
生物多样性 ›› 2018, Vol. 26 ›› Issue (11): 1133-1146. DOI: 10.17520/biods.2018098 cstr: 32101.14.biods.2018098
• 研究报告: 植物多样性 • 下一篇
吕丽莎1,2, 蔡宏宇2, 杨永3, 王志恒2,*( ), 曾辉1
), 曾辉1
收稿日期:2018-04-02
接受日期:2018-05-21
出版日期:2018-11-20
发布日期:2019-01-08
通讯作者:
王志恒
作者简介:# 共同第一作者
基金资助:
Lisha Lü1,2, Hongyu Cai2, Yong Yang3, Zhiheng Wang2,*(), Hui Zeng1
Received:2018-04-02
Accepted:2018-05-21
Online:2018-11-20
Published:2019-01-08
Contact:
Wang Zhiheng
About author:# Co-first authors
摘要:
物种多样性的大尺度空间格局是宏观生态学和生物地理学研究的核心问题之一。本文利用中国裸子植物分布数据, 结合气候、地形等环境信息, 分析了中国裸子植物物种多样性的大尺度格局及其影响因素, 比较了不同类群之间物种多样性格局和主导因子的差异, 并探讨了裸子植物在植物区系中所占比重的地理格局。结果表明, 中国裸子植物的物种多样性总体上呈现南高北低的趋势, 物种多样性在横断山区最高。在裸子植物的三个主要类群中, 松柏亚纲的物种多样性格局与整体相似, 买麻藤亚纲的多样性高值区则出现在中国西北部的干旱地区, 苏铁亚纲的分布区较为狭窄, 主要集中在南方地区。线性回归分析结果表明, 空间异质性和降水因子对中国裸子植物多样性格局的解释率最高, 末次冰期以来的气温变化、海拔高差和能量因子次之。这表明中国裸子植物物种多样性的格局受到了多种因素的影响, 其中空间异质性和降水因子影响最大。进一步分析发现, 物种多样性格局的主导因子在不同类群之间具有显著差异, 这可能反映了这些类群的进化历史以及生理适应的差异。裸子植物与被子植物的比例具有明显的空间格局: 在东部、南部气候环境优越的地区, 裸子植物与被子植物的比例低于0.06; 而在西部、北部等气候环境比较恶劣的地区, 裸子植物的比例则显著上升。回归分析表明, 能量和水分因子显著影响了裸子植物与被子植物的比例。随着能量的降低和降水的减少, 裸子植物与被子植物的比例会显著升高, 这可能是由于被子植物在温暖湿润地区具有较强竞争优势, 但裸子植物对极端环境具有更好的适应。
吕丽莎, 蔡宏宇, 杨永, 王志恒, 曾辉 (2018) 中国裸子植物的物种多样性格局及其影响因子. 生物多样性, 26, 1133-1146. DOI: 10.17520/biods.2018098.
Lisha Lü, Hongyu Cai, Yong Yang, Zhiheng Wang, Hui Zeng (2018) Geographic patterns and environmental determinants of gymnosperm species diversity in China. Biodiversity Science, 26, 1133-1146. DOI: 10.17520/biods.2018098.
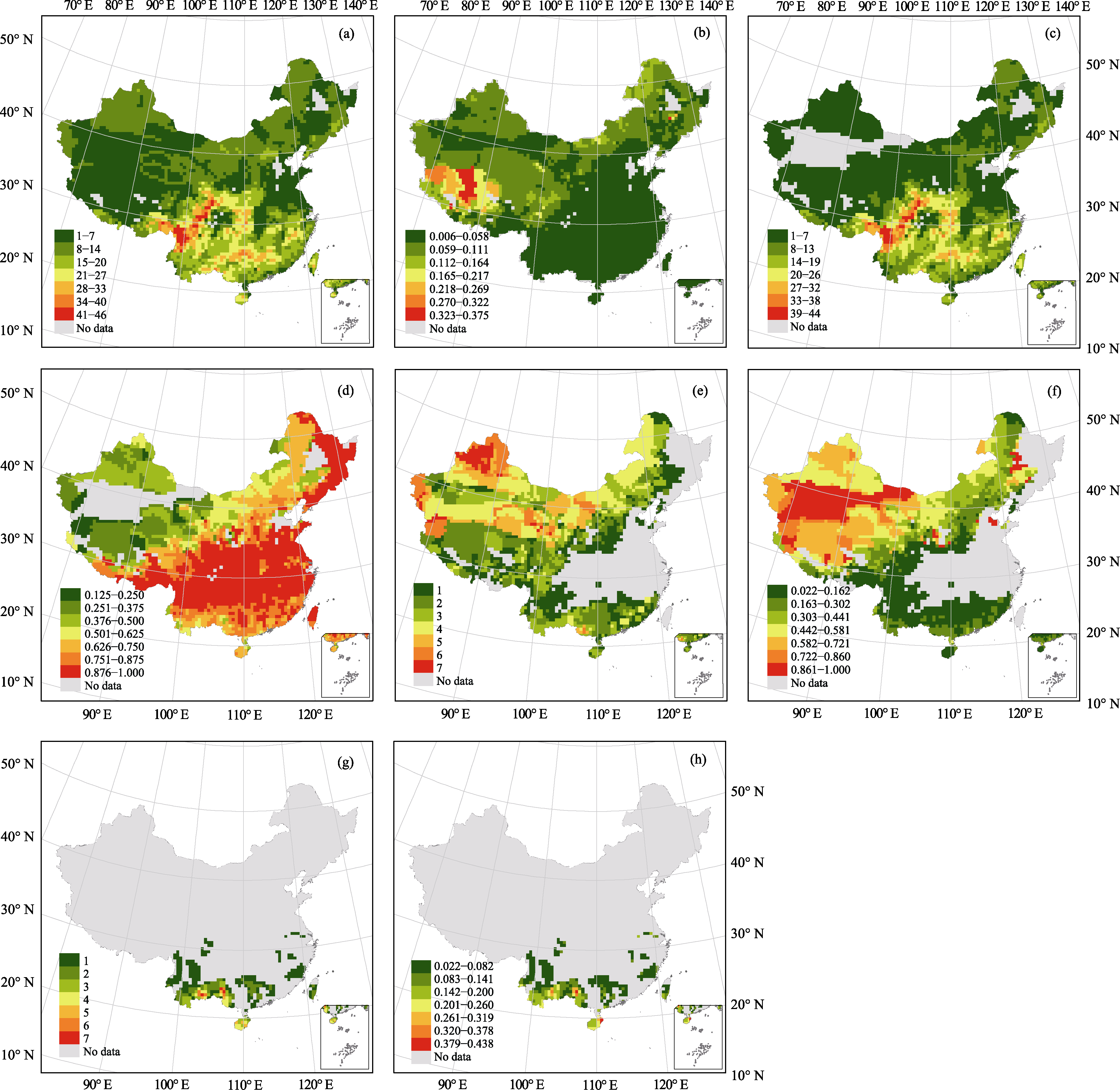
图1 中国裸子植物物种多样性格局。(a)全部裸子植物物种多样性; (b)裸子植物与被子植物的比例; (c)松柏亚纲物种多样性; (d)松柏亚纲与全部裸子植物的比例; (e)买麻藤亚纲物种多样性; (f)买麻藤亚纲与全部裸子植物的比例; (g)苏铁亚纲物种多样性; (h)苏铁亚纲与全部裸子植物的比例。
Fig. 1 Species diversity patterns of gymnosperms in China. (a) Species diversity of all gymnosperms; (b) Ratio of gymnosperms to angiosperms; (c) Species diversity of Pinidae; (d) Ratio of Pinidae to all gymnosperms; (e) Species diversity of Gnetidae; (f) Ratio of Gnetidae to all gymnosperms; (g) Species diversity of Cycadidae; (h) Ratio of Cycadidae to all gymnosperms.
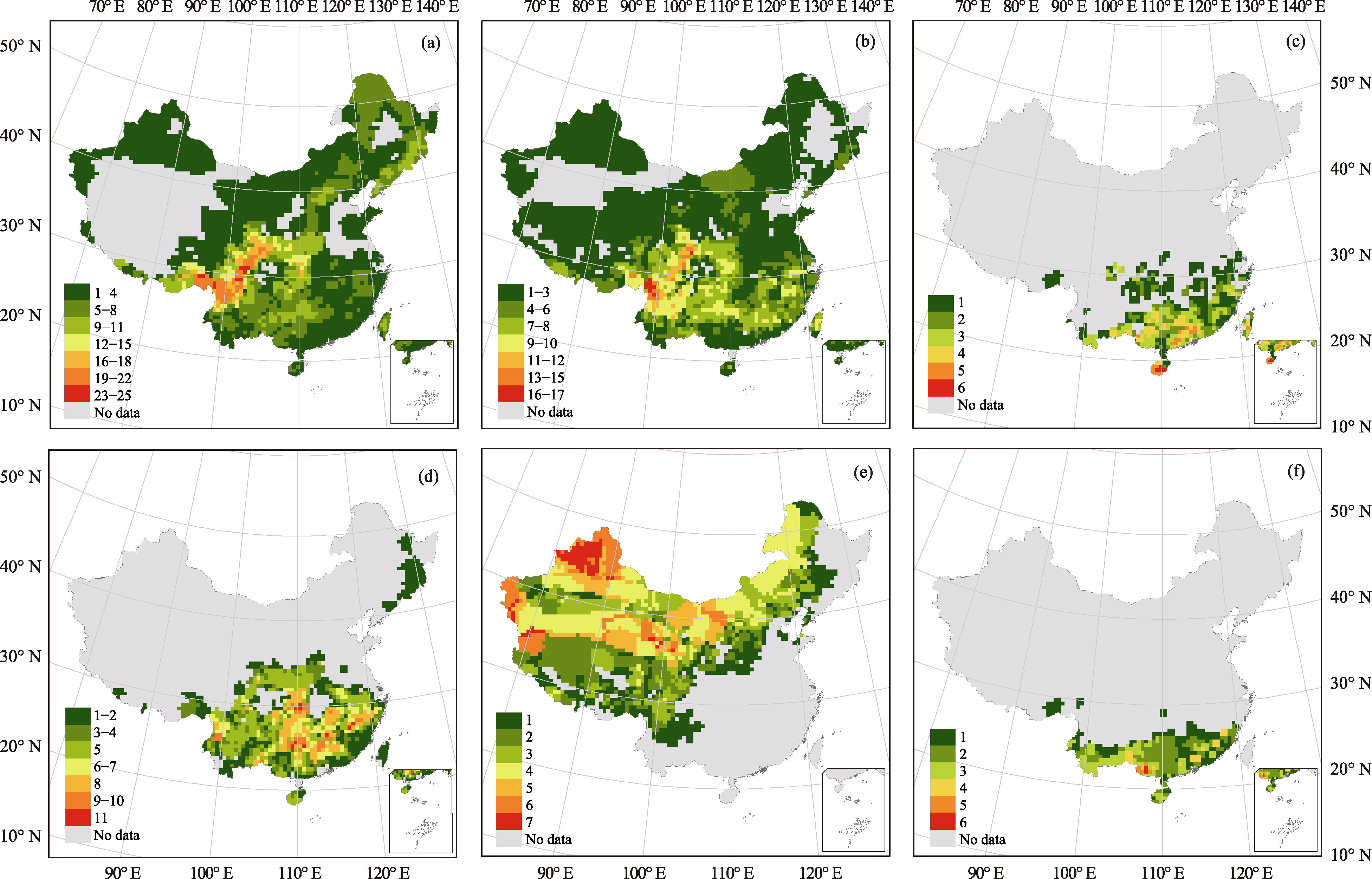
图2 中国裸子植物各科物种多样性格局。(a)松科; (b)柏科; (c)罗汉松科; (d)红豆杉科; (e)麻黄科; (f)买麻藤科。
Fig. 2 Species diversity patterns of gymnosperms in China. (a) Pinaceae; (b) Cupressaceae; (c) Podocarpaceae; (d) Taxaceae; (e) Ephedraceae; (f) Gnetaceae.
| 环境变量 Environmental determinants | 全部All gymno- sperms | 比例 Ratio | 松柏亚纲 Pinidae | 买麻藤亚纲 Gnetidae | 苏铁亚纲 Cycadidae | 松科 Pinaceae | 柏科 Cupres- saceae | 罗汉松科 Podocar- paceae | 红豆杉科 Taxaceae | 麻黄科 Ephed- raceae | 买麻藤科 Gnetaceae |
|---|---|---|---|---|---|---|---|---|---|---|---|
| MAT | 0.278* | -0.826* | 0.501* | -0.266* | 0.455* | 0.115 | 0.376* | 0.421* | 0.352* | -0.198* | 0.342* |
| MTCQ | 0.338* | -0.805* | 0.504* | -0.378* | 0.535* | 0.207* | 0.417* | 0.476* | 0.366* | -0.397* | 0.394* |
| WI | 0.232 | -0.790* | 0.429* | -0.191* | 0.467* | 0.024 | 0.277* | 0.429* | 0.230* | -0.084 | 0.346* |
| PET | 0.212 | -0.778* | 0.426* | -0.182* | 0.426* | 0.007 | 0.274* | 0.395* | 0.233* | -0.076 | 0.274* |
| MAP | 0.478* | -0.706* | 0.615* | -0.423* | 0.044 | 0.365* | 0.449* | 0.286* | 0.313* | -0.520* | 0.105 |
| PCQ | 0.331* | -0.117* | 0.416* | -0.288* | -0.169* | 0.113 | 0.323* | 0.087 | 0.303* | -0.291* | -0.117* |
| MI | 0.491* | -0.469* | 0.617* | -0.465* | -0.222* | 0.529* | 0.485* | 0.028 | 0.230* | -0.467* | -0.132* |
| AET | 0.391* | -0.748* | 0.555* | -0.459* | 0.403* | 0.336* | 0.350* | 0.392* | 0.246* | -0.547* | 0.291* |
| ER | 0.484* | 0.080 | 0.333* | 0.006 | -0.164* | 0.335* | 0.321* | 0.066 | 0.187* | 0.012 | -0.088 |
| TR | 0.462* | 0.089 | 0.313* | -0.015 | -0.140* | 0.334* | 0.306* | 0.071 | 0.163* | -0.014 | -0.067 |
| PR | 0.556* | -0.431* | 0.614* | -0.228* | 0.092 | 0.412* | 0.486* | 0.068 | 0.166* | -0.185* | -0.139* |
| MATano | -0.400* | 0.517* | -0.407* | 0.047 | -0.546* | -0.042 | -0.399* | -0.551* | -0.376* | -0.120* | -0.451* |
| MAPano | 0.001 | -0.099 | 0.020 | -0.033 | 0.170* | -0.079* | -0.112* | 0.501* | -0.113* | 0.132* | 0.192* |
表1 中国全部裸子植物及不同类群的物种多样性和裸子植物与被子植物比例与环境因子的单因素相关关系(R)
Table 1 Bivariate correlations (R) between species diversity of all gymnosperm species, species diversity of gymnosperms in different clades, ratio of all gymnosperms to angiosperms and environmental determinants in China
| 环境变量 Environmental determinants | 全部All gymno- sperms | 比例 Ratio | 松柏亚纲 Pinidae | 买麻藤亚纲 Gnetidae | 苏铁亚纲 Cycadidae | 松科 Pinaceae | 柏科 Cupres- saceae | 罗汉松科 Podocar- paceae | 红豆杉科 Taxaceae | 麻黄科 Ephed- raceae | 买麻藤科 Gnetaceae |
|---|---|---|---|---|---|---|---|---|---|---|---|
| MAT | 0.278* | -0.826* | 0.501* | -0.266* | 0.455* | 0.115 | 0.376* | 0.421* | 0.352* | -0.198* | 0.342* |
| MTCQ | 0.338* | -0.805* | 0.504* | -0.378* | 0.535* | 0.207* | 0.417* | 0.476* | 0.366* | -0.397* | 0.394* |
| WI | 0.232 | -0.790* | 0.429* | -0.191* | 0.467* | 0.024 | 0.277* | 0.429* | 0.230* | -0.084 | 0.346* |
| PET | 0.212 | -0.778* | 0.426* | -0.182* | 0.426* | 0.007 | 0.274* | 0.395* | 0.233* | -0.076 | 0.274* |
| MAP | 0.478* | -0.706* | 0.615* | -0.423* | 0.044 | 0.365* | 0.449* | 0.286* | 0.313* | -0.520* | 0.105 |
| PCQ | 0.331* | -0.117* | 0.416* | -0.288* | -0.169* | 0.113 | 0.323* | 0.087 | 0.303* | -0.291* | -0.117* |
| MI | 0.491* | -0.469* | 0.617* | -0.465* | -0.222* | 0.529* | 0.485* | 0.028 | 0.230* | -0.467* | -0.132* |
| AET | 0.391* | -0.748* | 0.555* | -0.459* | 0.403* | 0.336* | 0.350* | 0.392* | 0.246* | -0.547* | 0.291* |
| ER | 0.484* | 0.080 | 0.333* | 0.006 | -0.164* | 0.335* | 0.321* | 0.066 | 0.187* | 0.012 | -0.088 |
| TR | 0.462* | 0.089 | 0.313* | -0.015 | -0.140* | 0.334* | 0.306* | 0.071 | 0.163* | -0.014 | -0.067 |
| PR | 0.556* | -0.431* | 0.614* | -0.228* | 0.092 | 0.412* | 0.486* | 0.068 | 0.166* | -0.185* | -0.139* |
| MATano | -0.400* | 0.517* | -0.407* | 0.047 | -0.546* | -0.042 | -0.399* | -0.551* | -0.376* | -0.120* | -0.451* |
| MAPano | 0.001 | -0.099 | 0.020 | -0.033 | 0.170* | -0.079* | -0.112* | 0.501* | -0.113* | 0.132* | 0.192* |
| 校正后模型解释率 Adjust R2 | 能量 Energy factors | 水分 Precipitation factors | 异质性 Environmental heterogeneity | 过去气候变化 Climate anomaly | ||
|---|---|---|---|---|---|---|
| 裸子植物 All gymnosperms | 变量 Variables | 0.436 | PET | MI | ER | MATano |
| 偏决定系数 Partial R2 | 0.037 | 0.081 | 0.171 | 0.010 | ||
| 标准化系数 Standardized coefficient | 0.228 | 0.255 | 0.464 | -0.107 | ||
| 裸子植物/ 被子植物 Ratio of gymnosperms to angiosperms | 变量 Variables | 0.734 | MAT | AET | PR | MAPano |
| 偏决定系数 Partial R2 | 0.413 | 0.061 | 0.005 | 0.010 | ||
| 标准化系数 Standardized coefficient | -0.635 | -0.257 | -0.055 | 0.054 | ||
| 松柏亚纲 Pinidae | 变量 Variables | 0.526 | MAT | MI | ER | MATano |
| 偏决定系数 Partial R2 | 0.135 | 0.172 | 0.119 | 0.001 | ||
| 标准化系数 Standardized coefficient | 0.415 | 0.387 | 0.335 | 0.038 | ||
| 松科 Pinaceae | 变量 Variables | 0.331 | MAT | MI | ER | MATano |
| 偏决定系数 Partial R2 | 0.008 | 0.194 | 0.057 | 0.033 | ||
| 标准化系数 Standardized coefficient | 0.122 | 0.480 | 0.277 | 0.207 | ||
| 柏科 Cupressaceae | 变量 Variables | 0.360 | MAT | MI | ER | MAPano |
| 偏决定系数 Partial R2 | 0.107 | 0.080 | 0.084 | 0.026 | ||
| 标准化系数 Standardized coefficient | 0.314 | 0.290 | 0.311 | -0.126 | ||
| 罗汉松科 Podocarpaceae | 变量 Variables | 0.370 | PET | MI | TR | MATano |
| 偏决定系数 Partial R2 | 0.034 | 0.013 | 0.052 | 0.144 | ||
| 标准化系数 Standardized coefficient | 0.366 | 0.169 | 0.408 | -0.538 | ||
| 红豆杉科 Taxaceae | 变量 Variables | 0.282 | MAT | PCQ | ER | MAPano |
| 偏决定系数 Partial R2 | 0.087 | 0.059 | 0.099 | 0.046 | ||
| 标准化系数 Standardized coefficient | 0.454 | 0.220 | 0.498 | -0.127 | ||
| 买麻藤亚纲 Gnetidae | 变量 Variables | 0.364 | MTCQ | MI | PR | MATano |
| 偏决定系数 Partial R2 | 0.162 | 0.128 | 0.019 | 0.119 | ||
| 标准化系数 Standardized coefficient | -0.574 | -0.468 | 0.160 | -0.424 | ||
| 买麻藤科 Gnetaceae | 变量 Variables | 0.314 | MTCQ | PCQ | ER | MATano |
| 偏决定系数 Partial R2 | 0.034 | 0.018 | 0.074 | 0.178 | ||
| 标准化系数 Standardized coefficient | 0.551 | -0.124 | 0.569 | -0.704 | ||
| 麻黄科 Ephedraceae | 变量 Variables | 0.431 | MTCQ | AET | PR | MATano |
| 偏决定系数 Partial R2 | 0.141 | 0.186 | 0.022 | 0.062 | ||
| 标准化系数 Standardized coefficient | -0.571 | -0.830 | 0.156 | -0.296 | ||
| 苏铁亚纲 Cyacadidae | 变量 Variables | 0.416 | MTCQ | PCQ | PR | MATano |
| 偏决定系数 Partial R2 | 0.028 | 0.096 | 0.034 | 0.151 | ||
| 标准化系数 Standardized coefficient | 0.478 | -0.221 | 0.334 | -0.663 |
表2 中国裸子植物及主要类群物种多样性与环境因子的最优线性模型(P < 0.01)
Table 2 Environmental determinants of species diversity for all gymnosperm species, species diversity of gymnosperms in different clades and ratio of all gymnosperms to angiosperms in China from best-fit explanatory models
| 校正后模型解释率 Adjust R2 | 能量 Energy factors | 水分 Precipitation factors | 异质性 Environmental heterogeneity | 过去气候变化 Climate anomaly | ||
|---|---|---|---|---|---|---|
| 裸子植物 All gymnosperms | 变量 Variables | 0.436 | PET | MI | ER | MATano |
| 偏决定系数 Partial R2 | 0.037 | 0.081 | 0.171 | 0.010 | ||
| 标准化系数 Standardized coefficient | 0.228 | 0.255 | 0.464 | -0.107 | ||
| 裸子植物/ 被子植物 Ratio of gymnosperms to angiosperms | 变量 Variables | 0.734 | MAT | AET | PR | MAPano |
| 偏决定系数 Partial R2 | 0.413 | 0.061 | 0.005 | 0.010 | ||
| 标准化系数 Standardized coefficient | -0.635 | -0.257 | -0.055 | 0.054 | ||
| 松柏亚纲 Pinidae | 变量 Variables | 0.526 | MAT | MI | ER | MATano |
| 偏决定系数 Partial R2 | 0.135 | 0.172 | 0.119 | 0.001 | ||
| 标准化系数 Standardized coefficient | 0.415 | 0.387 | 0.335 | 0.038 | ||
| 松科 Pinaceae | 变量 Variables | 0.331 | MAT | MI | ER | MATano |
| 偏决定系数 Partial R2 | 0.008 | 0.194 | 0.057 | 0.033 | ||
| 标准化系数 Standardized coefficient | 0.122 | 0.480 | 0.277 | 0.207 | ||
| 柏科 Cupressaceae | 变量 Variables | 0.360 | MAT | MI | ER | MAPano |
| 偏决定系数 Partial R2 | 0.107 | 0.080 | 0.084 | 0.026 | ||
| 标准化系数 Standardized coefficient | 0.314 | 0.290 | 0.311 | -0.126 | ||
| 罗汉松科 Podocarpaceae | 变量 Variables | 0.370 | PET | MI | TR | MATano |
| 偏决定系数 Partial R2 | 0.034 | 0.013 | 0.052 | 0.144 | ||
| 标准化系数 Standardized coefficient | 0.366 | 0.169 | 0.408 | -0.538 | ||
| 红豆杉科 Taxaceae | 变量 Variables | 0.282 | MAT | PCQ | ER | MAPano |
| 偏决定系数 Partial R2 | 0.087 | 0.059 | 0.099 | 0.046 | ||
| 标准化系数 Standardized coefficient | 0.454 | 0.220 | 0.498 | -0.127 | ||
| 买麻藤亚纲 Gnetidae | 变量 Variables | 0.364 | MTCQ | MI | PR | MATano |
| 偏决定系数 Partial R2 | 0.162 | 0.128 | 0.019 | 0.119 | ||
| 标准化系数 Standardized coefficient | -0.574 | -0.468 | 0.160 | -0.424 | ||
| 买麻藤科 Gnetaceae | 变量 Variables | 0.314 | MTCQ | PCQ | ER | MATano |
| 偏决定系数 Partial R2 | 0.034 | 0.018 | 0.074 | 0.178 | ||
| 标准化系数 Standardized coefficient | 0.551 | -0.124 | 0.569 | -0.704 | ||
| 麻黄科 Ephedraceae | 变量 Variables | 0.431 | MTCQ | AET | PR | MATano |
| 偏决定系数 Partial R2 | 0.141 | 0.186 | 0.022 | 0.062 | ||
| 标准化系数 Standardized coefficient | -0.571 | -0.830 | 0.156 | -0.296 | ||
| 苏铁亚纲 Cyacadidae | 变量 Variables | 0.416 | MTCQ | PCQ | PR | MATano |
| 偏决定系数 Partial R2 | 0.028 | 0.096 | 0.034 | 0.151 | ||
| 标准化系数 Standardized coefficient | 0.478 | -0.221 | 0.334 | -0.663 |
| [1] | Araújo MB, Nogués-Bravo D, Diniz-Filho JAF, Haywood AM, Valdes PJ, Rahbek C (2008) Quaternary climate changes explain diversity among reptiles and amphibians. Ecography, 31, 8-15. |
| [2] | Augusto L, Davies TJ, Delzon S, de Schrijver A (2014) The enigma of the rise of angiosperms: Can we untie the knot? Ecology Letters, 17, 1326-1338. |
| [3] | Becker P (2000) Competition in the regeneration niche between conifers and angiosperms: Bond’s slow seedling hypothesis. Functional Ecology, 14, 401-412. |
| [4] | Biffin E, Brodribb TJ, Hill RS, Thomas P, Lowe AJ (2012) Leaf evolution in Southern Hemisphere conifers tracks the angiosperm ecological radiation. Proceedings of the Royal Society B: Biological Sciences, 279, 341-348. |
| [5] | Bond W (1989) The tortoise and the hare: Ecology of angiosperm dominance and gymnosperm persistence. Biological Journal of the Linnean Society, 36, 227-249. |
| [6] | Brodribb TJ, Feild TS (2008) Evolutionary significance of a flat-leaved Pinus in Vietnamese rainforest. New Phytologist, 178, 201-209. |
| [7] | Brodribb TJ, Pittermann J, Coomes DA (2012) Elegance versus speed: Examining the competition between conifer and angiosperm trees. International Journal of Plant Sciences, 173, 673-694. |
| [8] | Chave J, Coomes D, Jansen S, Lewis SL, Swenson NG, Zanne AE (2009) Towards a worldwide wood economics spectrum. Ecology Letters, 12, 351-366. |
| [9] | Chen S, Mao L, Zhang J, Zhou K, Gao J (2014) Environmental determinants of geographic butterfly richness pattern in eastern China. Biodiversity & Conservation, 23, 1453-1467. |
| [10] | Condamine FL, Nagalingum NS, Marshall CR, Morlon H (2015) Origin and diversification of living cycads: A cautionary tale on the impact of the branching process prior in Bayesian molecular dating. BMC Evolutionary Biology, 15, 65-65. |
| [11] | Coomes DA, Grubb PJ (2000) Impacts of root competition in forests and woodlands: A theoretical framework and review of experiments. Ecological Monographs, 70, 171-207. |
| [12] | Cun YZ, Wang XQ (2010) Plant recolonization in the Himalaya from the southeastern Qinghai-Tibetan Plateau: Geographical isolation contributed to high population differentiation. Molecular Phylogenetics & Evolution, 56, 972-982. |
| [13] | Currie DJ (1991) Energy and large-scale patterns of animal- and plant-species richness. The American Naturalist, 137, 27-49. |
| [14] | Currie DJ, Mittelbach GG, Cornell HV, Field R, Guegan J, Hawkins BA, Kaufman DM, Kerr JT, Oberdorff T, Obrien EM, Turner JRG (2004) Predictions and tests of climate-based hypotheses of broad-scale variation in taxonomic richness. Ecology Letters, 7, 1121-1134. |
| [15] | Dutilleul P, Clifford P, Richardson S, Hemon D (1993) Modifying the t test for assessing the correlation between two spatial processes. Biometrics, 49, 305-314. |
| [16] | Fang JY, Yoda K (1990) Climate and vegetation in China. III. Water balance and distribution of vegetation. Ecological Research, 5, 9-23. |
| [17] | Fang JY, Wang ZH, Tang ZY (2011) Atlas of Woody Plants in China: Distribution and Climate. Higher Education Press, Beijing & Springer, Berlin. |
| [18] | Farjon A (2010) A Handbook of the World’s Conifers. Brill Academic Publishers, Netherlands. |
| [19] | Fjeldså J, Bowie RCK, Rahbek C (2012) The role of mountain ranges in the diversification of birds. Annual Review of Ecology, Evolution, and Systematics, 43, 249-265. |
| [20] | Fragnière Y, Bétrisey S, Cardinaux L, Stoffel M, Kozlowski G (2015) Fighting their last stand? A global analysis of the distribution and conservation status of gymnosperms. Journal of Biogeography, 42, 809-820. |
| [21] | Gao JF, Ma KM, Feng ZW, Qi J, Feng Y (2009) Coupling effects of altitude and human disturbance on landscape and plant diversity in the vicinity of mountain villages of Beijing, China. Acta Ecologica Sinica, 29, 56-61. |
| [22] | Gear AJ, Huntley B (1991) Rapid changes in the range limits of Scots Pine 4000 years ago. Science, 251, 544-547. |
| [23] | Gerrienne P, Meyer-Berthaud B, Fairon-Demaret M, Streel M, Steemans P (2004) Runcaria, a Middle Devonian seed plant precursor. Science, 306, 856-858. |
| [24] | Hacke UG, Sperry JS, Pockman WT, Davis SD (2001) Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure. Oecologia, 126, 457-461. |
| [25] | Hawkins BA, Field R, Cornell HV, Currie DJ, Guegan J, Kaufman DM, Kerr JT, Mittelbach GG, Oberdorff T, O’brien EM, Porter EE, Turner JRG (2003) Energy, water, and broad-scale geographic patterns of species richness. Ecology, 84, 3105-3117. |
| [26] | Hill K (2012) The Cycad Pages. . (accessed on 2018-03-01 |
| [27] | Hong SK (1998) Changes in landscape patterns and vegetation process in the Far-Eastern cultural landscapes: Human activity on pine-dominated secondary vegetations in Korea and Japan. Phytocoenologia, 28, 45-66. |
| [28] | Hsieh L (1992) Origin and distribution of Ginkgo biloba. The Forestry Chronicle, 68, 612-613. |
| [29] | Hu HH (1998) How Metasequoia, the “living fossil”, was discovered in China. Arnoldia, 58, 4-7. |
| [30] | Jetz W, Rahbek C (2002) Geographic range size and determinants of avian species richness. Science, 297, 1548. |
| [31] | Ji RS (2007) Forest changes in Inner Mongolia—Forest changes during geological times. Inner Mongolia Forestry Investigation and Design, (S1), 1-94, 106-108. |
| (in Chinese) [纪仁生 (2007) 内蒙古森林变迁——地史时期森林的变迁. 内蒙古林业调查设计, (S1), 1-94, 106-108.] | |
| [32] | Kozlowski G, Stoffel M, Betrisey S, Cardinaux L, Mota M (2015) Hydrophobia of gymnosperms: Myth or reality? A global analysis. Ecohydrology, 8, 105-112. |
| [33] | Kreft H, Jetz W (2007) Global patterns and determinants of vascular plant diversity. Proceedings of the National Academy of Sciences, USA, 104, 5925-5930. |
| [34] | Latham RE, Ricklefs RE (1993) Global patterns of tree species richness in moist forests: Energy-diversity theory does not account for variation in species richness. Oikos, 67, 325-333. |
| [35] | Lennon JJ (2000) Red-shifts and red herrings in geographical ecology. Ecography, 23, 101-113. |
| [36] | Leslie AB, Beaulieu JM, Rai HS, Crane PR, Donoghue MJ, Mathews S (2012) Hemisphere-scale differences in conifer evolutionary dynamics. Proceedings of the National Academy of Sciences, USA, 109, 16217-16221. |
| [37] | Li G, Shen ZH, Ying TS, Fang JY (2009) The spatial pattern of species richness and diversity centers of gymnosperm in China. Biodiversity Science, 17, 272-279. |
| (in Chinese with English abstract) [李果, 沈泽昊, 应俊生, 方精云 (2009) 中国裸子植物物种丰富度空间格局与多样性中心. 生物多样性, 17, 272-279.] | |
| [38] | Li ZL (1981) Morphology and structure of drought vegetation. Biology Journal, (4), 9-12. (in Chinese) |
| [李正理 (1981) 旱生植物的形态和结构. 生物学通报, (4), 9-12.] | |
| [39] | Lin X, Wang ZH, Tang ZY, Zhao SQ, Fang JY (2009) Geographic patterns and environmental correlates of terrestrial mammal species richness in China. Biodiversity Science, 17, 652-663. (in Chinese with English abstract) |
| [林鑫, 王志恒, 唐志尧, 赵淑清, 方精云 (2009) 中国陆栖哺乳动物物种丰富度的地理格局及其与环境因子的关系. 生物多样性, 17, 652-663.] | |
| [40] | López-Pujol J, Zhang FM, Sun HQ, Ying TS, Ge S (2011) Mountains of southern China as “plant museums” and “plant cradles”: Evolutionary and conservation insights. Mountain Research and Development, 31, 261-269. |
| [41] | Maurer BA, Rosenzweig ML (1996) Species Diversity in Space and Time. Ecology, 77, 1314. |
| [42] | McCain CM (2009) Global analysis of bird elevational diversity. Global Ecology and Biogeography, 18, 346-360. |
| [43] | Mou FJ, Li JP, Chen LP, Li YG (2016) The progress on the relationship between the morphological and anatomical character and the resistant. Journal of Fujian Forestry Science & Technology, 43, 237-243. (in Chinese with English abstract) |
| [牟凤娟, 李军萍, 陈丽萍, 李一果 (2016) 裸子植物形态解剖结构特征与抗旱性研究进展. 福建林业科技, 43, 237-243.] | |
| [44] | Nagalingum NS, Marshall CR, Quental TB, Rai HS, Little DP, Mathews S (2011) Recent synchronous radiation of a living fossil. Science, 334, 796-799. |
| [45] | O’Brien EM (1998) Water-energy dynamics, climate, and prediction of woody plant species richness: An interim general model. Journal of Biogeography, 25, 379-398. |
| [46] | Osorio F, Vallejos R, Cuevas F (2016) SpatialPack: Computing the association between two spatial processes. arXiv preprint arXiv:1611.05289. |
| [47] | Palmer MW (1994) Variation in species richness: Towards a unification of hypotheses. Folia Geobotanica et Phytotaxonomica, 29, 511. |
| [48] | Pittermann J, Sperry JS, Wheeler JK, Hacke UG, Sikkema EH (2006) Mechanical reinforcement of tracheids compromises the hydraulic efficiency of conifer xylem. Plant Cell & Environment, 29, 1618. |
| [49] | Pittermann J, Stuart SA, Dawson TE, Moreau A (2012) Cenozoic climate change shaped the evolutionary ecophysiology of the Cupressaceae conifers. Proceedings of the National Academy of Sciences, USA, 109, 9647-9652. |
| [50] | Qian H, Jin Y, Ricklefs RE (2017) Phylogenetic diversity anomaly in angiosperms between eastern Asia and eastern North America. Proceedings of the National Academy of Sciences, USA, 114, 11452-11457. |
| [51] | Qin AL, Wang MM, Cun YZ, Yang FS, Wang SS, Ran JH, Wang XQ (2013) Phylogeographic evidence for a link of species divergence of Ephedra in the Qinghai-Tibetan Plateau and adjacent regions to the Miocene Asian Aridification. PLoS ONE, 8, e56243. |
| [52] | Ran JH, Wei XX, Wang XQ (2006) Molecular phylogeny and biogeography of Picea (Pinaceae): Implications for phylogeographical studies using cytoplasmic haplotypes. Molecular Phylogenetics and Evolution, 41, 405-419. |
| [53] | R Core Team (2016) R: A Language and Environment for Statistical Computing. . (accessed on 2018-01-01) |
| [54] | Rosenzweig ML (1995) Species Diversity in Space and Time. Cambridge University Press, Cambridge. |
| [55] | Rueda M, Godoy O, Hawkins BA (2017) Spatial and evolutionary parallelism between shade and drought tolerance explains the distributions of conifers in the conterminous United States. Global Ecology and Biogeography, 26, 31-42. |
| [56] | Sandel B, Arge L, Dalsgaard B, Davies RG, Gaston KJ, Sutherland WJ, Svenning J (2011) The influence of Late Quaternary climate-change velocity on species endemism. Science, 334, 660-664. |
| [57] | Sperry JS, Hacke UG, Pittermann J (2006) Size and function in conifer tracheids and angiosperm vessels. American Journal of Botany, 93, 1490-1500. |
| [58] | Stein A, Gerstner K, Kreft H (2014) Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecology Letters, 17, 866-880. |
| [59] | Stein A, Beck J, Meyer C, Waldmann E, Weigelt P, Kreft H (2015) Differential effects of environmental heterogeneity on global mammal species richness. Global Ecology and Biogeography, 24, 1072-1083. |
| [60] | Sun H (2002) Evolution of Arctic-Tertiary flora in Himalayan-Hengduan Mountains. Acta Botanica Yunnanica, 24, 671-688. (in Chinese with English abstract) |
| [孙航 (2002) 北极—第三纪成分在喜马拉雅—横断山的发展及演化. 云南植物研究, 24, 671-688.] | |
| [61] | Svenning JC, Skov F (2007) Ice age legacies in the geographical distribution of tree species richness in Europe. Global Ecology and Biogeography, 16, 234-245. |
| [62] | Taylor EL, Taylor TN, Krings M (2009) Paleobotany: The Biology and Evolution of Fossil Plants. Academic Press, Waltham. |
| [63] | Thornthwaite CW, Hare FK (1955) Climatic classification in forest. Unasylva, 9, 51-59. |
| [64] | Wan T, Liu ZM, Li LF, Leitch AR, Leitch IJ, Lohaus R, Liu ZJ, Xin HP, Gong YB, Liu Y, Wang WC, Chen LY, Yang Y, Kelly LJ, Yang J, Huang JL, Li Z, Liu P, Zhang L, Liu HM, Wang H, Deng SH, Liu M, Li J, Ma L, Liu Y, Lei Y, Xu W, Wu LQ, Liu F, Ma Q, Yu XR, Jiang Z, Zhang GQ, Li SH, Li RQ, Zhang SZ, Wang QF, de Peer YV, Zhang JB, Wang XM (2018) A genome for gnetophytes and early evolution of seed plants. Nature Plants, 4, 82-89. |
| [65] | Wang XQ, Han Y, Hong DY (1998) A molecular systematic study of Cathaya, a relic genus of the Pinaceae in China. Plant Systematics and Evolution, 213, 165-172. |
| [66] | Wang ZH, Fang JY, Tang ZY, Lin X (2011) Patterns, determinants and models of woody plant diversity in China. Proceedings of the Royal Society B: Biological Sciences, 278, 2122-2132. |
| [67] | Wang ZH, Fang JY, Tang ZY, Shi L (2012a) Geographical patterns in the beta diversity of China’s woody plants: The influence of space, environment and range size. Ecography, 35, 1092-1102. |
| [68] | Wang ZH, Fang JY, Tang ZY, Lin X (2012b) Relative role of contemporary environment versus history in shaping diversity patterns of China’s woody plants. Ecography, 35, 1124-1133. |
| [69] | Wang ZH, Tang ZY, Fang JY (2009) The species-energy hypothesis as a mechanism for species richness patterns. Biodiversity Science 17, 613-624. (in Chinese with English abstract) |
| [王志恒, 唐志尧, 方精云 (2009) 物种多样性地理格局的能量假说. 生物多样性, 17, 613-624.] | |
| [70] | Wei XX, Wang XQ (2004) Recolonization and radiation in Larix (Pinaceae): Evidence from nuclear ribosomal DNA paralogues. Molecular Ecology, 13, 3115-3123. |
| [71] | Wu Z, Raven P (1999) Flora of China, Vol. 4. Science Press, Beijing & Missouri Botanical Garden Press, St. Louis. |
| [72] | Xiang XG, Cao M, Zhou ZK (2006) Fossil history and modern distribution of the genus Abies (Pinaceae). Acta Botanica Yunnanica, 28, 439-452. (in Chinese with English abstract) |
| [向小果, 曹明, 周浙昆 (2006) 松科冷杉属植物的化石历史和现代分布. 云南植物研究, 28, 439-452.] | |
| [73] | Xing YW, Ree RH (2017) Uplift-driven diversification in the Hengduan Mountains, a temperate biodiversity hotspot. Proceedings of the National Academy of Sciences, USA, 114, E3444. |
| [74] | Yang Y, Wang ZH, Xu XT (2017) Taxonomy and Distribution of Global Gymnosperms. Shanghai Scientific and Technical Publishers, Shanghai. (in Chinese) |
| [杨永, 王志恒, 徐晓婷 (2017) 世界裸子植物的分类和地理分布. 上海科学技术出版社, 上海.] | |
| [75] | Yeaman S, Hodgins KA, Lotterhos KE, Suren H, Nadeau S, Degner JC, Nurkowski KA, Smets P, Wang T, Gray LK, Liepe KJ, Hamann A, Holliday J, Whitlock MC, Rieseberg LH, Aitken SN (2016) Convergent local adaptation to climate in distantly related conifers. Science, 353, 1431-1433. |
| [76] | Ye JF, Chen ZD, Liu B, Qin HN, Yang Y (2012) Disjunct distribution of vascular plants between southwestern area and Taiwan area in China: Disjunct distribution of vascular plants between southwestern area and Taiwan area in China. Biodiversity Science, 20, 482-494. (in Chinese with English abstract) |
| [叶建飞, 陈之端, 刘冰, 覃海宁, 杨永 (2012) 中国西南与台湾地区维管植物的间断分布格局及形成机制. 生物多样性, 20, 482-494.] | |
| [77] | Ying J, Chen M, Zhang H (2003) Atlas of the Gymnosperms of China. China Science & Technology Press, Beijing. |
| [78] | Zhang ZJ, Yan YJ, Tian Y, Li JS, He JS, Tang ZY (2015) Distribution and conservation of orchid species richness in China. Biological Conservation, 181, 64-72. |
| [1] | 吴晓晴 张美惠 葛苏婷 李漫淑 宋坤 沈国春 达良俊 张健. 上海近自然林重建过程中木本植物物种多样性与地上生物量的时空动态——以闵行区生态岛为例[J]. 生物多样性, 2025, 33(5): 24444-. |
| [2] | 王太, 宋福俊, 张永胜, 娄忠玉, 张艳萍, 杜岩岩. 河西走廊内陆河水系鱼类多样性及资源现状[J]. 生物多样性, 2025, 33(4): 24387-. |
| [3] | 张晶晶, 黄文彬, 陈奕廷, 杨泽鹏, 柯伟业, 彭昭杰, 魏世超, 张志伟, 胡怡思, 余文华, 周文良. 广东南澎列岛海洋生态国家级自然保护区造礁石珊瑚多样性及分布特征[J]. 生物多样性, 2025, 33(4): 24424-. |
| [4] | 尚华丹, 张楚晴, 王梅, 裴文娅, 李国宏, 王鸿斌. 中国杨树害虫物种多样性及其地理分布[J]. 生物多样性, 2025, 33(2): 24370-. |
| [5] | 吴昱萱, 王平, 胡晓生, 丁一, 彭甜恬, 植秋滢, 巴德木其其格, 李文杰, 关潇, 李俊生. 呼伦贝尔草地退化现状评估与植被特征变化[J]. 生物多样性, 2025, 33(2): 24118-. |
| [6] | 陈自宏, 张翼飞, 陈凯, 陈见影, 徐玲. 高黎贡山南段昆虫病原真菌物种多样性及影响因素[J]. 生物多样性, 2025, 33(1): 24228-. |
| [7] | 谭珂, 宁瑶, 王仁芬, 王晴, 梁丹萍, 辛子兵, 温放. 中国苦苣苔科植物名录与地理分布数据集[J]. 生物多样性, 2025, 33(1): 23275-. |
| [8] | 韩佳楠, 苏杨, 李霏, 刘君妍, 赵依林, 李琳, 赵建成, 梁红柱, 李敏. 河北省苔藓植物多样性[J]. 生物多样性, 2024, 32(9): 24096-. |
| [9] | 李东红, 郝媛媛, 甘辉林, 张航, 刘耀猛, 他富源, 胡桂馨. 祁连山北麓中段不同类型草地蝗虫种类及分布[J]. 生物多样性, 2024, 32(9): 24119-. |
| [10] | 牛红玉, 陈璐, 赵恒月, 古丽扎尔·阿不都克力木, 张洪茂. 城市化对动物的影响: 从群落到个体[J]. 生物多样性, 2024, 32(8): 23489-. |
| [11] | 白雪, 李正飞, 刘洋, 张君倩, 张多鹏, 罗鑫, 杨佳莉, 杜丽娜, 蒋玄空, 武瑞文, 谢志才. 西江流域大型底栖无脊椎动物物种多样性及维持机制[J]. 生物多样性, 2024, 32(7): 23499-. |
| [12] | 许佳, 崔小娟, 张翼飞, 吴昌, 孙远东. 南岭地区鱼类多样性及其地理分布[J]. 生物多样性, 2024, 32(7): 23482-. |
| [13] | 邝起宇, 胡亮. 广东东海岛与硇洲岛海域底栖贝类物种多样性及其地理分布[J]. 生物多样性, 2024, 32(5): 24065-. |
| [14] | 赵勇强, 阎玺羽, 谢加琪, 侯梦婷, 陈丹梅, 臧丽鹏, 刘庆福, 隋明浈, 张广奇. 退化喀斯特森林自然恢复中不同生活史阶段木本植物物种多样性与群落构建[J]. 生物多样性, 2024, 32(5): 23462-. |
| [15] | 徐伟强, 苏强. 分形模型与一般性物种多度分布关系的检验解析:以贝类和昆虫群落为例[J]. 生物多样性, 2024, 32(4): 23410-. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
备案号:京ICP备16067583号-7
Copyright © 2022 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn