
生物多样性 ›› 2012, Vol. 20 ›› Issue (3): 337-347. DOI: 10.3724/SP.J.1003.2012.11243 cstr: 32101.14.SP.J.1003.2012.11243
所属专题: 传粉生物学:理论探讨与初步实践; 传粉生物学
刘欣欣1,2, 吴小琴1,*( ), 张奠湘1
), 张奠湘1
收稿日期:2011-12-30
接受日期:2012-04-10
出版日期:2012-05-20
发布日期:2012-05-09
通讯作者:
吴小琴
作者简介:*E-mail: wxq@scbg.ac.cn基金资助:
Xinxin Liu1,2, Xiaoqin Wu1,*(), Dianxiang Zhang1
Received:2011-12-30
Accepted:2012-04-10
Online:2012-05-20
Published:2012-05-09
Contact:
Xiaoqin Wu
摘要:
异型花柱是一种受遗传因素控制的花型多态性现象, 包括二型花柱和三型花柱两种类型。本文以茜草科艳丽耳草(Hedyotis pulcherrima)为实验材料, 通过对其野外居群的花型、花部形态及花粉特征等观察, 发现艳丽耳草野外居群同时存在长花柱型花和短花柱型花, 长/短花柱型花的数量比例为1:1。两型花具有精确的交互式雌雄异位特征, 并且该特征与花冠长度相关性显著。长/短花柱型花的柱头裂片长度、花粉大小及淀粉含量等具有二型性。花粉体外培养时花粉萌发率及花粉管生长速率无显著性差异。人工授粉后, 艳丽耳草长/短花柱型花型间异交花粉管生长形态正常, 授粉24 h后花粉管均已进入子房。而长/短花柱型花在自交及型内异交下均表现为不亲和, 花粉管生长停止于柱头, 花粉管顶端累积胼胝质并膨大。艳丽耳草没有无融合生殖现象, 型间人工辅助异交授粉结实率为100%, 显著高于自然结实率。本研究结果表明, 艳丽耳草是典型的二型花柱植物, 并具有异型自交不亲和 系统。
刘欣欣, 吴小琴, 张奠湘 (2012) 艳丽耳草的二型花柱及异型自交不亲和系统. 生物多样性, 20, 337-347. DOI: 10.3724/SP.J.1003.2012.11243.
Xinxin Liu, Xiaoqin Wu, Dianxiang Zhang (2012) Distyly and heteromorphic self-incompatibility of Hedyotis pulcherrima (Rubiaceae). Biodiversity Science, 20, 337-347. DOI: 10.3724/SP.J.1003.2012.11243.
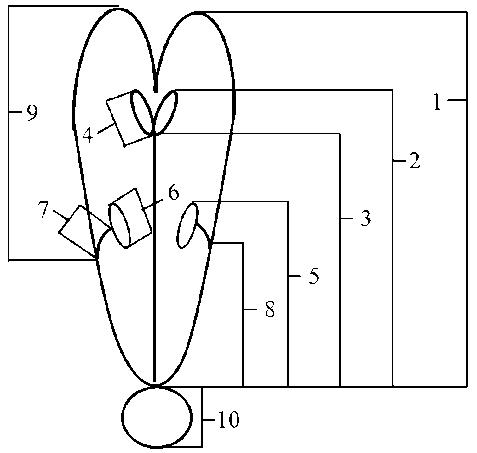
图1 艳丽耳草花部特征参数测量。(1)花冠长度; (2)柱头高度; (3)花柱长度; (4)柱头裂片长度; (5)花药高度; (6)花药长度; (7)花丝长度; (8)花丝基部至花冠基部距离; (9)花丝基部至花冠顶端距离; (10)子房高度。
Fig.1 Floral measurements of Hedyotis pulcherrima. (1) Corolla length; (2) Stigma height; (3) Style height; (4) Stigma lobe length; (5) Anther height; (6) Anther length; (7) Filament length; (8) From filament attachment to the base of corolla; (9) From filament attachment to the apex of corolla; (10) Ovary height.

图2 艳丽耳草的二型花。(A)长花柱型; (B)短花柱型; (C)长/短花柱花交互式雌雄异位, 左边是长花柱型花, 右边是短花柱型花。St: 柱头; An: 花药; 标尺 = 3 mm。
Fig. 2 Distylous flower of Hedyotis pulcherrima. (A) Long-styled flower; (B) Short-styled flower; (C) Reciprocal herkogamy, the left is long-styled flower, and the right is short-styled flower. St, Stigma; An, Anther. Bar = 3 mm.
| 花部和花粉特征 Floral and pollen attributes | 长花柱型 Long-styled morph (n = 30) | 短花柱型 Short-styled morph (n = 30) | t-检验 t-test |
|---|---|---|---|
| 花冠长度 Corolla length (mm) | 18.34±1.21 | 18.55±1.65 | 0.56 |
| 柱头高度 Stigma height (mm) | 15.13±0.94 | 11.18±1.03 | 15.47** |
| 花柱长度 Style height (mm) | 13.61±0.89 | 8.60±0.82 | 61.63** |
| 柱头裂片长度 Stigma lobe length (mm) | 1.63±0.25 | 2.83±0.46 | 12.48** |
| 花药高度 Anther height (mm) | 11.12±0.76 | 14.62±1.27 | 12.94** |
| 花药长度 Anther length (mm) | 2.11±0.18 | 2.38±0.16 | 6.10** |
| 花丝长度 Filament length (mm) | 0.42±0.11 | 1.22±0.14 | 24.93** |
| 花丝基部至花冠基部距离 From filament attachment to the base of corolla (mm) | 9.84±0.70 | 12.42±1.28 | 9.69** |
| 花丝基部至花冠顶端距离 From filament attachment to the apex of corolla (mm) | 8.31±0.65 | 6.25±0.66 | 12.15** |
| 子房高度 Ovary height (mm) | 2.11±0.19 | 2.22±0.26 | 1.83* |
| 花粉数 No. of pollen grains per flower | 8,980±144.05 | 7,988±181.85 | 9.56** |
| 胚珠数 No. of ovules per flower | 79.2±6.75 | 83.6±5.78 | 2.09* |
| 花粉/胚珠比 P/O ratio | 113.51±6.88 | 95.59±1.43 | 10.75** |
| 花粉极轴长度 Polar axis of pollen (P) (μm) | 24.89±1.42 | 27.18±2.16 | 4.85** |
| 花粉赤道长度 Equatorial axis of pollen (E) (μm) | 23.38±1.65 | 25.02±1.35 | 4.2** |
| 花粉极轴长/赤道轴长 (P/E) | 1.05±0.06 | 1.03±0.07 | 1.19 |
| 网孔直径 Diameter of lumina (μm) | 0.56±0.11 | 0.51±0.08 | 0.39 |
表1 艳丽耳草长/短花柱型花部及花粉特征参数
Table 1 Floral and pollen attributes of long-styled and short-styled flowers of Hedyotis pulcherrima
| 花部和花粉特征 Floral and pollen attributes | 长花柱型 Long-styled morph (n = 30) | 短花柱型 Short-styled morph (n = 30) | t-检验 t-test |
|---|---|---|---|
| 花冠长度 Corolla length (mm) | 18.34±1.21 | 18.55±1.65 | 0.56 |
| 柱头高度 Stigma height (mm) | 15.13±0.94 | 11.18±1.03 | 15.47** |
| 花柱长度 Style height (mm) | 13.61±0.89 | 8.60±0.82 | 61.63** |
| 柱头裂片长度 Stigma lobe length (mm) | 1.63±0.25 | 2.83±0.46 | 12.48** |
| 花药高度 Anther height (mm) | 11.12±0.76 | 14.62±1.27 | 12.94** |
| 花药长度 Anther length (mm) | 2.11±0.18 | 2.38±0.16 | 6.10** |
| 花丝长度 Filament length (mm) | 0.42±0.11 | 1.22±0.14 | 24.93** |
| 花丝基部至花冠基部距离 From filament attachment to the base of corolla (mm) | 9.84±0.70 | 12.42±1.28 | 9.69** |
| 花丝基部至花冠顶端距离 From filament attachment to the apex of corolla (mm) | 8.31±0.65 | 6.25±0.66 | 12.15** |
| 子房高度 Ovary height (mm) | 2.11±0.19 | 2.22±0.26 | 1.83* |
| 花粉数 No. of pollen grains per flower | 8,980±144.05 | 7,988±181.85 | 9.56** |
| 胚珠数 No. of ovules per flower | 79.2±6.75 | 83.6±5.78 | 2.09* |
| 花粉/胚珠比 P/O ratio | 113.51±6.88 | 95.59±1.43 | 10.75** |
| 花粉极轴长度 Polar axis of pollen (P) (μm) | 24.89±1.42 | 27.18±2.16 | 4.85** |
| 花粉赤道长度 Equatorial axis of pollen (E) (μm) | 23.38±1.65 | 25.02±1.35 | 4.2** |
| 花粉极轴长/赤道轴长 (P/E) | 1.05±0.06 | 1.03±0.07 | 1.19 |
| 网孔直径 Diameter of lumina (μm) | 0.56±0.11 | 0.51±0.08 | 0.39 |
| 花冠长度 Corolla length | 柱头高度 Stigma height | 花柱长度Style height | 柱头裂 片长度Stigma lobe length | 花药高度 Anther height | 花药长度 Anther length | 花丝长度Filament length | 花丝基部至花冠基部距离 From filament attachment to the base of corolla | 花丝基部至花冠顶端距离From filament attachment to the apex of corolla | |
|---|---|---|---|---|---|---|---|---|---|
| 长花柱型 Long-styled morph | |||||||||
| 柱头高度 Stigma height | 0.566** | ||||||||
| 花柱长度 Style height | 0.501** | 0.935** | |||||||
| 柱头裂片长度 Stigma lobe length | 0.373* | 0.540** | 0.206 | ||||||
| 花药高度 Anther height | 0.796** | 0.452* | 0.417* | 0.257 | |||||
| 花药长度 Anther length | 0.278 | 0.344 | 0.196 | 0.483** | 0.248 | ||||
| 花丝长度 Filament length | 0.475** | 0.261 | 0.286 | 0.040 | 0.368* | 0.184 | |||
| 花丝基部至花冠基部距离 From filament attachment to the base of corolla | 0.836** | 0.391* | 0.327 | 0.301 | 0.934** | 0.231 | 0.323 | ||
| 花丝基部至花冠顶端距离 From filament attachment to the apex of corolla | 0.723** | 0.442* | 0.429* | 0.202 | 0.377* | 0.319 | 0.325 | 0.411* | |
| 子房高度 Ovary height | 0.510** | 0.533** | 0.498** | 0.290 | 0.263 | 0.203 | 0.492** | 0.199 | 0.495** |
| 短花柱型 Short-styled morph | |||||||||
| 柱头高度 Stigma height | 0.723** | ||||||||
| 花柱长度 Style height | 0.606** | 0.873** | |||||||
| 柱头裂片长度 Stigma lobe length | 0.490** | 0.623** | 0.162 | ||||||
| 花药高度 Anther height | 0.825** | 0.620** | 0.583** | 0.319 | |||||
| 花药长度 Anther length | -0.206 | 0.056 | 0.058 | 0.020 | -0.109 | ||||
| 花丝长度 Filament length | 0.135 | 0.080 | 0.085 | 0.025 | 0.059 | -0.379* | |||
| 花丝基部至花冠基部距离 From filament attachment to the base of corolla | 0.902** | 0.732** | 0.620** | 0.486** | 0.875** | -0.238 | 0.154 | ||
| 花丝基部至花冠顶端距离 From filament attachment to the apex of corolla | 0.748** | 0.492** | 0.391* | 0.369* | 0.426* | -0.276 | 0.297 | 0.493** | |
| 子房高度 Ovary height | 0.336 | 0.307 | 0.250 | 0.221 | 0.046 | 0.091 | 0.125 | 0.097 | 0.501** |
表2 艳丽耳草长/短花柱型花的花部特征间的Pearson相关性分析
Table 2 Pearson correlation coefficients for traits of long-styled and short-styled flowers in Hedyotis pulcherrima
| 花冠长度 Corolla length | 柱头高度 Stigma height | 花柱长度Style height | 柱头裂 片长度Stigma lobe length | 花药高度 Anther height | 花药长度 Anther length | 花丝长度Filament length | 花丝基部至花冠基部距离 From filament attachment to the base of corolla | 花丝基部至花冠顶端距离From filament attachment to the apex of corolla | |
|---|---|---|---|---|---|---|---|---|---|
| 长花柱型 Long-styled morph | |||||||||
| 柱头高度 Stigma height | 0.566** | ||||||||
| 花柱长度 Style height | 0.501** | 0.935** | |||||||
| 柱头裂片长度 Stigma lobe length | 0.373* | 0.540** | 0.206 | ||||||
| 花药高度 Anther height | 0.796** | 0.452* | 0.417* | 0.257 | |||||
| 花药长度 Anther length | 0.278 | 0.344 | 0.196 | 0.483** | 0.248 | ||||
| 花丝长度 Filament length | 0.475** | 0.261 | 0.286 | 0.040 | 0.368* | 0.184 | |||
| 花丝基部至花冠基部距离 From filament attachment to the base of corolla | 0.836** | 0.391* | 0.327 | 0.301 | 0.934** | 0.231 | 0.323 | ||
| 花丝基部至花冠顶端距离 From filament attachment to the apex of corolla | 0.723** | 0.442* | 0.429* | 0.202 | 0.377* | 0.319 | 0.325 | 0.411* | |
| 子房高度 Ovary height | 0.510** | 0.533** | 0.498** | 0.290 | 0.263 | 0.203 | 0.492** | 0.199 | 0.495** |
| 短花柱型 Short-styled morph | |||||||||
| 柱头高度 Stigma height | 0.723** | ||||||||
| 花柱长度 Style height | 0.606** | 0.873** | |||||||
| 柱头裂片长度 Stigma lobe length | 0.490** | 0.623** | 0.162 | ||||||
| 花药高度 Anther height | 0.825** | 0.620** | 0.583** | 0.319 | |||||
| 花药长度 Anther length | -0.206 | 0.056 | 0.058 | 0.020 | -0.109 | ||||
| 花丝长度 Filament length | 0.135 | 0.080 | 0.085 | 0.025 | 0.059 | -0.379* | |||
| 花丝基部至花冠基部距离 From filament attachment to the base of corolla | 0.902** | 0.732** | 0.620** | 0.486** | 0.875** | -0.238 | 0.154 | ||
| 花丝基部至花冠顶端距离 From filament attachment to the apex of corolla | 0.748** | 0.492** | 0.391* | 0.369* | 0.426* | -0.276 | 0.297 | 0.493** | |
| 子房高度 Ovary height | 0.336 | 0.307 | 0.250 | 0.221 | 0.046 | 0.091 | 0.125 | 0.097 | 0.501** |
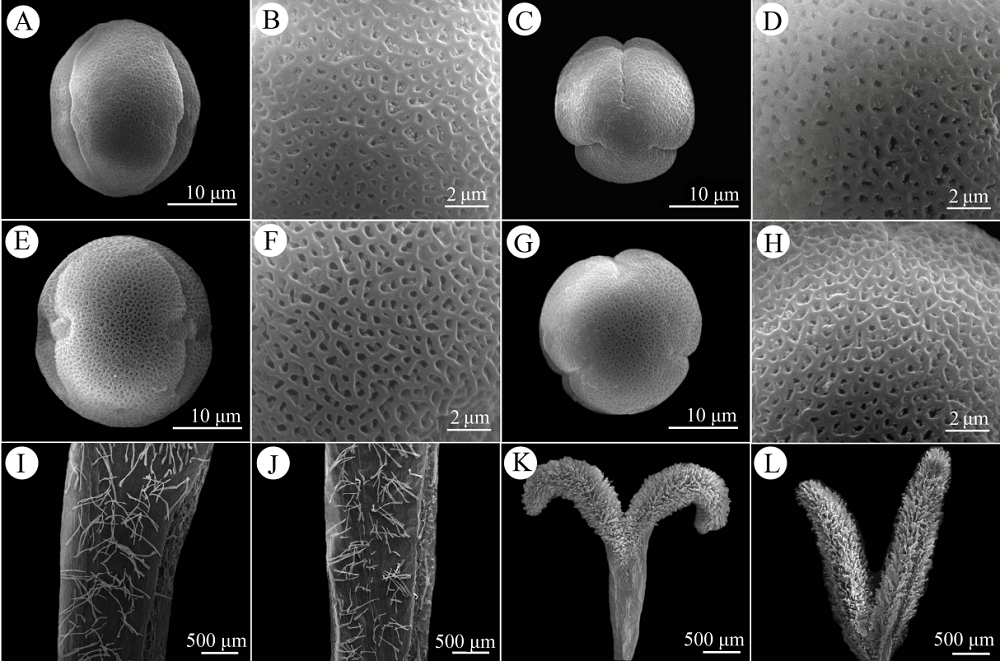
图3 艳丽耳草长/短花柱型花粉、花冠及柱头扫描电镜图。(A, B)长花柱型花粉赤道面; (C, D)长花柱型花粉极面; (E, F)短花柱型花粉赤道面; (G, H)短花柱型花粉极面; (I)长花柱型花冠喉部的毛; (J)短花柱型花冠喉部的毛; (K)长花柱型湿性柱头, 2裂; (L)短花柱型湿性柱头, 2裂。
Fig. 3 Morphology of pollen grains, corolla tubes and stigmas of Hedyotis pulcherrima under scanning electron microscopy. Equatorial (A, B) and polar view (C, D) of pollen grains from long-styled flowers; equatorial (E, F) and polar (G, H) view of pollen grains from short-styled flowers; pubescence at the throat of the corolla tube of a long-styled (I) and a short-styled (J) flower; bilobed stigma of the wet papillate type in a long-styled (K) and short-styled (L) flower.
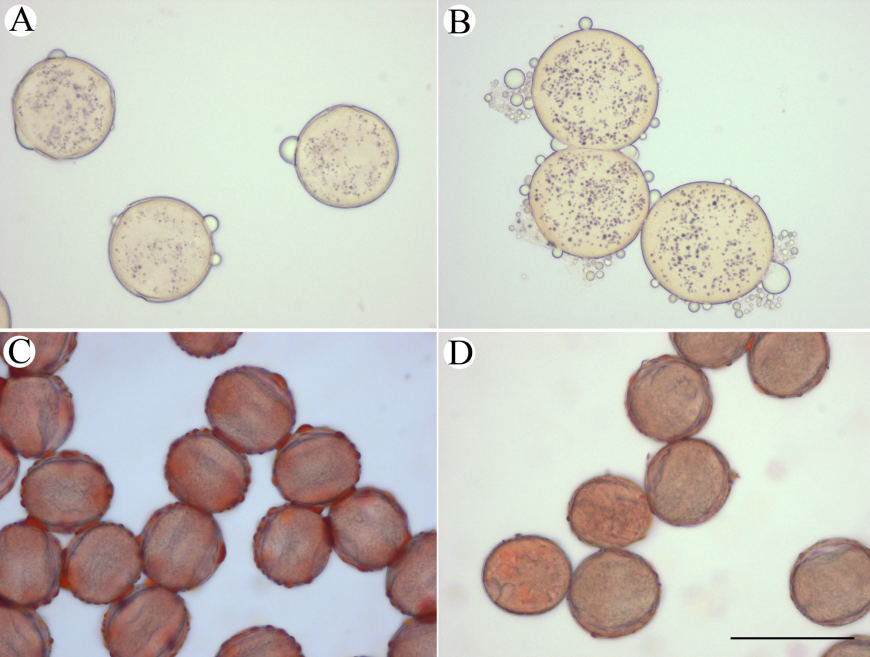
图4 艳丽耳草长/短花柱型花粉组织化学染色。(A)长花柱型花粉KI-I2染色; (B)短花柱型花粉KI-I2染色; (C)长花柱型花粉苏丹IV染色; (D)短花柱型花粉苏丹IV染色。标尺= 50 μm。
Fig. 4 Histochemical staining of Hedyotis pulcherrima pollen grains. (A) KI-I2 staining of long-styled pollen grains; (B) KI-I2 staining of short- styled pollen grains; (C) Sudan IV staining of long-styled pollen grains; (D) Sudan IV staining of short-styled pollens. Bar =50 μm.
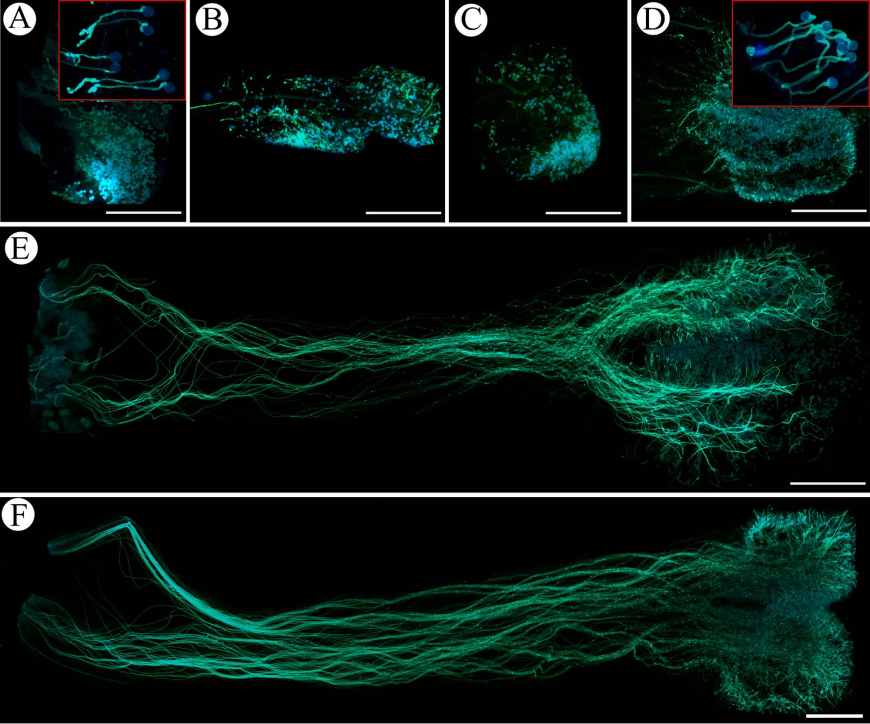
图5 艳丽耳草人工授粉处理后花粉管体内生长图。(A)长花柱型花自交处理24 h后, 红框内示花粉管顶端膨大; (B)短花柱型花自交处理24 h后; (C)长花柱型花型内异交处理24 h后; (D)短花柱型花型内异交处理24 h后, 红框内示花粉管顶端膨大; (E)短花柱型花型间异交处理24 h后; (F)长花柱型花型间异交处理24 h后; 标尺=1 mm。
Fig. 5 Pollen tube growth in vivo of Hedyotis pulcherrima following self-, intramorph and inter- morph pollination. (A) Long-styled morph stigma lobes and style 24 h after self-pollination; red ellipse showing accumulation of callose in the swollen tube; (B) Short-styled morph stigma lobes and style 24 h after self-pollination; (C) Long-styled morph stigma lobes and style 24 h after intramorph pollination; (D) Short-styled morph stigma lobes and style 24 h after intramorph pollination; red ellipse showing accumulation of callose in the swollen tube; (E) Short-styled morph stigma lobes and style 24 h after intermorph pollination; (F) Long-styled morph stigma lobes and style 24 h after intermorph pollination. Bar = 1 mm.
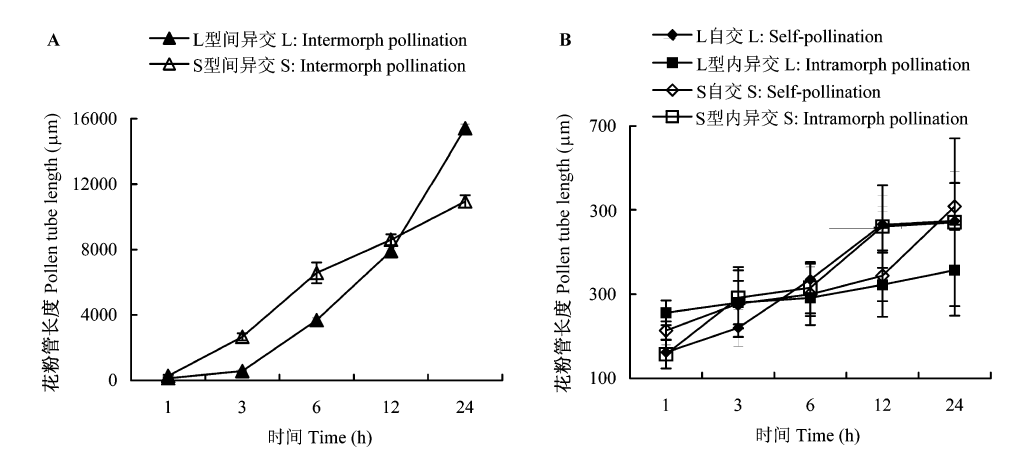
图6 艳丽耳草花粉管体内生长速度。(A)型间异交下花粉管生长速度; (B)自交及型内异交下花粉管生长速度。L: 长花柱型; S: 短花柱型。
Fig. 6 Pollen tube growth rate of Hedyotis pulcherrima in vivo. (A) In intermorph pollination; (B) In self- and intramorph pollination. L, Long-styled morph; S, Short-styled morph.
| 处理方式 Treatment | 结实率 Fruit sets (%) | t-检验 t-test | |
|---|---|---|---|
| 长花柱型 Long-styled morph | 短花柱型 Short-styled morph | ||
| 自交 Self-pollination | 0 (n = 35) | 0 (n = 52) | – |
| 型内异交 Intramorph pollination | 0 (n = 37) | 0 (n = 30) | – |
| 型间异交 Intermorph pollination | 100 (n = 31) | 100 (n = 29) | – |
| 去雄套袋 Netting with emasculation | 0 (n = 83) | 0 (n = 63) | – |
| 自然套袋 Netting without emasculation | 0 (n = 53) | 0 (n = 54) | – |
| 自然 Open pollination | 70.2±16.2 (n = 58) | 68.2±21.3 (n = 93) | 0.08 |
表3 艳丽耳草人工辅助授粉结实率
Table 3 Fruit sets of Hedyotis pulcherrima under artificial pollination
| 处理方式 Treatment | 结实率 Fruit sets (%) | t-检验 t-test | |
|---|---|---|---|
| 长花柱型 Long-styled morph | 短花柱型 Short-styled morph | ||
| 自交 Self-pollination | 0 (n = 35) | 0 (n = 52) | – |
| 型内异交 Intramorph pollination | 0 (n = 37) | 0 (n = 30) | – |
| 型间异交 Intermorph pollination | 100 (n = 31) | 100 (n = 29) | – |
| 去雄套袋 Netting with emasculation | 0 (n = 83) | 0 (n = 63) | – |
| 自然套袋 Netting without emasculation | 0 (n = 53) | 0 (n = 54) | – |
| 自然 Open pollination | 70.2±16.2 (n = 58) | 68.2±21.3 (n = 93) | 0.08 |
| [1] | Bahadur B (1968) Heterostyly in Rubiaceae: a review. Journal of Osmania University (Science). Golden Jubilee Special Volume, 207-238. |
| [2] | Barrett SCH, Jesson LK, Baker AM (2000) The evolution and function of stylar polymorphisms in flowering plants. Annals of Botany, 85, 253-256. |
| [3] | Barrett SCH, Shore JS (2008) New insights on heterostyly: comparative biology, ecology and genetics. In: Self-Incompatibility in Flowering Plants: Evolution, Diversity, and Mechanisms (ed. Franklin-Tong VE), pp. 3-32. Springer-Verlag, Berlin. |
| [4] | Bawa KS, Beach JH (1983) Self-incompatibility systems in the Rubiaceae of a tropical lowland wet forest. American Journal of Botany, 70, 1281-1288. |
| [5] | Bremer B, Manen JF (2000) Phylogeny and classification of the subfamily Rubioideae (Rubiaceae). Plant Systematics and Evolution, 225, 43-72. |
| [6] |
Castro CC, Oliveira PEAM, Alves MC (2004) Breeding system and floral morphometry of distylous Psychotria L. species in the Atlantic rain forest, SE Brazil. Plant Biology, 6, 755-760.
DOI URL PMID |
| [7] | Charlesworth D, Charlesworth B (1979) A model for the evolution of distyly. The American Naturalist, 114, 467-498. |
| [8] | Consolaro H, Silva SCS, Oliveira PE (2011) Breakdown of distyly and pin-monomorphism in Psychotria carthagenensis Jacq. (Rubiaceae). Plant Species Biology, 26, 24-32. |
| [9] |
Cruden RW, Lyon DL (1985) Patterns of biomass allocation to male and female functions in plants with different mating systems. Oecologia, 66, 299-306.
DOI URL PMID |
| [10] | Dafni A (1992) Pollination Ecology: A Practical Approach. Oxford University Press, New York. |
| [11] | Darwin C (1877) The Different Forms of Flowers on Plants of the Same Species. John Murray, London. |
| [12] | Dulberger R (1992) Floral polymorphisms and their functional significance in the heterostylous syndrome. In: Evolution and Function of Heterostyly (ed. Barrett SCH), pp. 41-84. Springer-Verlag, New York. |
| [13] |
Faivre AE (2002) Variation in pollen tube inhibition sites within and among three heterostylous species of Rubiaceae. International Journal of Plant Sciences, 163, 783-794.
DOI URL |
| [14] | Ganders FR (1979) The biology of heterostyly. New Zealand Journal of Botany, 17, 607-635. |
| [15] |
Geitmann A, Franklin-Tong VE, Emons AC (2004) The self-incompatibility response in Papaver rhoeas pollen causes early and striking alterations to organelles. Cell Death and Differentiation, 11, 812-822.
URL PMID |
| [16] | Guo X (郭兴) (2011) Phylogenetic Study on the Genus Hedyotis L. (Rubiaceae) (耳草属植物系统发育关系的研究). Master thesis, South China Botanical Garden, Chinese Academy of Sciences, Guangzhou. (in Chinese with English abstract) |
| [17] |
Hernández A, Ornelas JF (2007) Development of distylous flowers and investment of biomass in male and female function in Palicourea padifolia (Rubiaceae). Plant Biology, 9, 694-704.
DOI URL PMID |
| [18] |
Kårehed J, Groeninckx I, Dessein S, Motley TJ, Bremer B (2008) The phylogenetic utility of chloroplast and nuclear DNA markersand the phylogeny of the Rubiaceae tribe Spermacoceae. Molecular Phylogenetics and Evolution, 49, 843-866.
DOI URL PMID |
| [19] |
Klein DE, Freitas L, Da Cunha M (2009) Self-incompatibility in a distylous species of Rubiaceae: Is there a single incompatibility response of the morphs? Sexual Plant Reproduction, 22, 121-131.
URL PMID |
| [20] | Ko WC (高蕴璋) (1999) Hedyotis Linn. In: Flora Reipublicae Popularis Sinicae (ed. Lo HS (罗献瑞)) (中国植物志), Tomus 71, pp. 26-77. Science Press, Beijing. (in Chinese) |
| [21] | Li YQ (李永泉), Luo ZL (罗中莱), Zhang DX (张奠湘) (2007) Relationships between pollen histochemistry, pollen grain number, ovule number and pollinators in Phyllanthaceae. Biodiversity Science (生物多样性), 15, 645-651. (in Chinese with English abstract) |
| [22] | Lloyd DG (1979) Evolution towards dioecy in heterostylous populations. Plant Systematics and Evolution, 131, 71-80. |
| [23] | Lloyd DG, Webb CJ (1992) The evolution of heterostyly. In: Evolution and Function of Heterostyly (ed. Barrett SCH), pp. 151-178. Springer-Verlag, Berlin. |
| [24] | McCubbin A(2008) Heteromorphic self-incompatibility in Primula: twenty-first century tools promise to unravel a classic nineteenth century model system. In: Self-Incom- patibility in Flowering Plants: Evolution, Diversity and Mechanisms (ed. Franklin-Tong VE), pp. 289-308. Springer-Verlag, New York. |
| [25] |
Naiki A, Nagamasu H (2003) Distyly and pollen dimorphism in Damnacanthus (Rubiaceae). Journal of Plant Research, 116, 105-113.
DOI URL PMID |
| [26] | Ornduff R (1980) Heterostyly, population composition, and pollen flow in Hedyotis caerulea. American Journal of Botany, 67, 95-103. |
| [27] |
Quinet M, Cawoy V, Lefèvre I, van Miegroet F, Jacquemart AL, Kinet JM (2004) Inflorescence structure and control of flowering time and duration by light in buckwheat (Fagopyrum esculentum Moench). Journal of Experimental Botany, 55, 1509-1517.
DOI URL PMID |
| [28] | Richards AJ (1997) Plant Breeding Systems. Chapman & Hall, Cambridge. |
| [29] | Richards JH, Koptur S (1993) Floral variation and distyly in Guettarda scabra (Rubiaceae). American Journal of Botany, 80, 31-40. |
| [30] | Robbrecht E (1988) Tropical woody Rubiaceae. In: Opera Botanica Belgica, Vol 1, pp. 1-271. Nationale Plantentuin van België, Meise. |
| [31] | Robbrecht E, Manen JF (2006) The major evolutionary lineages of the coffee family (Rubiaceae, Angiosperms): combined analysis (nDNA and cpDNA) to infer the position of Coptosapelta and Luculia, and supertree construction based on rbcL, rps16, trnL-F and atpB-rbcL data. Systematics and Geography of Plants, 76, 85-146. |
| [32] |
Rounds CM, Winship LJ, Hepler PK (2011) Pollen tube energetics: respiration, fermentation and the race to the ovule. AoB Plants, plr019. doi: 10.1093/aobpla/plr019.
DOI URL PMID |
| [33] |
Sanchez JM, Ferrero V, Navarro L (2008) A new approach to the quantification of degree of reciprocity in distylous (sensu lato) plant populations. Annals of Botany, 102, 463-472.
URL PMID |
| [34] |
Valois-Cuesta H, Soriano PJ, Ornelas JF (2011) Dimorphisms and self-incompatibility in the distylous species Palicourea demissa (Rubiaceae): possible implications for its reproductive output. Journal of Plant Research, 124, 137-146.
URL PMID |
| [35] | Wagner WL, Lorence DH (1998) A new, dioecious species of Hedyotis (Rubiaceae) from Kaua’i, Hawaiian Islands, and the taxonomy of Kaua’i Hedyotis schlechtendahliana resolved. Novon, 8, 311-317. |
| [36] |
Wu XQ, Li AM, Zhang DX (2010) Cryptic self-incompatibility and distyly in Hedyotis acutangula Champ. (Rubiaceae). Plant Biology, 12, 484-494.
URL PMID |
| [37] | Zeng XF (曾宪锋), Zhuang XY (庄雪影), Tang GD (唐光大), Li QH (李清湖) (2009) A floristic study on the seed plants of Tongguzhang Mountains in east Guangdong. Journal of Wuhan Botanical Research (武汉植物学研究), 27, 381-386. (in Chinese with English abstract) |
| [38] | Zhong XS (钟小山), Xu RJ (徐瑞晶), Huang CT (黄川腾), Mo HZ (莫惠芝), Zeng XF (曾宪锋), Zhuang XY (庄雪影) (2011) Floristic diversity and dominant species of secondary forest on Fenghuangshan, Chaozhou, Guangdong. Guangdong Forestry Science and Technology (广东林业科技), 27, 19-26. (in Chinese with English abstract) |
| [1] | 丁翔, 余元钧, 宋希强, 罗毅波. 具有泛化访花者的海芋特化传粉系统[J]. 生物多样性, 2024, 32(6): 24069-. |
| [2] | 孙士国, 卢斌, 卢新民, 黄双全. 入侵植物的繁殖策略以及对本土植物繁殖的影响[J]. 生物多样性, 2018, 26(5): 457-467. |
| [3] | 吴凌云, 黄双全. 虫媒传粉植物荞麦的生物学特性与研究进展[J]. 生物多样性, 2018, 26(4): 396-405. |
| [4] | 黄至欢, 陆奇丰, 陈颖卓. 地锦苗在石灰岩土壤和红壤生境中的繁殖成功的比较[J]. 生物多样性, 2017, 25(9): 972-980. |
| [5] | 钱贞娜, 任明迅. “金虎尾路线”植物的花进化与传粉转变[J]. 生物多样性, 2016, 24(1): 95-101. |
| [6] | 钟云芳, 张哲, 宋希强, 周兆德. 海南凤仙花不同海拔种群的传粉生物学[J]. 生物多样性, 2014, 22(4): 467-475. |
| [7] | 路国辉, 武文华, 王瑞珍, 李新亮, 王英强. 野牡丹异型雄蕊的功能分化[J]. 生物多样性, 2009, 17(2): 174-181. |
| [8] | 张敬丽, 张长芹, 吴之坤, 乔琴. 探讨种间传粉在杜鹃花属自然杂交物种形成中的作用[J]. 生物多样性, 2007, 15(6): 658-665. |
| [9] | 贾效成, 李新亮, 丹阳, 路国辉, 王英强. 广东地区外来种五爪金龙的传粉生物学研究[J]. 生物多样性, 2007, 15(6): 592-598. |
| [10] | 王仲礼, 刘林德, 田国伟, 申家恒. 短柄五加开花及传粉生物学研究*[J]. 生物多样性, 1997, 05(4): 251-256. |
| [11] | 王洪新, 胡志昂. 植物的繁育系统、遗传结构和遗传多样性保护[J]. 生物多样性, 1996, 04(2): 92-96. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
备案号:京ICP备16067583号-7
Copyright © 2022 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn
![]()