
生物多样性 ›› 2021, Vol. 29 ›› Issue (3): 409-418. DOI: 10.17520/biods.2020276 cstr: 32101.14.biods.2020276
所属专题: 物种形成与系统进化
• 论坛 • 上一篇
李敏岚1,2, 王超1,*( ), 王瑞武1,*()
), 王瑞武1,*()
收稿日期:2020-07-11
接受日期:2021-01-04
出版日期:2021-03-20
发布日期:2021-01-13
通讯作者:
王超,王瑞武
作者简介:wangrw@nwpu.edu.cn#共同第一作者
基金资助:
Minlan Li1,2, Chao Wang1,*(), Ruiwu Wang1,*()
Received:2020-07-11
Accepted:2021-01-04
Online:2021-03-20
Published:2021-01-13
Contact:
Chao Wang,Ruiwu Wang
About author:First author contact:#Co-first authors
摘要:
生物科学几乎所有研究都需要物种概念作为基础, 生物多样性研究亦需要可操作的物种概念, 但现有物种概念存在不同程度的人为因素或难操作性, 对物种划分造成不利影响。本文引入“进化路径”这一概念, 说明适合度景观时刻变化着, 物种在每个进化时间点上依据瞬时适合度选择下一时刻的进化状态, 且总是沿着动态适合度景观中适合度增加的方向进化。基于演化博弈的方法, 以随机过程为例模拟物种的进化过程。进而提出路径依赖下的物种形成机制, 并在此基础上给出可操作的物种定义, 即: 针对基因、性状、生态过程等任一状态下两个群体内个体的多个变量做统计分析, 若群体之间同时在两个或多个维度状态下呈现出的不连续性d大于群体内变量呈现出的差异性σk, 则拥有相应变量的个体属于不同物种。
李敏岚, 王超, 王瑞武 (2021) 路径依赖下的物种形成机制. 生物多样性, 29, 409-418. DOI: 10.17520/biods.2020276.
Minlan Li, Chao Wang, Ruiwu Wang (2021) Path-dependent speciation in the process of evolution. Biodiversity Science, 29, 409-418. DOI: 10.17520/biods.2020276.
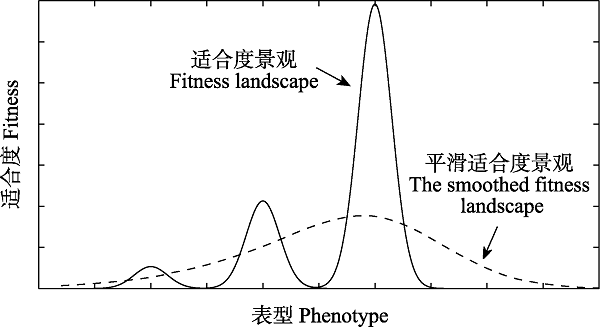
图1 表型的适合度景观图。实线表示真实的适合度景观, 当生物体在这样有高峰有山谷的适合度景观上进化时, 很容易被困在某一更低的局部峰值上。虚线是在Frank提出的反应规范作用下的表型适合度景观, 即由某基因控制的表型的适合度景观, 该景观只有一个单峰, 使得表型更容易进化到适合度更高的表型。
Fig. 1 Fitness landscape of phenotypes. The solid line shows the fitness landscape observed. When an organism evolves on such a multipeak fitness landscape, it’s easy to be trapped at a lower local peak. The dashed line displays the phenotypic fitness landscape smoothed by reaction norm, i.e., the expect fitness for a genotype with a certain average phenotype. This fitness landscape just has one single peak, which makes it easier for any phenotype to evolve to another one with higher fitness.
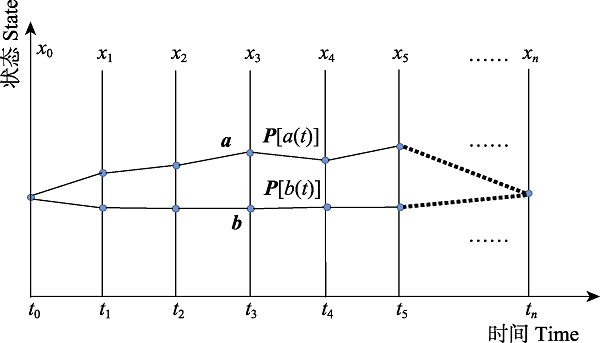
图2 群体内表型从x0进化到xn过程的示意图。个体在每一时刻都会根据瞬时适合度以一定概率去选择自己下一时刻期望进化到的表型, 因此群体内的表型从t0到tn的进化过程中可能产生多条路径, a, b即为其中的两条。假设路径a上的表型为${{x}_{0}},{{x}_{1}},{{x}_{2}},...,{{x}_{n}}$, 该路径出现的概率为$P[({{t}_{0}}),{{x}_{1}}({{t}_{1}}),{{x}_{2}}({{t}_{2}}),\ldots,{{x}_{n}}({{t}_{n}})]$。
Fig. 2 An evolutionary process for phenotype in a population from x0 to xn. An individual selects the next phenotype to evolve with a certain probability according to its instantaneous fitness at each time step, thus many possible paths of phenotype in a population occur including paths a and b. Suppose that trajectory a consists of phenotype ${{x}_{0}},{{x}_{1}},{{x}_{2}},...,{{x}_{n}}$, then the possibility is $P[{{x}_{0}}({{t}_{0}}),{{x}_{1}}({{t}_{1}}),{{x}_{2}}({{t}_{2}}),\ldots,{{x}_{n}}({{t}_{n}})]$。
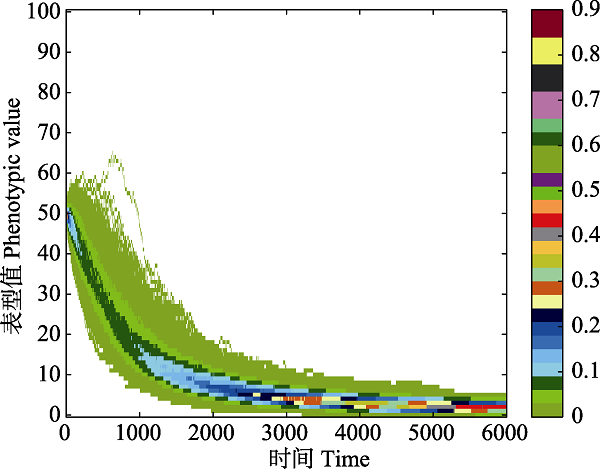
图3 进化路径的分布。不同颜色表示路径的频率。随着时间的演化, 表型从最初的状态分化出其他表型, 在此过程中, 表型只能向相邻的表型进化, 因此进化是连续的。若在t = 1,000这一时刻观察演化结果, 则可以看到明显不连续的物种分化(两个蓝色区域)。
Fig. 3 The distribution of evolutionary paths. Different colors represent different distribution probability of paths. As time goes by, one phenotype differentiates to other phenotypes with different probabilities. The evolutionary path is continuous in this process because that phenotypes can only evolve to adjacent phenotypes. If the evolutionary results are observed at the moment of t = 1,000, significant discontinuous phenotype differentiation (blue regions) can be seen.
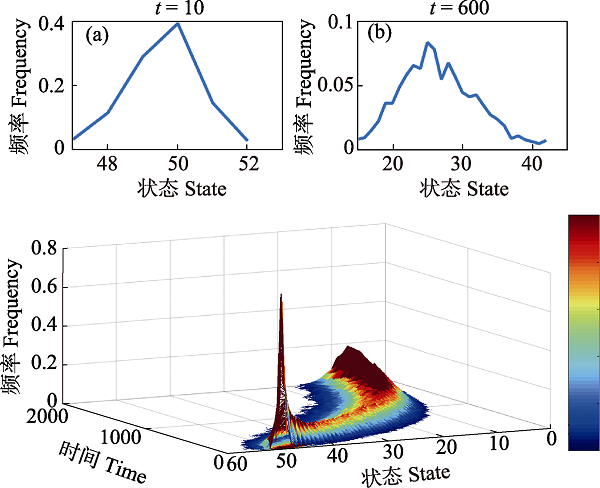
图4 路径依赖下的状态进化。(a)状态量随时间演化的频率分布。状态量可以指基因型、性状特征、生态过程。(b) t = 10时的表型频率分布。(c) t = 600时的表型频率分布。若以表型作为状态量, 分化初期显示为单峰的频率分布, 在时间t = 600处时出现明显的分化且显示为双峰的频率分布。这意味着群体内个体从初始的单一表型(50)逐渐分化成两种表型(25和28), 可以进一步判断对应的个体是否为不同物种。
Fig. 4 Path-dependent evolutionary process of states. (a) The frequency distribution curve of states over time. The states represent genotype, phenotype or ecological process. (b) The frequency distribution curve of phenotypes at t = 10, (c) The frequency distribution curve of phenotypes at t = 600. From the perspective of phenotype, the distribution curve changes from unimodal in the early phase to bimodal at t = 600, which means the phenotype differentiates into two phenotypes (25 and 28) at t = 600 from one phenotype (50) in the initial stage. Then we can judge if the species with phenotype 25 and 28 are different species.
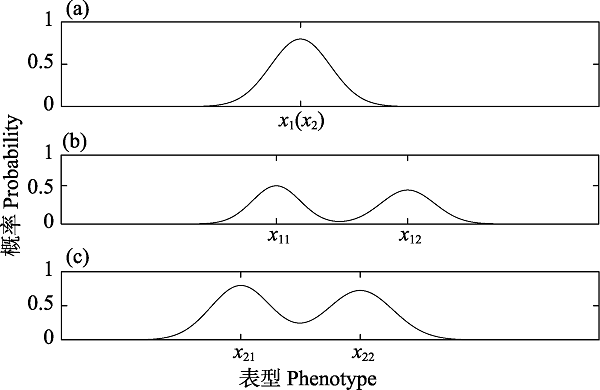
图5 不同时刻下表型和基因型的频率分布。(a)表示状态量x1(x2)未分化初期某时刻的频率分布; (b)表示群体内的表型x1在分化之后某时刻的频率分布; (c)表示群体内的基因型x2在分化之后相同时刻的频率分布。假设对于原始物种A, 其任意在两个状态x1和x2随着时间的演化受到基因突变和随机漂变的影响, 在相同或不同时刻出现不同程度的分化。当我们处于物种进化的一个“切面”去观察时, 表型x1分化成x11和x12, 并且进化依赖于特定的路径。若此时分化出的表型和基因型在数量统计上满足一定的条件, 同时在表型和基因型两个状态下同时具有特征x11, x21(x11, x22或x12, x21或x12, x22)的个体为新物种。
Fig. 5 Frequency distribution of phenotypes and genotypes at different time. (a) showing the frequency distribution curve of the state x1(x2) in the early differentiation. (b) demonstrating the frequency distribution curve of the phenotype x1 at a certain moment after differentiation. (c) indicating the frequency distribution curve of the genotype x2 at the same moment after differentiation. Suppose that any two states x1 and x2 of the original species A evolved influenced by random mutation and drift, then different degree of differentiation occurs in the same or different time. When observing in a “cutting plane” of a species evolution, we notice that x1(x2) differentiates into x11(x21) and x12(x22), and the evolution are path-dependent. Meanwhile, if the differentiated phenotypes and the genotypes in a quantity statistics satisfy certain conditions, the individuals with ${{x}_{11}},{{x}_{21}}({{x}_{11}},{{x}_{22}},or{{x}_{12}},{{x}_{21}},or{{x}_{12}},{{x}_{22}})$ are new species, respectively.
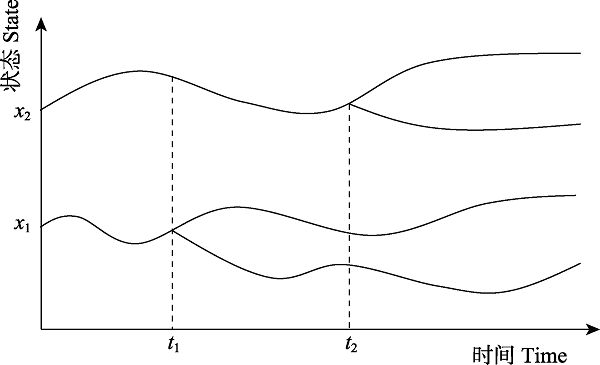
图6 随时间演替的物种界定示意图。状态量x1 (x2)在t1(t2)时刻出现分化, 在不同的时间点上根据分化情况判定物种是否属于新物种。
Fig. 6 schematic diagram of the species delimitation over time. The state x1 (x2) differentiates at t1(t2). It can be determined whether a specie at different moments is a new species.
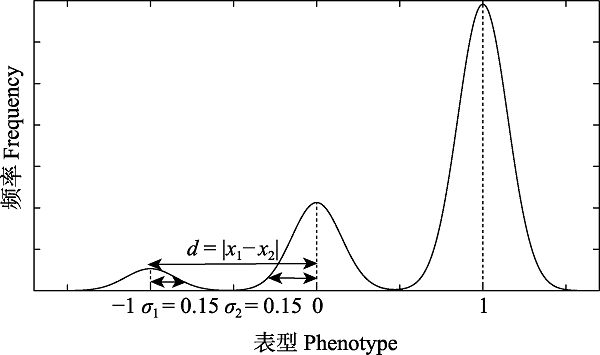
图7 一表型在某一时刻分化出三个独立表型的示意图。该图作为判定表型分化成功的一个特例, 此时种内均值为?1的表型存在连续且程度较低的差异(差异范围是± 0.15), 均值为0的亦然。而这两个种间表型的不连续性达到了1, 其不连续差异程度远高于种内连续差异程度, 因此可以认为该表型达到了形态学物种概念中对于形态的要求。同时考虑原始物种的其他状态上的分化, 若满足同样条件的差异, 即可将相应个体确定为新物种。
Fig. 7 Differentiation of three independent phenotypes from one phenotype of a species at a moment. The figure is a special case of judging whether the phenotypic differentiates completely. There is a continuous and a little difference for the phenotype ?1 within the species (the difference range is ± 0.15), and the phenotype 0 versa. The discontinuity of these two interspecies phenotypes is 1, and the discontinuity is much higher than the intraspecies continuous difference. Therefore, it can be considered that the phenotype meets the requirements for phenotype in the concept of morphological species. Taking into account the differentiation of another state of this species, if the difference satisfies the same conditions, the corresponding individuals can be determined as a new species.
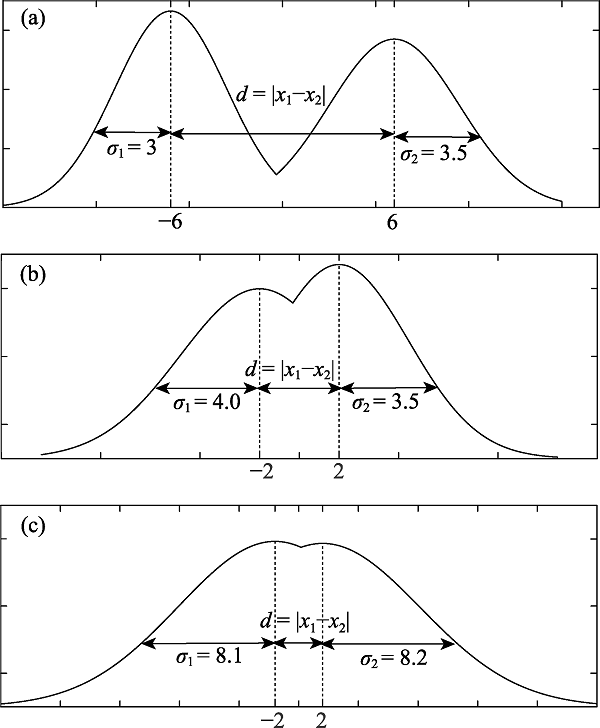
图8 不同的表型分化情形。如果已有一个生物学特征满足定义中的差异性关系, 再考虑性状这一特征。图(a)中, $d=|{{x}_{1}}-{{x}_{2}}|=|(-6)-6|=12,{{\sigma }_{1}}=3,{{\sigma }_{2}}=3.5,d>{{\sigma }_{1}}$且$d>{{\sigma }_{\mathbf{2}}}$, 若有表型分化满足此条件, 则对应个体属于不同物种; 图(b)中, $d=|{{x}_{1}}-{{x}_{2}}|=|(-2)-2|=4,{{\sigma }_{1}}=4,{{\sigma }_{2}}=3.5,d={{\sigma }_{1}}$且$d>{{\sigma }_{\mathbf{2}}}$, 若有表型分化满足此条件, 则对应个体属于同一物种; 图(c)中,$d=|{{x}_{1}}-{{x}_{2}}|=|(-2)-2|=4,{{\sigma }_{1}}=8.1,\ {{\sigma }_{2}}=8.2,$ $d<{{\sigma }_{1}}$且$d<{{\sigma }_{2}}$, 若有表型分化满足此条件, 则对应个体属于同一物种。
Fig. 8 Different case of phenotype differentiation. Imagine the relation of difference has been satisfied for one biological character. Then we consider phenotypes. In figure 8(a), $d=|{{x}_{1}}-{{x}_{2}}|=|(-6)-6|=12,{{\sigma }_{1}}=3,{{\sigma }_{2}}=3.5,d>{{\sigma }_{1}}$ and $d>{{\sigma }_{2}}$. If individuals in two populations satisfy these conditions, they are different species. In figure 8(b), $d=|{{x}_{1}}-{{x}_{2}}|=|(-2)-2|=4,{{\sigma }_{1}}=4,{{\sigma }_{2}}=3.5,d={{\sigma }_{1}}$ and $d>{{\sigma }_{2}}$. If those individuals satisfy these conditions, they are the same species. In figure 8(c), $d=|{{x}_{1}}-{{x}_{2}}|=|(-2)-2|=4,{{\sigma }_{1}}=8.1,{{\sigma }_{2}}=8.2,d<{{\sigma }_{1}}$ and $d<{{\sigma }_{2}}$. If those individuals satisfy these conditions, they are the same species.
| [1] |
Abbott R, Albach D, Ansell S, Arntzen JW, Baird SJE, Bierne N, Boughman J, Brelsford A, Buerkle CA, Buggs R, Butlin RK, Dieckmann U, Eroukhmanoff F, Grill A, Cahan SH, Hermansen JS, Hewitt G, Hudson AG, Jiggins C, Jones J, Keller B, Marczewski T, Mallet J, Martinez-Rodriguez P, Möst M, Mullen S, Nichols R, Nolte AW, Parisod C, Pfennig K, Rice AM, Ritchie MG, Seifert B, Smadja CM, Stelkens R, Szymura JM, Väinölä R, Wolf JBW, Zinner D (2013) Hybridization and speciation. Journal of Evolutionary Biology, 26,229-246.
URL PMID |
| [2] | Darwin C (1929) The origin of species by means of natural selection. American Anthropologist, 61,176-177. |
| [3] |
Frank SA (2011) Natural selection. II. Developmental variability and evolutionary rate. Journal of Evolutionary Biology, 24,2310-2320.
URL PMID |
| [4] | Hedberg O (1958) The taxonomic treatment of vicarious taxa. Uppsala Universitets Arsskrift, 6,186-195. |
| [5] | Hong DY (2016) Biodiversity pursuits need a scientific and operative species concept. Biodiversity Science, 24,979-999. (in Chinese with English abstract) |
| [ 洪德元 (2016) 生物多样性事业需要科学、可操作的物种概念. 生物多样性, 24,979-999.]. | |
| [6] | Li QJ, Li Y (2010) Lamarck redux—A revisit of Darwinism. Journal of Biology, 27(2),55-57. (in Chinese with English abstract) |
| [ 李启剑, 李越 (2010) 拉马克的归来: 对达尔文主义的再审视. 生物学杂志, 27(2),55-57.]. | |
| [7] | Liu JQ (2016) “The integrative species concept” and “species on the speciation way”. Biodiversity Science, 24,1004-1008. (in Chinese with English abstract) |
| [ 刘建全 (2016) “整合物种概念”和“分化路上的物种”. 生物多样性, 24,1004-1008.]. | |
| [8] | Lu BR, Wang Z (2016) What is a species: Conflict between evolutionary continuity and taxonomic discontinuity. Chinese Science Bulletin, 61,2663-2669. (in Chinese with English abstract) |
| [ 卢宝荣, 王哲 (2016) 什么是物种: 进化连续性与分类间断性冲突的产物. 科学通报, 61,2663-2669.]. | |
| [9] | Maynard Smith J (1982) Evolution and the theory of games. Cambridge University Press, Cambridge. |
| [10] | Traulsen A, Iwasa Y, Nowak MA (2007) The fastest evolutionary trajectory. Journal of Theoretical Biology, 249,617-623. |
| [11] | Nowak MA (2006) Evolutionary Dynamics:Exploring the Equations of Life. Harvard University Press, Cambridge. |
| [12] | Wang RW (2021) The End of Rationality and Selfness—A Story on the Asymmetry, Uncertainty and Evolution of Cooperation. China Commerce and Trade Press, Beijing. (in Chinese) (in press) |
| 王瑞武 (2021) 理性与自私的终结——非对称性、不确定性与社会合作行为. 中国商务出版社, 北京. | |
| [13] | Wilkins JS (2009) Species: A History of the Idea. University of California Press, Berkeley. |
| [14] | Zhou CF, Yang G (2011) Existence and Definition of Species. Science Press, Beijing. (in Chinese) |
| 周长发, 杨光 (2011) 物种的存在与定义. 科学出版社, 北京. |
| [1] | 梁健超, 陈淑甜, 何文祥, 熊春妮, 许镇洲, 黄源欣, 刘源志弘, 张浪, 胡慧建. 广州市陆生野生脊椎动物资源——编目、分布与保护[J]. 生物多样性, 2026, 34(5): 25422-. |
| [2] | 张海龙, 聂海燕, 刘萌萌, 魏美才, 李泽建. 浙江乌岩岭叶蜂多样性分布及影响因素[J]. 生物多样性, 2026, 34(5): 25459-. |
| [3] | 周丽洁, 郝珉辉, 何怀江, 程艳霞, 张春雨, 赵秀海. 小兴安岭森林β多样性格局、组分及其影响因素[J]. 生物多样性, 2026, 34(4): 25443-. |
| [4] | 张德喜, 周倩, 裴雪竹, 张红勇, 裴应泰, 孙杉, 刘名信, 张立勋, 赵长明. 甘肃兴隆山森林节肢动物标本照片与DNA条形码数据集[J]. 生物多样性, 2026, 34(4): 25409-. |
| [5] | 黄莉, 刘志发, 龚粤宁, 李步杭, 张健. 广东南岭16个1 ha森林动态监测样地植物群落组成与结构[J]. 生物多样性, 2026, 34(3): 25357-. |
| [6] | 夏东坡, 李静, 田军东, 黄中豪, 伍乘风, 陈仕望, 李进华. 中国野生猕猴属动物的研究进展[J]. 生物多样性, 2026, 34(3): 25460-. |
| [7] | 程晓帆, 李青媛, 李媛辉, 张明祥. 外来入侵物种治理政策体系的困境与出路[J]. 生物多样性, 2026, 34(2): 25332-. |
| [8] | 高雯琪, 向景荣, 赵耀, 范灵霜, 谷圆, 邵韦涵, 李高俊, 赵光军, 陈明斌, 蔡杏伟, 陈凯. 海南热带雨林国家公园黎母山和尖峰岭溪流鱼类群落特征及其对土地利用的响应[J]. 生物多样性, 2026, 34(2): 25374-. |
| [9] | 章旭日, 罗标, 赵彤, 黄丹, 艾为明. 浙江鱼类多样性: 编目、分布与保护[J]. 生物多样性, 2026, 34(2): 25225-. |
| [10] | 周婷婷, 张浩淼, 王浩杰, 廖健, 陈晓丹, 杨楠, 韩博平, 刘振元. 广东省蜻蜓目物种多样性空间格局及关键影响因素[J]. 生物多样性, 2026, 34(2): 25337-. |
| [11] | 周铝, 姚世贸, 赵战合, 郭画, 田成. 庞泉沟国家级自然保护区狍和野猪时空共存格局[J]. 生物多样性, 2026, 34(2): 25327-. |
| [12] | 罗熙, 王啟芳, 王建佳, 杨倩曦, 方鸿飞, 洪淼, 张琦, 蔡灵, 周曦杰, 黄丁勇, 郑新庆. 福建东山海域新记录种揭示我国北缘造礁石珊瑚群落多样性[J]. 生物多样性, 2026, 34(2): 25335-. |
| [13] | 程家劲, 马仁义, 毛康珊. 垂枝柏复合体的物种界定[J]. 生物多样性, 2026, 34(2): 25373-. |
| [14] | 邹怡. α多样性指数选择: 不等量采样下的模拟比较[J]. 生物多样性, 2026, 34(1): 25278-. |
| [15] | 谷际岐, 赖江山, 王瑛, 吴浩然, 张雪, 宋晓彤, 邵小明, 娄安如. 联合物种分布模型与生物群落层次建模框架: 生态学理论、方法及应用[J]. 生物多样性, 2026, 34(1): 25364-. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
备案号:京ICP备16067583号-7
Copyright © 2026 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn
![]()