
生物多样性 ›› 2018, Vol. 26 ›› Issue (10): 1051-1059. DOI: 10.17520/biods.2018086 cstr: 32101.14.biods.2018086
王梦茹, 傅声雷, 徐海翔, 王美娜, 时雷雷*( )
)
收稿日期:2018-03-26
接受日期:2018-07-17
出版日期:2018-10-20
发布日期:2019-01-06
通讯作者:
时雷雷
作者简介:# 共同第一作者
基金资助:
Mengru Wang, Shenglei Fu, Haixiang Xu, Meina Wang, Leilei Shi*()
Received:2018-03-26
Accepted:2018-07-17
Online:2018-10-20
Published:2019-01-06
Contact:
Shi Leilei
About author:# Co-first authors
摘要:
马陆是陆地生态系统中物种多样性极高的大型土壤无脊椎动物类群。作为营腐生动物, 马陆在陆地生态系统中具有不可替代的重要功能。通过大量取食及随后的肠道过程, 马陆在很大程度上决定着陆地生态系统凋落物的破碎、转化和分解过程, 从而驱动碳和关键养分元素的循环周转。然而, 目前对马陆生态功能的研究还非常有限, 远远落后于其他土壤动物类群(如蚯蚓等)。本文初步总结了马陆的生态功能: (1)通过破碎、取食凋落物来加速凋落物的分解。马陆偏好取食半分解的凋落物, 其同化效率受到凋落物来源、温度和凋落物中微生物含量的影响。(2)主要通过取食和排泄等活动影响养分循环。但对于马陆如何影响土壤碳循环, 存在两种不同的观点: 一是马陆粪球的分解速率比凋落物更快, 加速了碳的循环; 二是马陆粪球更难分解, 有助于碳的固存和稳定。马陆破碎凋落物后, 凋落物释放氮素进入土壤。此外, 马陆的活动也影响土壤磷的循环, 提高土壤中有效磷的含量。(3)调控微生物特性, 与蚯蚓也有互作关系。通过以上三个方面的总结, 展望了未来马陆的主要研究方向, 以期引起更多思考和研究。
王梦茹, 傅声雷, 徐海翔, 王美娜, 时雷雷 (2018) 陆地生态系统中马陆的生态功能. 生物多样性, 26, 1051-1059. DOI: 10.17520/biods.2018086.
Mengru Wang, Shenglei Fu, Haixiang Xu, Meina Wang, Leilei Shi (2018) Ecological functions of millipedes in the terrestrial ecosystem. Biodiversity Science, 26, 1051-1059. DOI: 10.17520/biods.2018086.
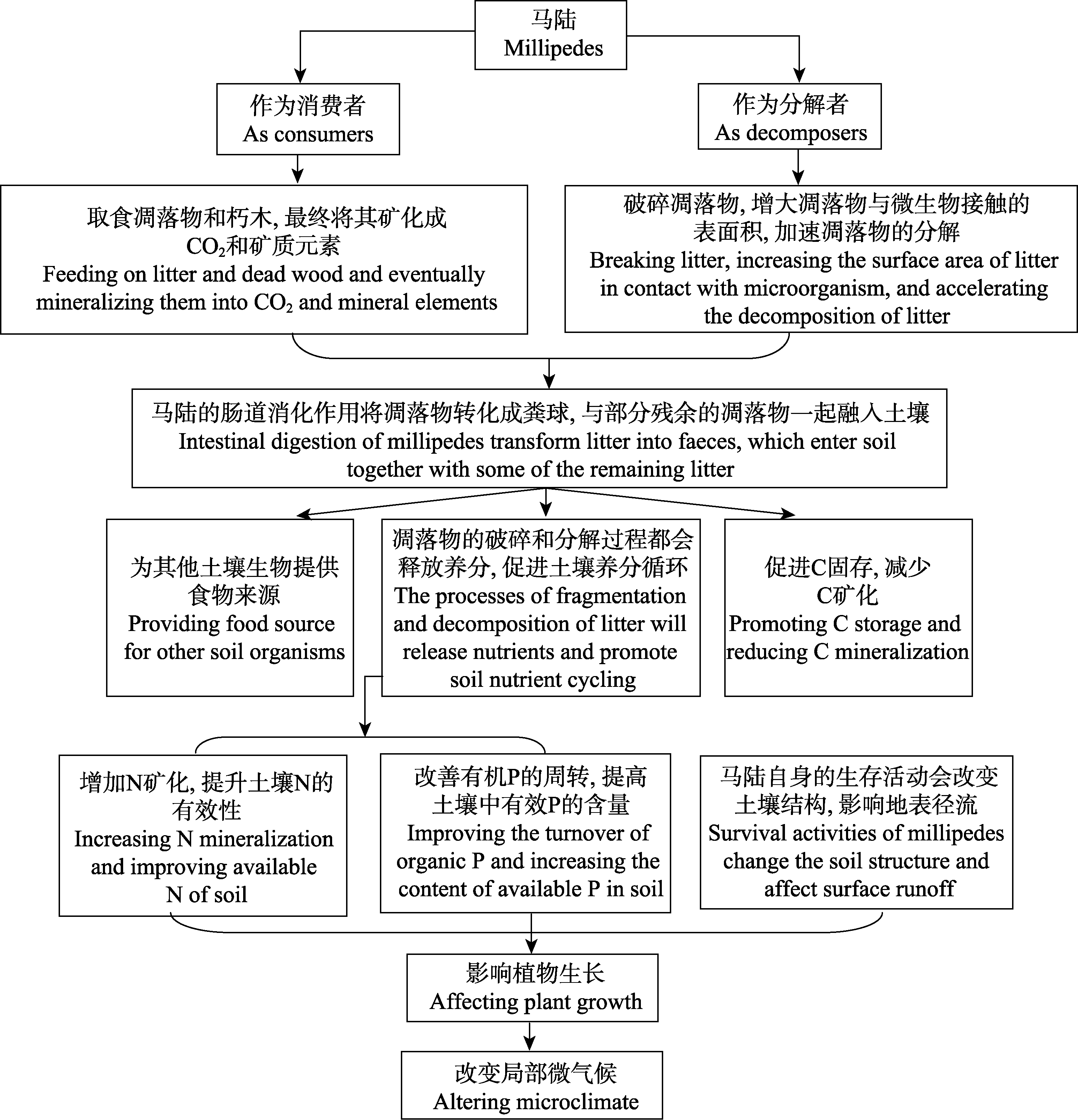
图1 马陆的生态功能
Fig. 1 Ecological functions of millipedes
| 目 Order | 科 Family | 属 Genus | 种 Species | 生存地点 Locality | 凋落物 Litters | 同化效率 Assimilation efficiency (%) | 参考文献 References |
|---|---|---|---|---|---|---|---|
| 球马陆目 Glomerida | Glomeridae | Glomeris | G. marginata | 荷兰栎树林 Oak forests in the Netherlands | 夏栎 Quercus robur | 9% | |
| 球马陆目 Glomerida | Glomeridae | Glomeris | G. marginata | 法国南部蒙彼利埃西北 25 km的冬青栎森林 Q. ilex forest of Puechabon, 25 km north-west of Montpellier (southern France) | 冬青栎 Quercus ilex | 6% | David & Gillon, 2002 |
| 带马陆目 Polydesmida | Xystodesmidae | Harpaphe | H.haydeniana | 加拿大马尔科姆·纳普研究森林—— 温哥华不列颠哥伦比亚大 学校园附近的成熟混合林 Malcolm Knapp Research Forest, near the University of British Columbia campus in Vancouver | 北美乔柏 Thuja plicata 西加云杉 Picea sitchensis 异叶铁杉 Tsuga heterophylla 花旗松 Pseudotsuga menziesii | 10% | Cárcamo et al, 2000 |
| 异蛩目 Spirostreptida | Spirostreptidae | Orthoporus | O. ornatus | 美国德克萨斯州大弯国家公园 Big Bend National Park, Texas, USA | 麻黄属 Ephedra 蜂蜜牧豆树 Prosopis glandulosa | 20-37% | Wooten & Crawford, 1975 |
| 山蛩目 Spirobolida | Pachybolidae | Trigoniulus | T. lumbricinus | 古巴 Cuba | 咖啡属 Coffea 木槿 Hibiscus 黍属 Panicum | 55% | Pobozsny et al, 1992 |
表1 马陆的同化效率
Table 1 Assimilation efficiency of millipedes
| 目 Order | 科 Family | 属 Genus | 种 Species | 生存地点 Locality | 凋落物 Litters | 同化效率 Assimilation efficiency (%) | 参考文献 References |
|---|---|---|---|---|---|---|---|
| 球马陆目 Glomerida | Glomeridae | Glomeris | G. marginata | 荷兰栎树林 Oak forests in the Netherlands | 夏栎 Quercus robur | 9% | |
| 球马陆目 Glomerida | Glomeridae | Glomeris | G. marginata | 法国南部蒙彼利埃西北 25 km的冬青栎森林 Q. ilex forest of Puechabon, 25 km north-west of Montpellier (southern France) | 冬青栎 Quercus ilex | 6% | David & Gillon, 2002 |
| 带马陆目 Polydesmida | Xystodesmidae | Harpaphe | H.haydeniana | 加拿大马尔科姆·纳普研究森林—— 温哥华不列颠哥伦比亚大 学校园附近的成熟混合林 Malcolm Knapp Research Forest, near the University of British Columbia campus in Vancouver | 北美乔柏 Thuja plicata 西加云杉 Picea sitchensis 异叶铁杉 Tsuga heterophylla 花旗松 Pseudotsuga menziesii | 10% | Cárcamo et al, 2000 |
| 异蛩目 Spirostreptida | Spirostreptidae | Orthoporus | O. ornatus | 美国德克萨斯州大弯国家公园 Big Bend National Park, Texas, USA | 麻黄属 Ephedra 蜂蜜牧豆树 Prosopis glandulosa | 20-37% | Wooten & Crawford, 1975 |
| 山蛩目 Spirobolida | Pachybolidae | Trigoniulus | T. lumbricinus | 古巴 Cuba | 咖啡属 Coffea 木槿 Hibiscus 黍属 Panicum | 55% | Pobozsny et al, 1992 |
| [1] | Adis J (2002) Taxonomical classification and biodiversity. ACM Transactions on Graphics, 33, 1-10. |
| [2] | Allison SD, Treseder KK (2010) Warming and drying suppress microbial activity and carbon cycling in boreal forest soils. Global Change Biology, 14, 2898-2909. |
| [3] | Ambarish CN, Sridhar KR (2016) Chemical and microbial characterization of feed and faeces of two giant pill-millipedes from forests in the western Ghats of India. Pedosphere, 26, 861-871. |
| [4] | Anderson JM, Bignell DE (1980) Bacteria in the food, gut contents and faeces of the litter-feeding millipede Glomeris marginata (Villers). Soil Biology & Biochemistry, 12, 251-254. |
| [5] | Anderson JM, Ineson P (1984) Interactions Between Microorganisms and Soil Invertebrates in Nutrient Flux Pathways of Forest Ecosystems. Cambridge University Press, Cambridge. |
| [6] | Anderson JM, Ineson P, Huish SA (1983) Nitrogen and cation mobilization by soil fauna feeding on leaf litter and soil organic matter from deciduous woodlands. Soil Biology & Biochemistry, 15, 463-467. |
| [7] | Ashwini KM, Sridhar KR (2005) Leaf litter preference and conversion by a saprophagous tropical pill millipede, Arthrosphaera magna, Attems. Pedobiologia, 49, 307-316. |
| [8] | Bailey PT (1989) Millipede parasitoid Pelidnoptera nigripennis (F.) (Diptera: Sciomyzidae) for the biological control of the millipede Ommatoiulus moreleti (Lucas) (Diplopoda: Julida: Julidae) in Australia. Bulletin of Entomological Research, 7, 381-391. |
| [9] | Barlow CA (1957) A factorial analysis of distribution in three species of Diplopods. Tijdschrift Voor Entomologie, 100, 349-426. |
| [10] | Bardgett RD, Wardle DA (2010) Aboveground-belowground Linkages:Biotic Interactions, Ecosystem Processes, and Global Change. Oxford University Press, Oxford. |
| [11] | Bonkowski M, Scheu S, Schaefer M (1998) Interactions of earthworms (Octolasion lacteum), millipedes (Glomeris marginata) and plants (Hordelymus europaeus) in a beechwood on a basalt hill: Implications for litter decomposition and soil formation. Applied Soil Ecology, 9, 161-166. |
| [12] | Brewer MS, Spruill CL, Rao NS, Bond JE (2012) Phylogenetics of the millipede genus Brachycybe Wood, 1864 (Diplopoda: Platydesmida: Andrognathidae): Patterns of deep evolutionary history and recent speciation. Molecular Phylogenetics & Evolution, 64, 232-242. |
| [13] | Brogden MC, Cortes C, Vandevoort AR, Snyder BA (2018) Soil nitrification analysis and millipede contribution. Georgia Journal of Science, 76, 117. |
| [14] | Buch AC, Sisinno CLS, Correia MEF, Silva-Filho EV (2018) Food preference and ecotoxicological tests with millipedes in litter contaminated with mercury. Science of the Total Environment, 633, 1173-1182. |
| [15] | Byzov BA, Tretyakova EB, Zvyagintsev DG, Claus H, Filip Z (1996) Effects of soil invertebrates on the survival of some genetically engineered bacteria in leaf litter and soil. Biology and Fertility of Soils, 23, 221-228. |
| [16] | Byzov BA, Kurakov AV, Tretyakova EB, Thanh VN, Luu ND, Rabinovich YM (1998a) Principles of the digestion of microorganisms in the gut of soil millipedes: Specificity and possible mechanisms. Applied Soil Ecology, 9, 145-151. |
| [17] | Byzov BA, Thanh VN, Babeva IP, Tretyakova EB, Dyvak IA, Yam R (1998b) Killing and hydrolytic activities of the gut fluid of the millipede Pachyiulus flavipes C. L. Koch on yeast cells. Soil Biology & Biochemistry, 30, 1137-1145. |
| [18] | Cárcamo HA, Abe TA, Prescott CE, Holl FB, Chanway CP (2000) Influence of millipedes on litter decomposition, N mineralization, and microbial communities in a coastal forest in British Columbia, Canada. Canadian Journal of Forest Research, 30, 817-826. |
| [19] | Choudhari CR, Dumbare YK, Theurkar SV (2014) Diversity of millipedes along the Northern Western Ghats, Rajgurunagar (MS), India (Arthropod: Diplopod). Journal of Entomology and Zoology Studies, 2, 254-257. |
| [20] | Coleman DC, Crossley DA (1996) Fundamentals of Soil Ecology. Academic Press, San Diego, CA. |
| [21] | Crawford CS (1975) Food, ingestion rates, and assimilation in the desert millipede Orthoporus ornatus (Girard) (Diplopoda). Oecologia, 20, 231-236. |
| [22] | David JF, Gillon D (2002) Annual feeding rate of the millipede Glomeris marginata, on holm oak (Quercus ilex) leaf litter under Mediterranean conditions. Pedobiologia, 46, 42-52. |
| [23] | Devi DS, Prabhoo NR (1990) Studies on food and feeding habits, food preference and feeding mechanism in the millipede Jonespeltis splendidus Verhoeff in captivity. Uttar Pradesh Journal Zoology, 10, 48-56. |
| [24] | Drift JVD (1975) Progress in Soil Zoology. Springer, the Netherlands. |
| [25] | Edwards CA, Hendrix PF (2004) Earthworm Ecology, 2nd edn. St. Lucie Press, Boca Raton. |
| [26] | Fan YL, Hu N, Ding SY, Liang GF, Lu XL (2016) Progress in terrestrial ecosystem services and biodiversity. Acta Ecologica Sinica, 36, 4583-4593. (in Chinese with English abstract) |
| [范玉龙, 胡楠, 丁圣彦, 梁国付, 卢训令 (2016) 陆地生态系统服务与生物多样性研究进展. 生态学报, 36, 4583-4593.] | |
| [27] | Frouz J, Špaldoňová A, Fričová K, Bartuška M (2014) The effect of earthworms (Lumbricus rubellus) and simulated tillage on soil organic carbon in a long-term microcosm experiment. Soil Biology & Biochemistry, 78, 58-64. |
| [28] | Fujimaki R, Sato Y, Okai N, Kaneko N (2010) The train millipede (Parafontaria laminata) mediates soil aggregation and N dynamics in a Japanese larch forest. Geogerma, 159, 216-220. |
| [29] | Golovatch SI, Kime RD (2009) Millipede (Diplopoda) distributions: A review. Soil Organisms, 81, 337-346. |
| [30] | Han HQ, Zhang JY, Ma G, Zhang XD, Bai YM (2018) Advances on impact of climate change on ecosystem services. Journal of Nanjing Forestry University (Natural Science Edition), 42, 184-190. (in Chinese with English abstract) |
| [韩会庆, 张娇艳, 马庚, 张新鼎, 白玉梅 (2018) 气候变化对生态系统服务影响的研究进展. 南京林业大学学报(自然科学版), 42, 184-190.] | |
| [31] | Hashimoto M, Kaneko N, Ito MT, Toyota A (2004) Exploitation of litter and soil by the train millipede Parafontaria laminata (Diplopoda: Xystodesmidae) in larch plantation forests in Japan. Pedobiologia, 48, 71-81. |
| [32] | He JZ, Lu YH, Fu BJ (2015) Frontiers of Soil Biology. Science Press, Beijing. (in Chinese) |
| [贺纪正, 陆雅海, 傅伯杰 (2015) 土壤生物学前沿. 科学出版社, 北京.] | |
| [33] | Holdsworth AR, Frelich LE, Reich PB (2007) Regional extent of an ecosystem engineer: Earthworm invasion in northern hardwood forests. Ecological Applications, 17, 1666-1677. |
| [34] | Hopkin SP, Read HJ (1992) The Biology of Millipedes. Quarterly Review of Biology. Oxford University Press, Oxford. |
| [35] | Iwashima N, Kaneko N, Sato K, Wakatsuki T, Masunaga, T (2011) Comparison of the faecal chemical properties of two geophagous millipede species Parafontaria laminata and Parafontaria tonominea (Xystodemidae) considering effects of habitat density and type of food. Edaphologia, 88, 43-53. (in Japanese with English summary) |
| [36] | Joly FX, Coulis M, Gérard A (2015) Litter-type specific microbial responses to the transformation of leaf litter into millipede feces. Soil Biology & Biochemistry, 86, 17-23. |
| [37] | Jousset A, Scheu S, Bonkowski M (2008) Secondary metabolite production facilitates establishment of rhizobacteria by reducing both protozoan predation and the competitive effects of indigenous bacteria. Functional Ecology, 22, 714-719. |
| [38] | Kaneko N (1999) Effect of millipede Parafontaria tonominea Attems (Diplopoda: Xystodesmidae) adults on soil biological activities: A microcosm experiment. Ecological Research, 14, 271-279. |
| [39] | Kheirallah AM (1990) Fragmentation of leaf litter by a natural population of the millipede Julus scandinavius (Latzel 1884). Biology and Fertility of Soils, 10, 202-206. |
| [40] | Köiuhler HR, Alberti G (1990) Morphology of the mandibles in the millipedes (Diplopoda, Arthropoda). Zoologica Scripta, 19, 195-202. |
| [41] | Lavelle P, Spain AV (2001) Soil Ecology. Kluwer Academic Publishers,Dordrecht. |
| [42] | Liao CH, Chen MQ, Chen JH (1992) Population ecology of two species of terrestrial isopods and their role in litter decomposition. Acta Zoologica Sinica, 38, 23-29. (in Chinese with English abstract) |
| [廖崇惠, 陈茂乾, 陈锦华 (1992) 两种陆栖等足类的种群及其分解落叶的作用. 动物学报, 38, 23-30.] | |
| [43] | Lyford WH (1943) Palatability of freshly fallen leaves of forest trees to millipedes. Ecology, 24, 252-261. |
| [44] | Makoto K, Arai M, Kaneko N (2014) Change the menu? Species-dependent feeding responses of millipedes to climate warming and the consequences for plant-soil nitrogen dynamics. Soil Biology & Biochemistry, 72, 19-25. |
| [45] | Maraun M, Scheu S (1996) Changes in microbial biomass, respiration and nutrient status of beech (Fagus sylvatica) leaf litter processed by millipedes (Glomeris marginata). Oecologia, 107, 131-140. |
| [46] | Marek PE, Bond JE (2006) Phylogenetic systematics of the colorful, cyanide-producing millipedes of appalachia (Polydesmida, Xystodesmidae, Apheloriini) using a total evidence Bayesian Approach. Molecular Phylogenetics & Evolution, 41, 704-729. |
| [47] | Naveed M, Moldrup P, Arthur E, Holmstrup M, Nicolaisen M, Tuller M, Herath L, Hamamoto S, Kawamoto K, Komatsu T, Vogeland H, Jonge L (2014) Simultaneous loss of soil biodiversity and functions along a copper contamination gradient: When soil goes to sleep. Soil Science Society of America Journal, 78, 1239-1250. |
| [48] | Oeyen JP, Wesener T (2018) A first phylogenetic analysis of the pill millipedes of the order glomerida, with a special assessment of mandible characters (Myriapoda, Diplopoda, Pentazonia). Arthropod Structure & Development, 47, 214-228. |
| [49] | Pobozsny M, Gonzales Oliver R, Rodriguez ME (1992) The role of Trigonoiulus lumbricinus Gerst. (Diplopoda) in the decomposition of leaf litter in some plant communities of Cuba. Opuscula Zoologica Budapest, 25, 89-93. |
| [50] | Rawlins AJ, Bull ID, Poirier N, Ineson P, Evershed RP (2006) The biochemical transformation of oak (Quercus robur) leaf litter consumed by the pill millipede (Glomeris marginata). Soil Biology & Biochemistry, 38, 1063-1076. |
| [51] | Reichle DE, Shanks MH, Crossley DA (1969) Calcium, potassium, and sodium content of forest floor arthropods. Annals of the Entomological Society of America, 62, 57-62. |
| [52] | Rong H, Fan HL, Li Q, Li J, Hong W, Wu CZ (2011) Effects of simulated nitrogen deposition on soil macrofauna in agroecosystem. Journal of North East Forestry University, 39(1), 85-88. (in Chinese with English abstract) |
| [荣海, 范海兰, 李茜, 李键, 洪伟, 吴承祯 (2011) 模拟氮沉降对农田大型土壤动物的影响. 东北林业大学学报, 39(1), 85-88.] | |
| [53] | Sakwa WN (1974) A consideration of the chemical basis of food preference in millipedes. Symposium of Zoological Society of London, 32, 329-346. |
| [54] | Scheu S, Wolters V (1991) Influence of fragmentation and bioturbation on the decomposition of 14C-labelled beech leaf litter. Soil Biology & Biochemistry, 23, 1029-1034. |
| [55] | Scheu S (1987) The role of substrate feeding earthworms (Lumbricidae) for bioturbation in a beechwood soil. Oecologia, 72, 192-196. |
| [56] | Shao YH, Zhang WX, Liu SJ, Wang XL, Fu SL (2015) Diversity and function of soil fauna. Acta Ecologica Sinica, 35, 6614-6625. (in Chinese with English abstract) |
| [邵元虎, 张卫信, 刘胜杰, 王晓丽, 傅声雷 (2015) 土壤动物多样性及其生态功能. 生态学报, 35, 6614-6625.] | |
| [57] | Shelley RM (2003) Taxonomy of extant Diplopoda (Millipeds) in the modern era: Perspectives for future advancements and observations on the global Diplopod community (Arthropoda: Diplopoda). Zootaxa, 1668, 343-362. |
| [58] | Shelley RM, Shear WA (2005) A new milliped of the genus Stenozonium Shelley 1998 from Washington State, U.S.A.: First record of the genus and family from North of the Columbia River (Polyzoniida: Polyzoniidae). Zootaxa, 1017, 25-32. |
| [59] | Sierwald P, Bond JE (2007) Current status of the Myriapod class Diplopoda (Millipedes): Taxonomic diversity and phylogeny. Annual Review of Entomology, 52, 401-420. |
| [60] | Silva VMD, Antoniolli ZI, Jacques RJS, Ott R, Andrade FV, Passos RR (2017) Influence of the tropical millipede, Glyphiulus granulatus (Gervais, 1847), on aggregation, enzymatic activity, and phosphorus fractions in the soil. Geoderma, 289, 135-141. |
| [61] | Smit AM, Van Aarde RJ (2001) The influence of millipedes on selected soil elements: A microcosm study on three species occurring on coastal sand dunes. Functional Ecology, 15, 51-59. |
| [62] | Snyder BA, Callaham MAJ, Lowe CN, Hendrix PF (2013) Earthworm invasion in North America: Food resource competition affects native millipede survival and invasive earthworm reproduction. Soil Biology & Biochemistry, 57, 212-216. |
| [63] | Suzuki Y, Grayston SJ, Prescott CE (2013) Effects of leaf litter consumption by millipedes (Harpaphe haydeniana) on subsequent decomposition depends on litter type. Soil Biology & Biochemistry, 57, 116-123. |
| [64] | Tajovský K, Santruckova H, Hanel L, Balik V, Lukesova A (1992) Decomposition of faecal pellets of the millipede Glomeris hexasticha (Diplopoda) in forest soil. Pedobiologia, 36, 146-158. |
| [65] | Toyota A, Kaneko N, Ito MT (2006) Soil ecosystem engineering by the train millipede Parafontaria laminata, in a Japanese larch forest. Soil Biology & Biochemistry, 38, 1840-1850. |
| [66] | Wang MN, Lu XL, Ding SY, Ren JY, Bian ZQ, Xu Z (2017) Pollinator diversity in different habitats of the agricultural landscape in the middle and lower reaches of the Yellow River based on the three-color pan trap method. Acta Ecologica Sinica, 37, 148-155. |
| [67] | Wolters V (2000) Invertebrate control of soil organic matter stability. Biology and Fertility of Soils, 31, 1-19. |
| [68] | Wooten RC, Crawford CS (1975) Food, ingestion rates, and assimilation in the desert millipede Orthoporus ornatus (Girard) (Diplopoda). Oecologia, 20, 231-236. |
| [69] | Yin XQ, Song B, Qiu LL (2007) Dynamic characteristic of N, P, K in the litter-soil fauna-soil system of mixed Pinus koraiensis and broad-leaved forest. Acta Ecologica Sinica, 27, 128-134. (in Chinese with English abstract) |
| [殷秀琴, 宋博, 邱丽丽 (2007) 红松阔叶混交林凋落物-土壤动物-土壤系统中N、P、K的动态特征. 生态学报, 27, 128-134.] | |
| [70] | Zhang WX, Chen DM, Zhao CC (2007) Functions of earthworm in ecosystem. Biodiversity Science, 15, 142-153. (in Chinese with English abstract) |
| [张卫信, 陈迪马, 赵灿灿 (2007) 蚯蚓在生态系统中的作用. 生物多样性, 15, 142-153.] | |
| [71] | Zhang XP, Li CY, Zhang SC (2001) Study of the function of millipedes in substance decomposition. Acta Ecologica Sinica, 21, 75-79. (in Chinese with English abstract) |
| [张雪萍, 李春艳, 张思冲 (2001) 马陆在森林生态系统物质转化中的功能研究. 生态学报, 21, 75-79.] |
| [1] | 张明燡, 王晓梅, 郑言鑫, 吴楠, 李东浩, 樊恩源, 李娜, 单秀娟, 于涛, 赵春暖, 李波, 徐帅, 吴玉萍, 任利群. 黄河口典型牡蛎礁分布区资源状况和栖息地功能[J]. 生物多样性, 2025, 33(4): 24208-. |
| [2] | 刘淑琪, 崔东, 江智诚, 刘江慧, 闫江超. 短期氮、水添加和刈割减弱了苦豆子型退化草地土壤生物多样性与生态系统多功能性的联系[J]. 生物多样性, 2025, 33(3): 24305-. |
| [3] | 陈丁松, 刘子恺, 贺子洋, 陈伟东. 缓步动物多样性、分布特征和生态功能研究进展[J]. 生物多样性, 2025, 33(2): 24406-. |
| [4] | 王嘉陈, 徐汤俊, 许唯, 张高季, 尤艺瑾, 阮宏华, 刘宏毅. 城市景观格局对大蚰蜒种群遗传结构的影响[J]. 生物多样性, 2025, 33(1): 24251-. |
| [5] | 马骅, 李常青, 余品锋, 陈杰, 贺天耀, 王可洪. 澎溪河消落带大型土壤动物群落分布格局及其影响因素[J]. 生物多样性, 2024, 32(7): 24117-. |
| [6] | 孙怡欣, 侯春雨, 周磊, 魏雪, 马金豪, 薛娟, 李小涵, 吴鹏飞. 青藏高原盆栽一年生和多年生豆科牧草对土壤线虫群落的影响[J]. 生物多样性, 2024, 32(7): 24040-. |
| [7] | 王党军, 谢午阳, 林小元, 乔秀娟, 徐耀粘, 田秋香, 刘峰, 张娅妮, 左娟, 江明喜. 八大公山森林土壤动物群落与叶经济谱及凋落物分解速率的关系[J]. 生物多样性, 2024, 32(12): 24261-. |
| [8] | 谢致敬, 刘相钰, 孙晓铭, 刘继亮, 刘占锋, 张晓珂, 陈军, 杨效东, 朱波, 柯欣, 吴东辉. 中国土壤动物多样性监测网络建设、进展与展望[J]. 生物多样性, 2023, 31(12): 23365-. |
| [9] | 潘雪, 刘冬. 2020-2021年世界甲螨亚目新分类单元和近15年中国发表新种概况——纪念中国甲螨学开创100周年[J]. 生物多样性, 2022, 30(12): 22193-. |
| [10] | 王军, 赵超. 中国菌食性管蓟马物种多样性及分布格局[J]. 生物多样性, 2022, 30(12): 22128-. |
| [11] | 李帆, 王党军, 林小元, 纪康, 叶露萍, 黄超, 郑勇, Zhun Mao, 左娟. 八大公山亚热带森林木质残体中大型无脊椎动物群落特征[J]. 生物多样性, 2022, 30(12): 21476-. |
| [12] | 姚海凤, 张赛超, 上官华媛, 李志鹏, 孙新. 城市化对土壤动物群落结构和多样性的影响[J]. 生物多样性, 2022, 30(12): 22547-. |
| [13] | 程建伟, 王亚东, 王桠楠, 李莹, 郭颖, 白正, 刘新民, 李永宏. 半干旱草原大中型土壤动物在畜粪分解中的作用[J]. 生物多样性, 2022, 30(12): 22575-. |
| [14] | 徐聪, 张飞宇, 俞道远, 孙新, 张峰. 土壤动物的分子分类预测策略评估[J]. 生物多样性, 2022, 30(12): 22252-. |
| [15] | 姚保民, 曾青, 张丽梅. 土壤原生生物多样性及其生态功能研究进展[J]. 生物多样性, 2022, 30(12): 22353-. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
备案号:京ICP备16067583号-7
Copyright © 2022 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn