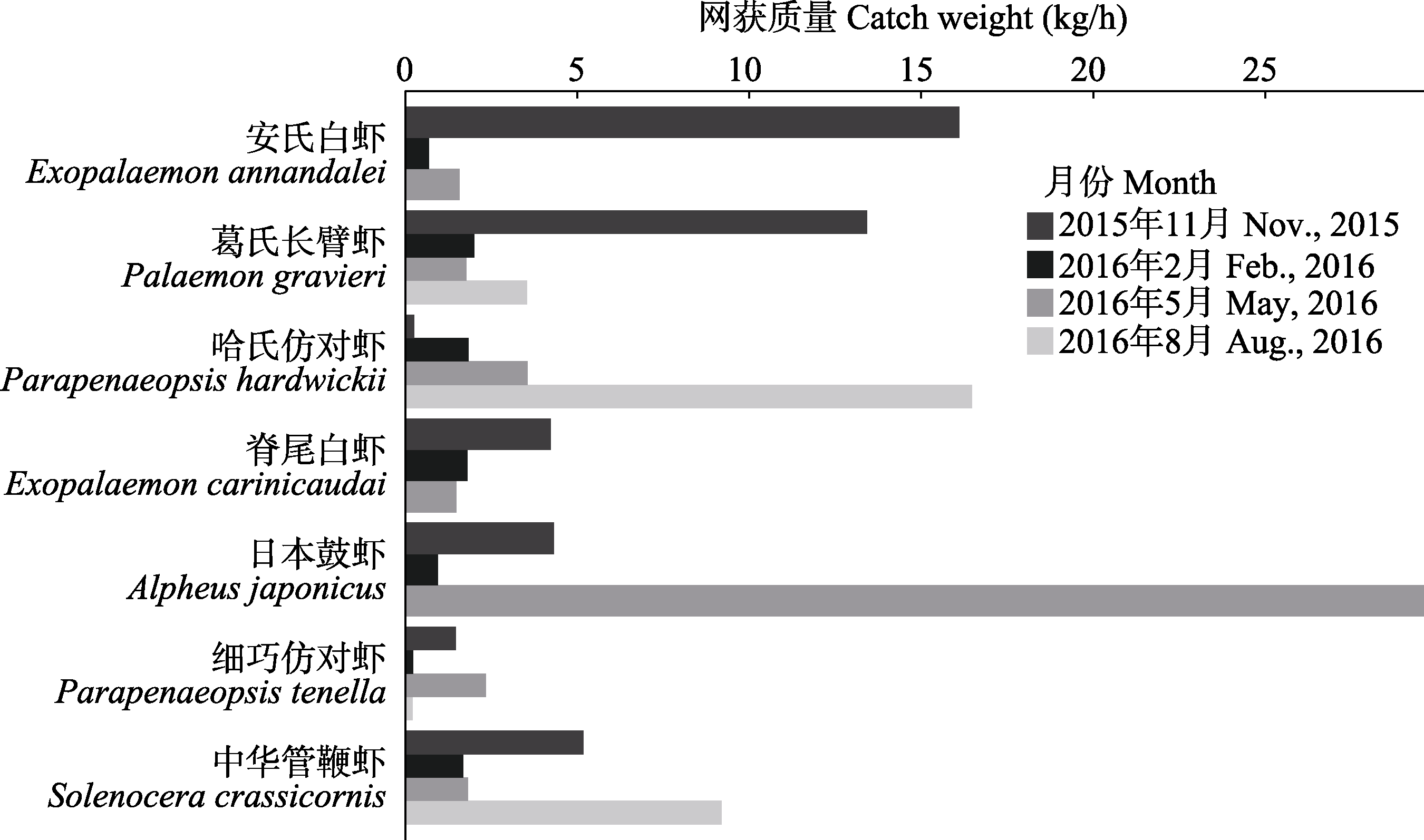
图3 调查区域各季节优势种虾类网获捕获重量变化
Fig. 3 The change of catch weight of dominant shrimp species in each season during the investigation
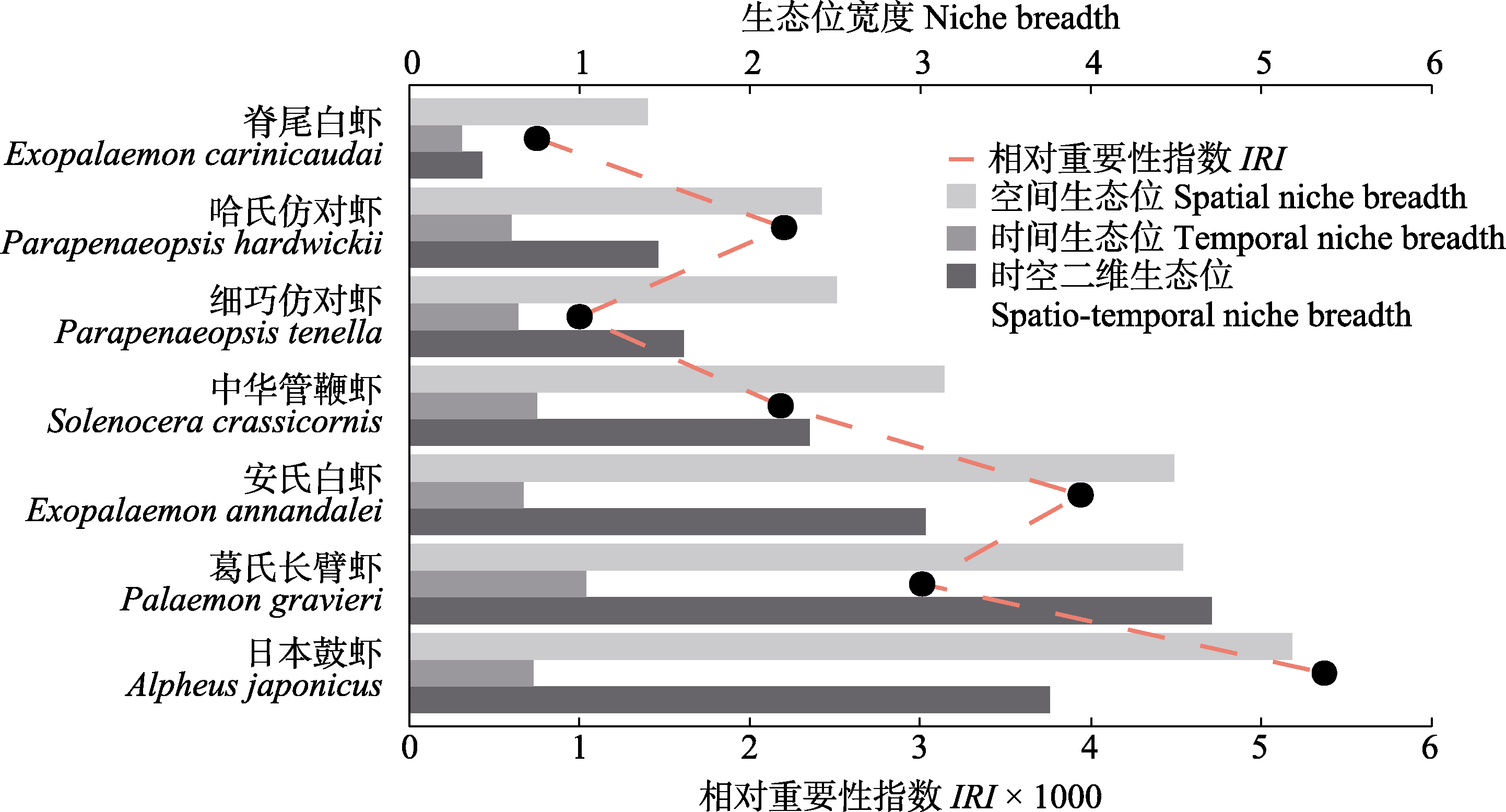
图4 优势种虾类的时间、空间生态位宽度, 时空生态位宽度以及相对重要性指数的比较
Fig. 4 Comparison of temporal niche breadth, spatial niche breadth, spatio-temporal niche breadth and IRI of dominant shrimp species
| [1] | Bennett JR, Cumming BF, Ginn BK, Smol JP (2010) Broad-scale environmental response and niche conservatism in lacustrine diatom communities. Global Ecology and Biogeography, 19, 724-732. |
| [2] | Chen WF, Peng X, Wang ZH, Xie QL, Chen SB, Ye S, Chen X, Ai WM (2017) Community structure characteristics of fishes in the coastal area of south Zhejiang during autumn and winter. Ocean Development and Management, 34(11), 111-119. (in Chinese with English abstract) |
| [陈伟峰, 彭欣, 汪振华, 谢起浪, 陈少波, 叶深, 陈骁, 艾未明 (2017) 浙南近海秋冬季鱼类群落结构研究. 海洋开发与管理, 34(11), 111-119.] | |
| [3] | Ge BM, Bao YX, Zheng X, Cheng HY (2005) The structure of the macrobenthic community and niche analysis at a tidal flat of Lingkun Island, China. Acta Ecologica Sinica, 25, 3037-3043. (in Chinese with English abstract) |
| [葛宝明, 鲍毅新, 郑祥, 程宏毅 (2005) 灵昆岛潮间带大型底栖动物群落结构与生态位分析. 生态学报, 25, 3037-3043.] | |
| [4] | General Administration of Quality Supervision, Inspection and Qurantine of the People’s Republic of China(2007) Specifications for Oceanographic Survey-Part 6: Marine Biological Survey. Standards Press of China, Beijing. (in Chinese) |
| [中华人民共和国国家质量监督检验检疫总局(2007) 海洋调查规范第6部分: 海洋生物调查. 中国标准出版社, 北京.] | |
| [5] | Guo TY, Xu RM (2003) Study on time niche among the ectoparasites of the rodent in Dongling Mountain in Beijing, China. Chinese Journal of Vector Biology and Control, 14, 30-32. (in Chinese with English abstract) |
| [郭天宇, 许荣满 (2003) 北京东灵山地区鼠类体外寄生虫时间生态位的研究. 中国媒介生物学及控制杂志, 14, 30-32.] | |
| [6] | Han DY, Xue Y, Ji YP, Xu BD, Liu H, Ma QY (2013) Trophic and spatial niche of five gobiid fishes in Jiaozhou Bay. Journal of Fishery Sciences of China, 20, 148-156. (in Chinese with English abstract) |
| [韩东燕, 薛莹, 纪毓鹏, 徐宾铎, 刘贺, 麻秋云 (2013) 胶州湾5种虾虎鱼类的营养和空间生态位. 中国水产科学, 20, 148-156.] | |
| [7] | He ZT, Xu KD, Xue LJ, Song HT (2009) Estimation of biological parameter, biomass, and yield for Palaemon gravieri in the northern area of East China Sea. Journal of Zhejiang Ocean University (Natural Science), 28, 286-291. (in Chinese with English abstract) |
| [贺舟挺, 徐开达, 薛利建, 宋海棠 (2009) 东海北部葛氏长臂虾生长死亡参数及资源量、渔获量的分析. 浙江海洋学院学报(自然科学版), 28, 286-291.] | |
| [8] | He ZT, Zhang HL, Xu KD, Liu ZF (2012) Diversity and distribution of benthic organisms in rocky intertidal zone of Jiushan Islands Nature Reserve, China. Fishery Information & Strategy, 27(3), 151-156. (in Chinese with English abstract) |
| [贺舟挺, 张洪亮, 徐开达, 刘子藩 (2012) 韭山列岛自然保护区岩相潮间带底栖生物多样性与分布. 渔业信息与战略, 27(3), 151-156.] | |
| [9] | Li DZ, Shi Q, Zang RG, Wang XP, Sheng LJ, Zhu ZL, Wang CA (2006) Models for niche breadth and niche overlap of species or populations, China. Scientia Silvae Sinicae, 42(7), 95-103. (in Chinese with English abstract) |
| [李德志, 石强, 臧润国, 王绪平, 盛丽娟, 朱志玲, 王长爱 (2006) 物种或种群生态位宽度与生态位重叠的计测模型. 林业科学, 42(7), 95-103.] | |
| [10] | Li R, Li SC, Tian RJ (2007) Structure and niche of spider community in vegetable field, China. Acta Arachnologica Sinica, 16, 116-120. (in Chinese with English abstract) |
| [李锐, 李生才, 田瑞钧 (2007) 菜田蜘蛛群落组成及生态位分析. 蛛形学报, 16, 116-120.] | |
| [11] | Liu RY (1963) Zoogeographical studies on the macrurous crustacean fauna of the Yellow Sea and the East China Sea. Oceanologia et Limnologia Sinica, 5(3), 230-244. (in Chinese with English abstract) |
| [刘瑞玉 (1963) 黄、东海虾类动物地理学研究. 海洋与湖沼, 5(3), 230-244.] | |
| [12] | Liu RY (2008) Checklist of Marine Biota of China Seas. Science Press, Beijing. (in Chinese) |
| [刘瑞玉 (2008) 中国海洋生物名录. 科学出版社, 北京.] | |
| [13] | Lu ZH, Xu HX, Xue LJ, Zhu WB (2015) A study on seasonal changes of the shrimp communities in the middle East China Sea. Journal of Zhejiang Ocean University (Natural Science), 34, 497-504. (in Chinese with English abstract) |
| [卢占晖, 徐汉祥, 薛利建, 朱文斌 (2015) 东海中部虾类群落的季节变化研究. 浙江海洋学院学报(自然科学版), 34, 497-504.] | |
| [14] | Lu ZH, Xue LJ, Zhang L, Xu KD, Zhang YZ (2013) Assessment of shrimp biomass in continental shelf waters of the East China Sea, China. Acta Hydrobiologica Sinica, 37, 855-862. (in Chinese with English abstract) |
| [卢占晖, 薛利建, 张龙, 徐开达, 张亚洲 (2013) 东海大陆架虾类资源量评估. 水生生物学报, 37, 855-862.] | |
| [15] | Lu ZH, Zhou YD, Zhu WB, Xu KD (2018) Spatial niches analysis of dominant shrimp species in the coastal area of Zhejiang Province. Acta Oceanologica Sinica, 40(2), 77-86. (in Chinese with English abstract) |
| [卢占晖, 周永东, 朱文斌, 徐开达 (2018) 浙江沿岸海域虾类优势种空间生态位分析. 海洋学报, 40(2), 77-86.] | |
| [16] | Magurran AE (1988) Ecological Diversity and Its Measurement. Princeton University Press, New Jersey. |
| [17] | May RM (1975) Some notes on estimating the competition matrix. Ecology, 56, 737-741. |
| [18] | Peng SY, Li XZ, Wang HF, Zhang BL (2015) Niche analysis of dominant species of macrozoobenthic community in the southern Yellow Sea in spring. Acta Ecologica Sinica, 35, 1917-1928. (in Chinese with English abstract) |
| [彭松耀, 李新正, 王洪法, 张宝琳 (2015) 南黄海春季大型底栖动物优势种生态位. 生态学报, 35, 1917-1928.] | |
| [19] | Pianka ER (1973) The structure of lizard communities. Annual Review of Ecology & Systematics, 4(41), 53-74. |
| [20] | Pinkas L, Oliphant MS, Iverson ILK (1971) Food habits of albacore, bluefin tuna, and bonito in California waters. Fish Bulletin, 152, 1-105. |
| [21] | Qi HM, Sun Y, Xu ZL, Sun LF, Gao Q, Que JL, Tian W (2013) Shrimp community structure and its influential factors in the Jiaojiang River estuary during spring and autumn. Chinese Journal of Applied Ecology, 24, 3546-3552. (in Chinese with English abstract) |
| [齐海明, 孙岳, 徐兆礼, 孙鲁峰, 高倩, 阙江龙, 田伟 (2013) 椒江口海域春秋季虾类群落结构及其影响因素. 应用生态学报, 24, 3546-3552.] | |
| [22] | R Core Team (2016) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna. |
| [23] | Shi YH, Zhang GY, Liu JZ, Zhu YZ, Zhu XD (2009) Growth and reproductive characteristics of Exopalaemon carinicauda bred in different months. Fisheries Science & Technology Information, 36(3), 131-136. (in Chinese with English abstract) |
| [施永海, 张根玉, 刘建忠, 朱雅珠, 朱晓东 (2009) 不同月份出生的脊尾白虾之生长及生殖特性. 水产科技情报, 36(3), 131-136.] | |
| [24] | Song HT (2002) The ecological colony and fauna characteristics of East China Sea shrimp. Studia Marina Sinica, 44, 124-133. (in Chinese with English abstract) |
| [宋海棠 (2002) 东海虾类的生态群落与区系特征. 海洋科学集刊, 44, 124-133.] | |
| [25] | Song HT, Yu CG, Xue LJ (2009) Study on the biomass distribution and growth property of Parapenaeopsis hardwickii in the East China Sea. Acta Hydrobiologica Sinica, 33, 15-21. (in Chinese with English abstract) |
| [宋海棠, 俞存根, 薛利建 (2009) 东海哈氏仿对虾的数量分布和生长特性研究. 水生生物学报, 33, 15-21.] | |
| [26] | Song HT, Yu CG, Xue LJ (2012) Fishery Biology of Commercial Shrimp and Crab Species in the East China Sea. China Ocean Press, Beijing. (in Chinese) |
| [宋海棠, 俞存根, 薛利建 (2012) 东海经济虾蟹类渔业生物学. 海洋出版社, 北京.] | |
| [27] | Sun LH, Li XF, Wang YN, Jiao HF, Shen YQ, Zhang SF, He J (2014) The species and distribution of macrobenthos in the Nanjiushan Sea area. Ecological Science, 33(1), 32-37. (in Chinese with English abstract) |
| [孙立辉, 李祥付, 王一农, 焦海峰, 沈叶青, 张树枫, 何京 (2014) 南韭山周围海域大型底栖动物种类与分布. 生态科学, 33(1), 32-37.] | |
| [28] | Tang FH, Wu YM, Fan W, Shen XQ, Wang YL (2010) Preliminary discussion on phytoplankton distribution and its relation to the runoff in the Yangtze River estuary. Ecology and Environment Sciences, 19, 2934-2940. (in Chinese with English abstract) |
| [唐峰华, 伍玉梅, 樊伟, 沈新强, 王云龙 (2010) 长江口浮游植物分布情况及与径流关系的初步探讨. 生态环境学报, 19, 2934-2940.] | |
| [29] | Tang YB, Liao YB, Shou L, Zeng JN, Gao AG, Chen QZ (2016) Intertidal zone of Nanji Islands is a niche for dominant species of the microbenthic community. Acta Ecologica Sinica, 36, 489-498. (in Chinese with English abstract) |
| [汤雁滨, 廖一波, 寿鹿, 曾江宁, 高爱根, 陈全震 (2016) 南麂列岛潮间带大型底栖动物群落优势种生态位. 生态学报, 36, 489-498.] | |
| [30] | Tian MC, Sun BL, Yang JM (1993) An analysis of ichthyologic fauna in Bohai Sea. Studia Marina Sinica, 34, 157-167. (in Chinese) |
| [田明诚, 孙宝龄, 杨纪明 (1993) 渤海鱼类区系分析. 海洋科学集刊, 34, 157-167.] | |
| [31] | Wang YH, Wu H, Zhu JR, Shen J (2015) Numerical simulation on the phytoplankton dynamics in Changjiang estuary and Min-Zhe coastal waters. Journal of East China Normal University (Natural Science), (4), 97-109. (in Chinese with English abstract) |
| [王一鹤, 吴辉, 朱建荣, 沈健 (2015) 长江口及闽浙海域浮游植物动力学模型研究. 华东师范大学学报(自然科学版), (4), 97-109.] | |
| [32] | Wathne JA, Haug T, Lydersen C (2000) Prey preference and niche overlap of ringed seals Phoca hispida and harp seals P. groenlandica in the Barents Sea. Marine Ecology Progress, 194, 233-239. |
| [33] | Wu CW, Wang WH (1995) A study of biology and ecological distribution of Exopalaemon annandalei in Hangzhou Bay. Journal of Zhejiang Ocean University (Natural Science), 12, 21-31. (in Chinese) |
| [吴常文, 王伟洪 (1995) 杭州湾海域安氏白虾生物学与生态分布的研究. 浙江水产学院学报(自然科学版), 12, 21-31.] | |
| [34] | Xia LJ, Chen WD, Zheng J, Cai HC, Wu EW, Bi SY, Xie X, Yu CG (2016) Species composition and quantitative distribution of shrimp in the Nanji Islands Marine Conservation Area. Journal of Fishery Sciences of China, 23, 648-660. (in Chinese with English abstract) |
| [夏陆军, 陈万东, 郑基, 蔡厚才, 伍尔魏, 毕耜瑶, 谢旭, 俞存根 (2016) 南麂列岛海洋自然保护区的虾类种类组成和数量分布. 中国水产科学, 23, 648-660.] | |
| [35] | Xie X, Yu CG, Zheng J, Xia LJ, Bi SY, Zhang P, Deng XY, Miao L (2017) Community structure and relationship with environment in shrimp in Pishan waters at Yushan Fishing Ground. Journal of Dalian Ocean University, 32, 713-723. (in Chinese with English abstract) |
| [谢旭, 俞存根, 郑基, 夏陆军, 毕耜瑶, 张平, 邓小艳, 苗露 (2017) 鱼山渔场披山海域虾类群落结构及其与环境因子的关系. 大连海洋大学学报, 32, 713-723.] | |
| [36] | Xu CY, Yu QJ, Xu FJ, Hu XQ, You WH (2012) Niche analysis of phytoplankton’s dominant species in Dianshan Lake of East China. Chinese Journal of Applied Ecology, 23, 2550-2558. (in Chinese with English abstract) |
| [徐春燕, 俞秋佳, 徐凤洁, 胡雪芹, 由文辉 (2012) 淀山湖浮游植物优势种生态位. 应用生态学报, 23, 2550-2558.] | |
| [37] | Xu JP, Yang SY (1993) A discussion of environmental resources and its development in Sanmen Bay. Ocean Development and Management, (2), 14-16. (in Chinese) |
| [许建平, 杨士英 (1993) 三门湾环境资源及其综合开发探讨. 海洋与海岸带开发, (2), 14-16.] | |
| [38] | Xu JY (1990) A preliminary research of reproduction of Exopalaemon annandalei in Yueqing Bay, Zhejiang. Chinese Journal of Zoology, 25(6), 3-7. (in Chinese) |
| [徐君义 (1990) 浙江乐清湾脊尾白虾的繁殖和世代的初步研究. 动物学杂志, 25(6), 3-7.] | |
| [39] | Xu XQ, Zeng JN, Chen QZ, Liu JJ, Du P, Wang GZ (2013) Spatial niches of dominant zooplankton species in Sanmen Bay, Zhejiang Province of East China. Chinese Journal of Applied Ecology, 24, 818-824. (in Chinese with English abstract) |
| [徐晓群, 曾江宁, 陈全震, 刘晶晶, 杜萍, 王桂忠(2013) 浙江三门湾浮游动物优势种空间生态位. 应用生态学报, 24, 818-824.] | |
| [40] | Xu ZG, He Y, Yan BX, Song CC (2007) Niche characteristics of typical marsh wetland plant populations in Sanjiang Plain. Chinese Journal of Applied Ecology, 18, 783-787. (in Chinese with English abstract) |
| [徐治国, 何岩, 闫百兴, 宋长春 (2007) 三江平原典型沼泽湿地植物种群的生态位. 应用生态学报, 18, 783-787.] | |
| [41] | Xue LJ, He ZT, Xu KD, Song HT (2009) Population dynamics and estimation of sustainable yield for Solenocera crassicornis in the East China Sea. Journal of Fujian Fisheries, (4), 48-54. (in Chinese with English abstract) |
| [薛利建, 贺舟挺, 徐开达, 宋海棠 (2009) 东海中华管鞭虾种群动态及持续渔获量分析. 福建水产, (4), 48-54.] | |
| [42] | Yu ZH, Jin XS, Li XS (2010) Analysis of ecological niche for major fish species in the central and southern Yellow Sea. Marine Fisheries Research, 31(6), 1-8. (in Chinese with English abstract) |
| [于振海, 金显仕, 李显森 (2010) 黄海中南部主要鱼种的生态位分析. 渔业科学进展, 31(6), 1-8.] | |
| [43] | Zhang GL, Zhang JT (2002) Niche analysis of dominant species in Shenweigou of Guandi Mountain. Journal of Wuhan Botanical Research, 20, 203-208. (in Chinese with English abstract) |
| [张桂莲, 张金屯 (2002) 关帝山神尾沟优势种生态位分析. 武汉植物学研究, 20, 203-208.] | |
| [44] | Zhang HL, Xu KD, He ZT, Liu ZF, Zhou YD (2008) Analysis on the condition of fishery resources in the sea area around the Jiushan Archipelago. Marine Fisheries, 30, 105-113. (in Chinese with English abstract) |
| [张洪亮, 徐开达, 贺舟挺, 刘子藩, 周永东 (2008) 韭山列岛附近海域渔业资源分析. 海洋渔业, 30, 105-113.] | |
| [45] | Zhang JT (2011) Quantitative Ecology, 2nd edn. Science Press, Beijing. |
| [张金屯 (2011) 数量生态学, 第2版. 科学出版社, 北京.] | |
| [46] | Zhang YZ, He ZT (2013) Analysis on the fishery resources in the sea area of Jiushan Archipelago Oceanic Ecology Reserve during spring and summer. Journal of Zhejiang Ocean University (Natural Science), 32, 292-298. (in Chinese with English abstract) |
| [张亚洲, 贺舟挺 (2013) 春、夏季韭山列岛海洋生态自然保护区海域渔业资源分析. 浙江海洋学院学报(自然科学版), 32, 292-298.] |
| [1] | 吕晓波, 李东海, 杨小波, 张孟文. 红树林群落通过淹水时间及海水盐度的生态位分化实现物种共存[J]. 生物多样性, 2024, 32(3): 23302-. |
| [2] | 陈敏豪, 张超, 王嘉栋, 湛振杰, 陈君帜, 栾晓峰. 北美水貂和欧亚水獭在东北地区的分布与生态位重叠[J]. 生物多样性, 2023, 31(1): 22289-. |
| [3] | 郭朝丹, 赵彩云, 李飞飞, 李俊生. 天然林和人工林外来入侵和本地植物对比研究: 以弄岗国家级自然保护区为例[J]. 生物多样性, 2022, 30(4): 21356-. |
| [4] | 王剑, 董乙乂, 马丽滨, 潘勃, 马方舟, 丁晖, 胡亚萍, 彭艳琼, 吴孝兵, 王波. 西双版纳国家级自然保护区蚂蚁-树互作网络空间变异[J]. 生物多样性, 2020, 28(6): 695-706. |
| [5] | 孙蓓蓓, 俞存根, 刘惠, 颜文超, 张文俊, 戴冬旭. 南麂列岛东侧海域春秋季虾蟹类生物多样性[J]. 生物多样性, 2019, 27(7): 787-795. |
| [6] | 宋乃平, 王兴, 陈林, 薛毅, 陈娟, 随金明, 王磊, 杨新国. 荒漠草原“土岛”生境群落物种共存机制[J]. 生物多样性, 2018, 26(7): 667-677. |
| [7] | 孙燕, 周忠实, 王瑞, HeinzMüller-Schärer. 气候变化预计会减少东亚地区豚草的生物防治效果**[J]. 生物多样性, 2017, 25(12): 1285-1294. |
| [8] | 龚雪伟, 吕光辉. 艾比湖流域杜加依林荒漠植物群落多样性及优势种生态位[J]. 生物多样性, 2017, 25(1): 34-45. |
| [9] | 胡成业, 水玉跃, 田阔, 李良, 覃胡林, 张春草, 冀萌萌, 水柏年. 浙江七星列岛海洋特别保护区主要鱼类功能群划分及生态位分析[J]. 生物多样性, 2016, 24(2): 175-184. |
| [10] | 李渊, 张静, 张然, 宋普庆, 钟指挥, 王燕平, 林龙山. 南沙群岛西南部和北部湾口海域鱼类物种多样性[J]. 生物多样性, 2016, 24(2): 166-174. |
| [11] | 田中平, 庄丽, 李建贵, 程模香. 伊犁河谷北坡野果林木本植物种间关系 及环境解释[J]. 生物多样性, 2011, 19(3): 335-342. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
备案号:京ICP备16067583号-7
Copyright © 2022 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn