
生物多样性 ›› 2018, Vol. 26 ›› Issue (2): 202-209. DOI: 10.17520/biods.2017305 cstr: 32101.14.biods.2017305
所属专题: 青藏高原生物多样性与生态安全; 生态文明建设
乔慧捷1, 汪晓意2, 王伟3, 罗振华4, 唐科2, 黄燕2,5, 杨胜男2,5, 曹伟伟6, 赵新全2,7, 江建平2, 胡军华2,*
收稿日期:2017-11-14
接受日期:2018-02-14
出版日期:2018-02-20
发布日期:2018-05-05
通讯作者:
胡军华
作者简介:# 共同第一作者
基金资助:Huijie Qiao1, Xiaoyi Wang2, Wei Wang3, Zhenhua Luo4, Ke Tang2, Yan Huang2,5, Shengnan Yang2,5, Weiwei Cao6, Xinquan Zhao2,7, Jianping Jiang2, Junhua Hu2,*
Received:2017-11-14
Accepted:2018-02-14
Online:2018-02-20
Published:2018-05-05
Contact:
Hu Junhua
About author:# Co-first authors
摘要:
三江源地区具有丰富的高原生物多样性, 是我国重要的生态安全屏障。作为我国第一个体制试点的国家公园, 三江源国家公园将被建成青藏高原大自然保护展示和生态文化传承区。为更好地服务于三江源国家公园建设, 本研究从海拔、年均温和年降水等方面, 在环境空间上系统比较了国家公园与三江源和可可西里自然保护区的差异; 以4种两栖爬行动物即高原林蛙(Rana kukunoris)、倭蛙(Nanorana pleskei)、西藏齿突蟾(Scutiger boulengeri)和青海沙蜥(Phrynocephalus vlangalii)分布点环境条件在国家公园内外的异同为例, 探讨分布受环境制约大、扩散能力弱的物种保护在国家公园建设中可能面临的机遇和挑战。结果表明, 国家公园与已建自然保护区在地理空间上重叠较大, 但它们在自然环境上的差别很明显; 自然保护区内两栖爬行类所偏好的环境条件较多未包含在国家公园内。鉴于三江源的区位特殊性及对气候变化的敏感性, 区域内两栖爬行动物等对环境依赖性强、扩散能力弱物种的有效保护不仅有助于保持物种遗传多样性和区域生态系统的完整性, 也有利于更好地实现国家公园目标定位并服务于生态文明建设。为此, 在国家公园内, 应开展两栖爬行动物种群动态和群落结构的长期监测, 加强基础生物学研究, 掌握环境变化对两栖爬行动物分布、遗传、行为、形态、种群动态及群落可能产生的影响, 实现区域内两栖爬行动物及其类似物种在环境变化下的永续生存。
乔慧捷, 汪晓意, 王伟, 罗振华, 唐科, 黄燕, 杨胜男, 曹伟伟, 赵新全, 江建平, 胡军华 (2018) 从自然保护区到国家公园体制试点: 三江源国家公园环境覆盖的变化及其对两栖爬行类保护的启示. 生物多样性, 26, 202-209. DOI: 10.17520/biods.2017305.
Huijie Qiao, Xiaoyi Wang, Wei Wang, Zhenhua Luo, Ke Tang, Yan Huang, Shengnan Yang, Weiwei Cao, Xinquan Zhao, Jianping Jiang, Junhua Hu (2018) From nature reserve to national park system pilot: Changes of environmental coverage in the Three-River-Source National Park and implications for amphibian and reptile conservation. Biodiversity Science, 26, 202-209. DOI: 10.17520/biods.2017305.
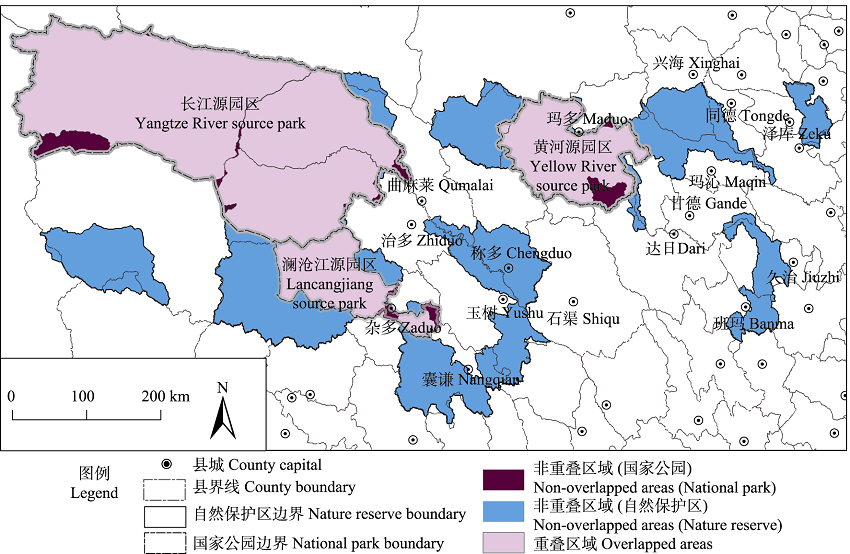
图1 三江源国家公园与自然保护区地理空间分布比较
Fig. 1 The differences of geographic distributions between the Three-River-Source National Park and the nature reserves
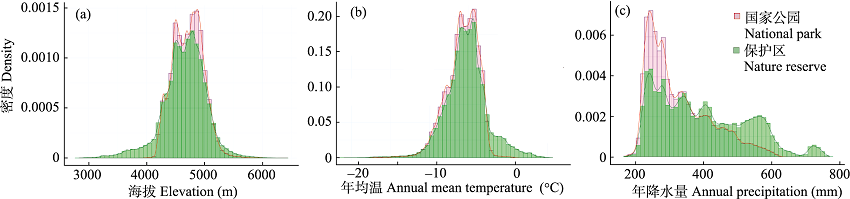
图2 三江源国家公园与自然保护区海拔、年均温和年降水量不同水平上的密度比较
Fig. 2 The comparisons of the density in different levels in elevation, annual mean temperature and annual precipitation between the Three-River-Source National Park and the nature reserves
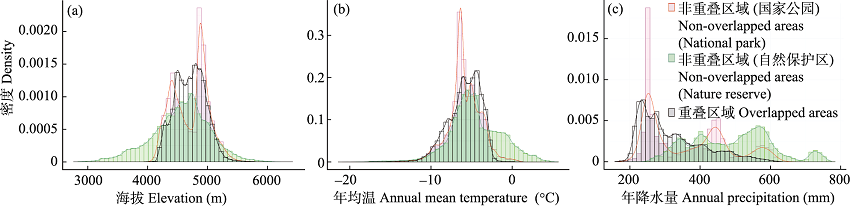
图3 三江源国家公园与自然保护区重叠与非重叠区域海拔、年均温和年降水量比较
Fig. 3 The comparisons between the overlapped and non-overlapped areas of the Three-River-Source National Park and the nature reserves in elevation, annual mean temperature and annual precipitation
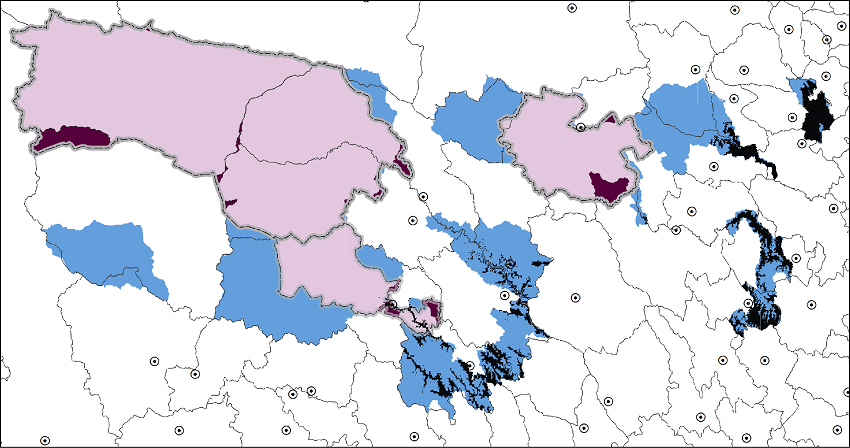
图4 同时满足3个条件(海拔 < 4,200 m、年均温 > -3℃且年降水 > 500 mm)区域(黑色)的空间分布
Fig. 4 The spatial distribution of areas (black) with the combined condition of elevation < 4,200 m, annual temperature > -3℃ and annual precipitation > 500 mm
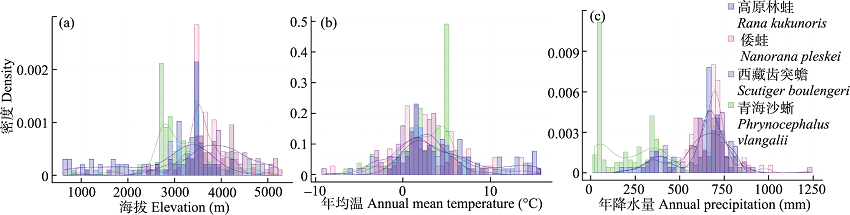
图5 高原林蛙、倭蛙、西藏齿突蟾和青海沙蜥分布点的海拔、年均温和年降水量直方图与密度曲线
Fig. 5 The histograms and density curves of elevation, annual mean temperature and annual precipitation for the known occurrences of Rana kukunoris, Nanorana pleskei, Scutiger boulengeri, and Phrynocephalus vlangalii
| 变量 Variable | 阈值 Threshold | 物种 Species | 分布点数量 No. of occurrences | 不属于国家公园环境空间的比例 Proportion outside the environmental space of National Park (%) | |
|---|---|---|---|---|---|
| 大于阈值 > Threshold | 小于阈值 < Threshold | ||||
| 海拔 Elevation | 4,200 m | 倭蛙 Nanorana pleskei | 7 | 64 | 90.1 |
| 青海沙蜥 Phrynocephalus vlangalii | 23 | 188 | 89.1 | ||
| 高原林蛙 Rana kukunoris | 3 | 101 | 97.1 | ||
| 西藏齿突蟾 Scutiger boulengeri | 36 | 64 | 64.0 | ||
| 年均温 Annual mean temperature | -3℃ | 倭蛙 Nanorana pleskei | 68 | 3 | 95.8 |
| 青海沙蜥 Phrynocephalus vlangalii | 192 | 19 | 91.0 | ||
| 高原林蛙 Rana kukunoris | 104 | 0 | 100 | ||
| 西藏齿突蟾 Scutiger boulengeri | 90 | 10 | 90.0 | ||
| 年降水量 Annual precipitation | 500 mm | 倭蛙 Nanorana pleskei | 69 | 2 | 97.2 |
| 青海沙蜥 Phrynocephalus vlangalii | 29 | 182 | 13.7 | ||
| 高原林蛙 Rana kukunoris | 84 | 20 | 80.8 | ||
| 西藏齿突蟾 Scutiger boulengeri | 76 | 24 | 76.0 | ||
表1 高原林蛙、倭蛙、西藏齿突蟾和青海沙蜥分布点的海拔、年均温和年降水量统计表
Table 1 The statistics of elevation, annual mean temperature and annual precipitation for the known occurrences of Rana kukunoris, Nanorana pleskei, Scutiger boulengeri, and Phrynocephalus vlangalii
| 变量 Variable | 阈值 Threshold | 物种 Species | 分布点数量 No. of occurrences | 不属于国家公园环境空间的比例 Proportion outside the environmental space of National Park (%) | |
|---|---|---|---|---|---|
| 大于阈值 > Threshold | 小于阈值 < Threshold | ||||
| 海拔 Elevation | 4,200 m | 倭蛙 Nanorana pleskei | 7 | 64 | 90.1 |
| 青海沙蜥 Phrynocephalus vlangalii | 23 | 188 | 89.1 | ||
| 高原林蛙 Rana kukunoris | 3 | 101 | 97.1 | ||
| 西藏齿突蟾 Scutiger boulengeri | 36 | 64 | 64.0 | ||
| 年均温 Annual mean temperature | -3℃ | 倭蛙 Nanorana pleskei | 68 | 3 | 95.8 |
| 青海沙蜥 Phrynocephalus vlangalii | 192 | 19 | 91.0 | ||
| 高原林蛙 Rana kukunoris | 104 | 0 | 100 | ||
| 西藏齿突蟾 Scutiger boulengeri | 90 | 10 | 90.0 | ||
| 年降水量 Annual precipitation | 500 mm | 倭蛙 Nanorana pleskei | 69 | 2 | 97.2 |
| 青海沙蜥 Phrynocephalus vlangalii | 29 | 182 | 13.7 | ||
| 高原林蛙 Rana kukunoris | 84 | 20 | 80.8 | ||
| 西藏齿突蟾 Scutiger boulengeri | 76 | 24 | 76.0 | ||
| [1] | Cai B, Li JT, Chen YY, Wang YZ (2016) Exploring the status and causes of China’s threatened reptiles through the red list assessment. Biodiversity Science, 24, 578-587. (in Chinese with English abstract) |
| [蔡波, 李家堂, 陈跃英, 王跃招 (2016) 通过红色名录评估探讨中国爬行动物受威胁现状及原因. 生物多样性, 24, 578-587.] | |
| [2] | Chen YH, Huang D, Yan SQ (2014) Discussions on public welfare, state dominance and scientificity of national park. Scientia Geographica Sinica, 34, 257-264. (in Chinese with English abstract) |
| [陈耀华, 黄丹, 颜思琦 (2014) 论国家公园的公益性、国家主导性和科学性. 地理科学, 34, 257-264.] | |
| [3] | Dudley N (2008) Guidelines for Applying Protected Area Management Categories. IUCN, Gland, Switzerland. |
| [4] | Hijmans RJ, Cameron SE, Parra JL, Jones PG, Jarvis A (2005) Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology, 25, 1965-1978. |
| [5] | Hof C, Araujo MB, Jetz W, Rahbek C (2011) Additive threats from pathogens, climate and land-use change for global amphibian diversity. Nature, 480, 516-519. |
| [6] | Jiang JP, Xie F, Zang CX, Cai L, Li C, Wang B, Li JT, Wang J, Hu JH, Wang Y, Liu JY (2016) Assessing the threat status of amphibians in China. Biodiversity Science, 24, 588-597. (in Chinese with English abstract) |
| [江建平, 谢锋, 臧春鑫, 蔡蕾, 李成, 王斌, 李家堂, 王杰, 胡军华, 王燕, 刘炯宇 (2016) 中国两栖动物受威胁现状评估. 生物多样性, 24, 588-597.] | |
| [7] | Li YM, Cohen JM, Rohr JR (2013) Review and synthesis of the effects of climate change on amphibians. Integrative Zoology, 8, 145-161. |
| [8] | Liu MC, Li DQ, Wen YM, Luan XF (2005) The ecological function analysis and evaluation of ecosystem in Sanjiangyuan region. Acta Scientiae Circumstantiae, 25, 1280-1286. (in Chinese with English abstract) |
| [刘敏超, 李迪强, 温琰茂, 栾晓峰 (2005) 三江源地区生态系统生态功能分析及其价值评估. 环境科学学报, 25, 1280-1286.] | |
| [9] | Ma KP (2014) Nature conservation is the first priority for a national park. Biodiversity Science, 22, 415-417. (in Chinese) |
| [马克平 (2014) 国家公园首先是自然保护基地. 生物多样性, 22, 415-417.] | |
| [10] | Ma KP (2017) A significant achievement in the development of national parks in China. Biodiversity Science, 25, 1031-1032. (in Chinese) |
| [马克平 (2017) 中国国家公园建设取得标志性进展. 生物多样性, 25, 1031-1032.] | |
| [11] | Tang XP (2014) On the system of national parks and the path of development in China. Biodiversity Science, 22, 427-430. (in Chinese) |
| [唐小平 (2014) 中国国家公园体制及发展思路探析. 生物多样性, 22, 427-430.] | |
| [12] | Wake DB (2012) Facing extinction in real time. Science, 335, 1052-1053. |
| [13] | Wang Y (2017) Practice and innovation for overarching institution design of China’s national park. Biodiversity Science, 25, 1037-1039. (in Chinese) |
| [王毅 (2017) 中国国家公园顶层制度设计的实践与创新. 生物多样性, 25, 1037-1039.] | |
| [14] | Wei J, Guo YM, Sun L, Jiang T, Tian XP, Sun GD (2015) Evaluation of ecological environment vulnerability for Sanjiangyuan area. Chinese Journal of Ecology, 34, 1968-1975. (in Chinese with English abstract) |
| [韦晶, 郭亚敏, 孙林, 江涛, 田信鹏, 孙光德 (2015) 三江源地区生态环境脆弱性评价. 生态学杂志, 34, 1968-1975.] | |
| [15] | Xiang BH, Zeng YX (2017) Ecotourism construction and operating mechanism in the Sanjiangyuan National Park System Pilot Area, China. Resources Science, 39, 50-60. (in Chinese with English abstract) |
| [向宝惠, 曾瑜皙 (2017) 三江源国家公园体制试点区生态旅游系统构建与运行机制探讨. 资源科学, 39, 50-60.] | |
| [16] | Xie F, Lau MWN, Stuart SN, Chanson JS, Cox NA, Fischman DL (2007) Conservation needs of amphibians in China: A review. Science in China Series C: Life Sciences, 50, 265-276. |
| [17] | Xinhua News Agency (2013) Decision of the Central Committee of the Communist Party of China on a number of significant issues in the overall deepening of reform. Qianxian, 34(12), 9-22. (in Chinese) |
| [新华社 (2013) 中共中央关于全面深化改革若干重大问题的决定. 前线, 34(12), 9-22.] | |
| [18] | Zhu CQ (2014) Perspective on development of national park system in China. Biodiversity Science, 22, 418-420. (in Chinese) |
| [朱春全 (2014) 关于建立国家公园体制的思考. 生物多样性, 22, 418-420.] | |
| [19] | Zhu CQ (2017) The objectives and missions of establishing China’s national park system. Biodiversity Science, 25, 1047-1049. (in Chinese) |
| [朱春全 (2017) 国家公园体制建设的目标与任务. 生物多样性, 25, 1047-1049.] |
| [1] | 祝晓雨, 王晨灏, 王忠君, 张玉钧. 城市绿地生物多样性研究进展与展望[J]. 生物多样性, 2025, 33(5): 25027-. |
| [2] | 王欣, 鲍风宇. 基于鸟类多样性提升的南滇池国家湿地公园生态修复效果分析[J]. 生物多样性, 2025, 33(5): 24531-. |
| [3] | 明玥, 郝培尧, 谭铃千, 郑曦. 基于城市绿色高质量发展理念的中国城市生物多样性保护与提升研究[J]. 生物多样性, 2025, 33(5): 24524-. |
| [4] | 刘咏华, 童光蓉, 余航远, 王宁宁, 任海保, 陈磊, 马克平, 米湘成. 钱江源-百山祖国家公园候选区钱江源园区冠层三维结构及光谱特征对人为干扰的响应[J]. 生物多样性, 2025, 33(4): 24174-. |
| [5] | 卢晓强, 董姗姗, 马月, 徐徐, 邱凤, 臧明月, 万雅琼, 李孪鑫, 于赐刚, 刘燕. 前沿技术在生物多样性研究中的应用现状、挑战与展望[J]. 生物多样性, 2025, 33(4): 24440-. |
| [6] | 赵维洋, 王伟, 马冰然. 其他有效的区域保护措施(OECMs)研究进展与展望[J]. 生物多样性, 2025, 33(3): 24525-. |
| [7] | 周志华, 金效华, 罗颖, 李迪强, 岳建兵, 刘芳, 何拓, 李希, 董晖, 罗鹏. 中国林草部门落实《昆明-蒙特利尔全球生物多样性框架》的机制、成效分析及建议[J]. 生物多样性, 2025, 33(3): 24487-. |
| [8] | 苏红巧, 余得光, 牟昆仑. 国家公园与国土空间规划和用途管制制度衔接路径探讨[J]. 生物多样性, 2025, 33(3): 24570-. |
| [9] | 刘立, 臧明月, 马月, 万雅琼, 胡飞龙, 卢晓强, 刘燕. 央地协同推动国家生物多样性战略和行动计划执行的措施、进展与展望[J]. 生物多样性, 2025, 33(3): 24532-. |
| [10] | 顾婧婧, 刘宜卓, 苏杨. 基层地方政府在完成《昆蒙框架》中的作用和难点: 基于《联合国气候变化框架公约》任务的比较[J]. 生物多样性, 2025, 33(3): 24585-. |
| [11] | 田志奇, 苏杨. 环境相关国际公约的中国履约模式和在《生物多样性公约》中的应用: 从完成《昆蒙框架》目标和发挥国家公园作用的角度[J]. 生物多样性, 2025, 33(3): 24593-. |
| [12] | 弋维, 艾鷖, 吴萌, 田黎明, 泽让东科. 青藏高原高寒草甸土壤古菌群落对不同放牧强度的响应[J]. 生物多样性, 2025, 33(1): 24339-. |
| [13] | 宋阳, 柳军, 何少林, 徐薇, 程琛, 刘博, 余绩庆. 我国能源企业生物多样性保护主流化管理路径[J]. 生物多样性, 2025, 33(1): 24345-. |
| [14] | 张雨琦, 文君, 张引, 李晟之. 大熊猫国家公园全民公益性评价研究: 基于利益相关者感知视角[J]. 生物多样性, 2024, 32(9): 24240-. |
| [15] | 苏荣菲, 陈睿山, 俞霖琳, 吴婧彬, 康燕. 基于红外相机调查的上海市长宁区社区生境花园生物多样性[J]. 生物多样性, 2024, 32(8): 24068-. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
备案号:京ICP备16067583号-7
Copyright © 2022 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn