
|
|
||
|
传粉昆虫下降背景下的授粉生态弹性: 内涵、机制和展望
生物多样性
2021, 29 (7):
980-994.
DOI: 10.17520/biods.2020470
全球传粉昆虫多样性正在下降, 如何保障农林生态系统传粉功能是当前研究的热点。理论上说, 传粉功能不仅与生态系统的传粉昆虫多样性相关, 还与生态系统的调节能力有关。近年来, 学者们逐渐认识到授粉生态弹性对传粉功能的影响。本文在回顾已有研究的基础之上, 总结传粉昆虫授粉生态弹性的内涵, 厘清授粉生态弹性与工程弹性、稳定性和抗性的异同。目前, 学者对授粉生态弹性形成机制开展广泛探讨, 提出功能冗余假说、密度补偿假说、响应多样性假说、连接周转假说和跨尺度弹性假说, 但这5个假说间的关系仍不清楚, 存在一词多义、词意混淆等现象。我们依次阐述功能冗余假说、密度补偿假说、响应多样性假说、连接周转假说和跨尺度弹性假说, 介绍不同假说中授粉生态弹性形成过程、研究热点和发展动态。通过解析授粉生态弹性的形成机制可知, 5个假说在内涵上存在紧密联系, 它们从不同空间尺度和研究对象下解释传粉昆虫授粉生态弹性的形成机制。未来授粉生态弹性研究将整合传粉昆虫群落动态和传粉功能动态的量化方法, 通过实验验证5个假说的合理性, 并揭示不同假说间的联系, 由此阐明授粉生态弹性的发生条件、形成阈值和动态规律。随着研究的深入, 授粉生态弹性理论有望用于指导农林生态系统传粉功能的经营管理。 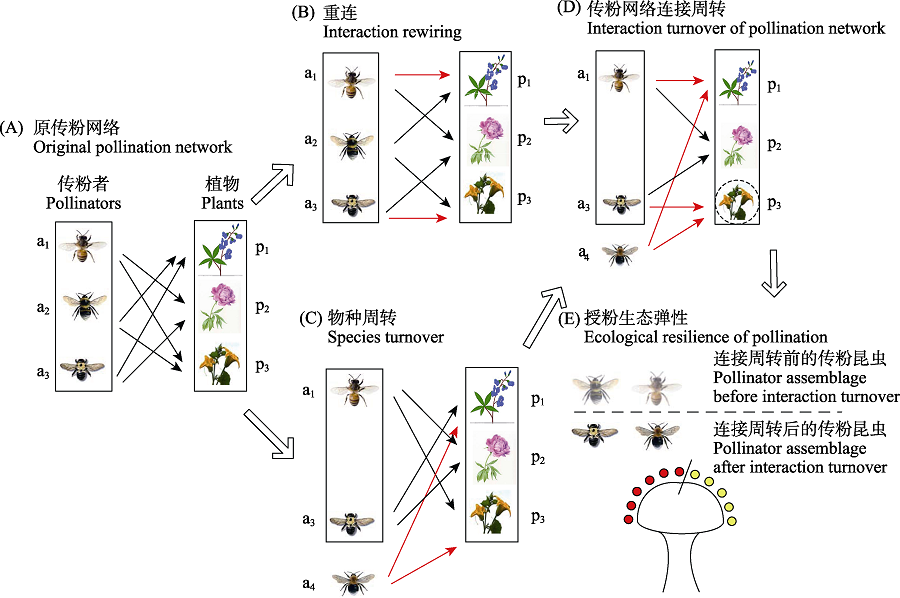 View image in article
图5
连接周转引起授粉生态弹性的机制示意图。在生态系统中, 假设原传粉网络存在3种传粉昆虫(a1, a2, a3)和3种开花植物(p1, p2, p3) (A)。干扰作用下, 传粉昆虫和开花植物间部分连接消失但新的连接形成, 网络连接发生重连(B); 或者传粉昆虫下降, 对应的连接消失, 或新的传粉昆虫(a4)进入网络, 建立新连接, 传粉昆虫发生物种周转(C)。在传粉网络重连和物种周转的共同作用下, 传粉昆虫和开花植物形成传粉网络连接周转(D)。对开花植物来说, 连接周转前后传粉昆虫群落组成存在差异, 但原传粉网络中的传粉昆虫和连接周转后的传粉昆虫传播等量花粉, 形成授粉生态弹性(E)。黑线示意周转前传粉昆虫同开花植物间的联系, 红线示意形成的新连接。
正文中引用本图/表的段落
传粉网络的连接周转(或连接替换)是指干扰作用下传粉昆虫和开花植物的部分连接消失但同时建立新连接的现象(图5) (Poisot et al, 2012; Simanonok & Burkle, 2014)。传粉昆虫和开花植物间的连接重连(interaction rewiring)和物种周转(species turnover)均可引起传粉网络的连接周转(Poisot et al, 2012; CaraDonna et al, 2017)。连接重连指群落组成不变的情况下传粉昆虫和开花植物部分连接消失但同时建立新连接(图5B) (Kaiser-Bunbury et al, 2010; Simanonok & Burkle, 2014)。物种周转指部分物种消失或新物种进入网络后传粉昆虫或开花植物因物种组成变化而形成新连接(图5C) (CaraDonna et al, 2017)。网络重连和物种更替往往同时发生, 但在不同的生态系统中二者对网络变化的相对贡献存在差异(图5D) (Simanonok & Burkle, 2014; Robroek et al, 2021)。从开花植物的角度来看, 网络变化后部分连接和传粉昆虫消失, 对应的传粉功能下降, 但新连接的传粉昆虫同样可以为开花植物授粉, 在传粉功能上表现为补偿作用(图5E)。由此可见, 网络变化后传粉昆虫群落组成和多度维持在原网络相似水平时, 传粉昆虫群落形成授粉生态弹性。
前期研究多假设传粉网络为静止状态, 主要描述传粉网络的物种组成和拓扑结构的特征(Memmott et al, 2004; Bascompte & Jordano, 2007)。近年来, 学者们普遍认为自然界中传粉网络是动态变化的, 传粉昆虫和开花植物的连接周转是常见的生态现象(Tylianakis & Morris, 2017)。在干扰作用下, 传粉昆虫与开花植物的平行下降(parallel decline)和传粉昆虫提升对开花植物的相遇概率(encounter probability)都可以引起传粉昆虫和开花植物的新连接(图5) (Biesmeijer, 2006; Tylianakis & Morris, 2017)。例如, 在气候变化和生物入侵等作用下, 生态系统传粉网络可以形成新的连接(Burkle & Alarcón, 2011; Burkle et al, 2013; Kaiser-Bunbury et al, 2017)。
普遍的观点认为连接周转可以使传粉昆虫形成授粉生态弹性(Kaiser-Bunbury et al, 2017; Fantinato et al, 2019)。从网络水平来看, 网络变化后传粉网络拓扑结构稳固性(robustness)并不下降(Kaiser-Bunbury et al, 2010; Vizentin-Bugoni et al, 2020)。从群落水平来看, 网络变化后开花植物的传粉昆虫泛化度(generality)保持不变(Fantinato et al, 2019)。在物种水平上来看, 网络变化后开花植物连接的传粉昆虫多样性(partner diversity)保持不变(Fantinato et al, 2019)。不同水平的指标表明, 连接周转后虽然传粉昆虫群落结构和组成发生变化, 但变化后的传粉昆虫同样为开花植物授粉, 传粉昆虫群落形成授粉生态弹性(图5D)。然而, 网络变化后传粉功能受昆虫的传粉效率和访花频率的影响, 而访花频率又受开花植物和传粉昆虫间物候性状的匹配性、开花植物的特化程度以及它们之间互作关系的影响(Garibaldi et al, 2015; CaraDonna et al, 2017; Kaiser-Bunbury et al, 2017)。因而, 传粉昆虫是否形成授粉生态弹性还取决于网络变化后开花植物的传粉昆虫群落结构。例如, 传粉昆虫物种或多度下降后, 传粉昆虫间的竞争作用降低, 增加了传粉昆虫访问其他开花植物的概率。传粉昆虫对开花植物的忠诚度(floral fidelity)下降, 同一开花植物间的花粉传播数量下降(Brosi & Briggs, 2013)。又如, 网络变化后春美草(Claytonia virginica)的传粉昆虫访花频率大幅下降, 传粉功能冗余度降低, 为开花植物提供的传粉功能降低(Burkle et al, 2013)。在此条件下, 网络变化后传粉昆虫未能使生态系统的传粉功能保持不变或维持在可接受的水平, 传粉昆虫群落未形成授粉生态弹性。因此, 传粉昆虫是否形成授粉生态弹性还取决于网络变化后传粉昆虫群落结构。
弹性可简单分为生态弹性(ecological resilience)和工程弹性(engineering resilience) ( Density compensation can cause no effect of biodiversity on ecosystem functioning 1 2007 ... 密度补偿作用的研究多以植物、哺乳动物和鸟类为对象, 对传粉昆虫的研究相对较少( Ecosystem restoration strengthens pollination network resilience and function 3 2017 ... 前期研究多假设传粉网络为静止状态, 主要描述传粉网络的物种组成和拓扑结构的特征(
弹性可简单分为生态弹性(ecological resilience)和工程弹性(engineering resilience) ( Partitioning interaction turnover among alpine pollination networks: Spatial, temporal, and environmental patterns 3 2014 ... 传粉网络的连接周转(或连接替换)是指干扰作用下传粉昆虫和开花植物的部分连接消失但同时建立新连接的现象(
本文的其它图/表
|

{kind=link}