
生物多样性 ›› 2008, Vol. 16 ›› Issue (4): 353-361. DOI: 10.3724/SP.J.1003.2008.08031 cstr: 32101.14.SP.J.1003.2008.08031
孙华之1, 谭敦炎1,*( ), 曲荣明2
), 曲荣明2
收稿日期:2008-02-02
接受日期:2008-04-13
出版日期:2008-07-20
发布日期:2008-07-20
通讯作者:
谭敦炎
基金资助:
Huazhi Sun1, Dunyan Tan1,*(), Rongming Qu2
Received:2008-02-02
Accepted:2008-04-13
Online:2008-07-20
Published:2008-07-20
Contact:
Dunyan Tan
摘要:
小疮菊(Garhadiolus papposus)是准噶尔荒漠中常见的具异形果实的菊科早春短命植物, 其同一果序中由外到内分别产生外围果、过渡果和中央果3种不同形态和扩散方式的果实。在形态特征上, 每果序中外围果5.24±0.05个, 柱状弧形曲, 深黄、黄至黄白色, 果体长6.67±0.10 mm, 无冠毛, 成熟时被宿存苞片所包被, 果体背面下部与苞片合生, 果皮厚约156.7 µm, 表面较光滑, 瘦果及胚百粒重分别为118.10±1.30 mg和46.70±0.50 mg; 过渡果5.39±0.14个, 柱状弧形曲, 灰白色, 果体长7.60±0.11 mm, 具少量短冠毛, 果皮厚约82.1 µm, 表面有两条纵向细棱, 瘦果及胚百粒重分别为88.30±1.30 mg和36.80±0.30 mg; 中央果5.77±0.13个, 柱形近直立, 深褐色, 果体长9.74±0.14 mm, 具发育完全的冠毛, 果皮厚约69.7 µm, 表面有多条纵向细棱, 瘦果及胚百粒重分别为69.00±0.60 mg和36.90±0.30 mg。在扩散特点上, 3种瘦果的扩散能力表现为中央果>过渡果>外围果, 且散布能力与其形态特征密切相关。中央果成熟后容易从母株上脱落, 发育完全的冠毛能借风力进行远距离扩散; 外围果与宿存苞片紧密相连, 成熟后不易脱落, 散布距离很近; 过渡果的散布特征介于两者之间。本文还讨论了小疮菊异形瘦果的形态特征与扩散特点对其在准噶尔荒漠中成功定居与生长发育以及物种延续和种群扩大、减少同胞果实间竞争等方面的影响。
孙华之, 谭敦炎, 曲荣明 (2008) 短命植物小疮菊异形瘦果特性及其对荒漠环境的适应. 生物多样性, 16, 353-361. DOI: 10.3724/SP.J.1003.2008.08031.
Huazhi Sun, Dunyan Tan, Rongming Qu (2008) Characteristics of heteromorphic achenes of Garhadiolus papposus, an ephemeral Asteraceae species,with reference to their adaptations to desert environment. Biodiversity Science, 16, 353-361. DOI: 10.3724/SP.J.1003.2008.08031.
| 观测指标 Traits | 外围果 Peripheral achenes | 过渡果 Intermediate achenes | 中央果 Central achenes | F | P |
|---|---|---|---|---|---|
| 形状 Shape | 柱状, 弧形曲 Columnar and arcuate | 柱状, 弧形曲 Columnar and arcuate | 柱状, 弧形曲或近直立 Columnar and arcuate or straight | - | - |
| Color | 深黄、黄至黄白色 Dark yellow, yellow or yellow whitish | 灰白色 Offwhite | 深褐色 Dark brown | - | - |
| 附属结构 Additional structure | 苞片膨大宿存紧包在瘦果外, 苞片外侧具短刺; 果喙短, 无冠毛 Phyllary possessing short spine subtends achene closely with short fruit beak and without pappus | 果喙较长, 具少量短冠毛 Elongated fruit beak with a few short pappus | 果喙细长, 具发育完全的冠毛 Elongated fruit beak with developed pappus | - | - |
| 果体长 Length of achene (mm) | 6.67 ± 0.10 Cc | 7.60 ± 0.11 Bb | 9.74 ± 0.14 Aa | 176.95 | 0.00 |
| 背腹宽 Width from back to face (mm) | 0.88 ± 0.03 Aa | 0.60 ± 0.01 Bb | 0.46 ± 0.01 Cc | 103.44 | 0.00 |
| 两侧宽 Width from side to side (mm) | 0.92 ± 0.02 Aa | 0.57 ± 0.01 Bb | 0.41 ± 0.01 Cc | 202.04 | 0.00 |
| 果喙长 Length of beak (mm) | 3.02 ± 0.07 Cc | 3.89 ± 0.78 Bb | 5.49 ± 0.09 Aa | 245.43 | 0.00 |
| 冠毛数量 Number of pappus | - | 7.14 ± 0.61 Bb | 17.36 ± 0.53 Aa | 349.94 | 0.00 |
| 冠毛长 Length of pappus (mm) | - | 0.35 ± 0.02 Bb | 1.02 ± 0.03 Aa | 503.31 | 0.00 |
表1 小疮菊3种瘦果的形态特征比较(不同小写字母表示3种瘦果在0.05的水平上存在差异; 不同大写字母表示3种瘦果在0.01的水平上存在差异)
Table 1 The comparison of characteristics in three types of achenes of Garhadiolus papposus. Different small letters mean difference at 0.05 level, and different capital letters mean difference at 0.01 level
| 观测指标 Traits | 外围果 Peripheral achenes | 过渡果 Intermediate achenes | 中央果 Central achenes | F | P |
|---|---|---|---|---|---|
| 形状 Shape | 柱状, 弧形曲 Columnar and arcuate | 柱状, 弧形曲 Columnar and arcuate | 柱状, 弧形曲或近直立 Columnar and arcuate or straight | - | - |
| Color | 深黄、黄至黄白色 Dark yellow, yellow or yellow whitish | 灰白色 Offwhite | 深褐色 Dark brown | - | - |
| 附属结构 Additional structure | 苞片膨大宿存紧包在瘦果外, 苞片外侧具短刺; 果喙短, 无冠毛 Phyllary possessing short spine subtends achene closely with short fruit beak and without pappus | 果喙较长, 具少量短冠毛 Elongated fruit beak with a few short pappus | 果喙细长, 具发育完全的冠毛 Elongated fruit beak with developed pappus | - | - |
| 果体长 Length of achene (mm) | 6.67 ± 0.10 Cc | 7.60 ± 0.11 Bb | 9.74 ± 0.14 Aa | 176.95 | 0.00 |
| 背腹宽 Width from back to face (mm) | 0.88 ± 0.03 Aa | 0.60 ± 0.01 Bb | 0.46 ± 0.01 Cc | 103.44 | 0.00 |
| 两侧宽 Width from side to side (mm) | 0.92 ± 0.02 Aa | 0.57 ± 0.01 Bb | 0.41 ± 0.01 Cc | 202.04 | 0.00 |
| 果喙长 Length of beak (mm) | 3.02 ± 0.07 Cc | 3.89 ± 0.78 Bb | 5.49 ± 0.09 Aa | 245.43 | 0.00 |
| 冠毛数量 Number of pappus | - | 7.14 ± 0.61 Bb | 17.36 ± 0.53 Aa | 349.94 | 0.00 |
| 冠毛长 Length of pappus (mm) | - | 0.35 ± 0.02 Bb | 1.02 ± 0.03 Aa | 503.31 | 0.00 |

图2 小疮菊3种瘦果果皮微形态特征(柱状横线为比例尺, 为300 µm)。(A) 外围果; (B) 过渡果; (C) 中央果。
Fig. 2 A micrograph taken with a scanning electron microscope, showing the surface ornamentation in three types of achenes of Garhadiolus papposus. (A) Peripheral achene; (B) Intermediate achene; (C) Central achene. Bars = 300 µm.
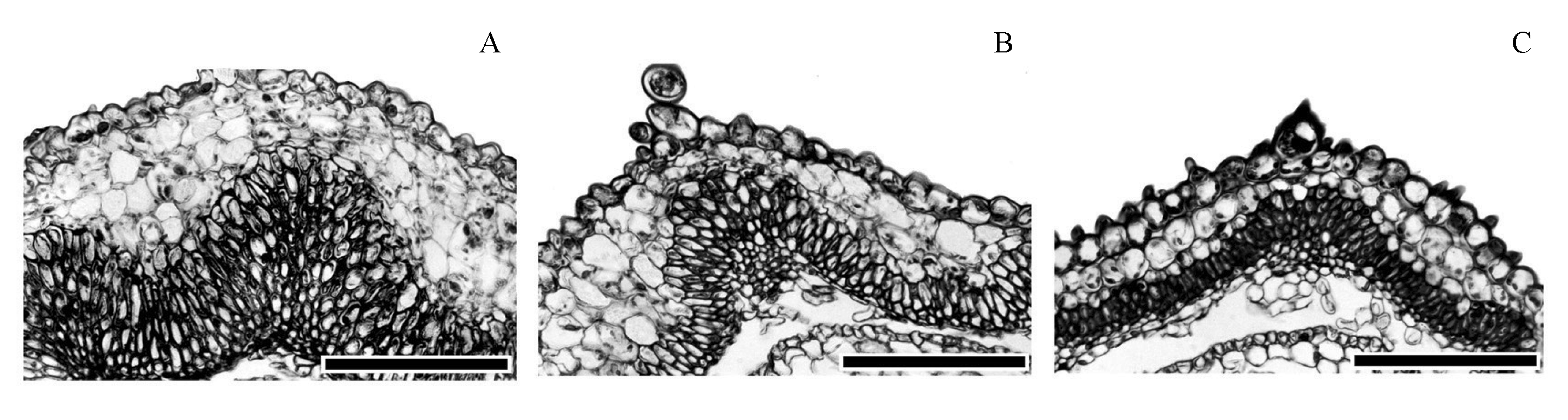
图3 小疮菊3种瘦果的果皮解剖结构特征(比例尺为100 µm)。(A) 外围果; (B) 过渡果; (C) 中央果。
Fig. 3 Pericarp structures in three types of achenes in Garhadiolus papposus. (A) Peripheral achene; (B) Intermediate achene; (C) Central achene. Bars = 100 µm.
| 测量指标 Parameter | 外围果 Peripheral achene | 过渡果 Intermediate achene | 中央果 Central achene | F | P |
|---|---|---|---|---|---|
| 每果序中瘦果数(个) Number of achenes per infructescence | 5.24±0.05Bb | 5.39± 0.14Bb | 5.77 ±0.13 Aa | 5.60 | 0.00 |
| 果实百粒重(mg) Mass per 100 achenes | 118.10± 1.30Aa | 88.30± 1.30Bb | 69.00± 0.60Bb | 357.22 | 0.00 |
| 胚百粒重(mg) Mass per 100 embryos | 46.70± 0.50Aa | 36.80 ±0.30 Bb | 36.90± 0.30Bb | 159.44 | 0.00 |
| 扩散单元百粒重(mg) Mass per 100 dispersal units | 252.70 ±3.40 Aa | 88.30 ± 1.30 Bb | 69.00 ±0.60 Cc | 2,511.48 | 0.00 |
| 果皮/瘦果质量比(%±SE) Mass ratio of pericarp/achene | 60.46± 0.22Aa | 58.29 ±0. 60 Aa | 46.46±1.33Bb | 159.56 | 0.00 |
表2 小疮菊异形瘦果的数量与质量特征(小写字母不同表示变量间在0.05水平上存在差异; 大写字母不同表示变量间在0.01水平上存在差异)
Table 2 Quantity and quality characteristics of three types of achenes in Garhadiolus papposus. Different small letters mean difference at 0.05 level, and different capital letters mean difference at 0.01 level.
| 测量指标 Parameter | 外围果 Peripheral achene | 过渡果 Intermediate achene | 中央果 Central achene | F | P |
|---|---|---|---|---|---|
| 每果序中瘦果数(个) Number of achenes per infructescence | 5.24±0.05Bb | 5.39± 0.14Bb | 5.77 ±0.13 Aa | 5.60 | 0.00 |
| 果实百粒重(mg) Mass per 100 achenes | 118.10± 1.30Aa | 88.30± 1.30Bb | 69.00± 0.60Bb | 357.22 | 0.00 |
| 胚百粒重(mg) Mass per 100 embryos | 46.70± 0.50Aa | 36.80 ±0.30 Bb | 36.90± 0.30Bb | 159.44 | 0.00 |
| 扩散单元百粒重(mg) Mass per 100 dispersal units | 252.70 ±3.40 Aa | 88.30 ± 1.30 Bb | 69.00 ±0.60 Cc | 2,511.48 | 0.00 |
| 果皮/瘦果质量比(%±SE) Mass ratio of pericarp/achene | 60.46± 0.22Aa | 58.29 ±0. 60 Aa | 46.46±1.33Bb | 159.56 | 0.00 |
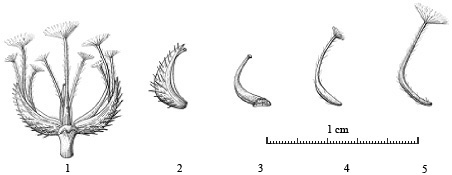
图1 小疮菊3种瘦果的形态特征。1: 果序(纵剖面), 示3种瘦果在果序上的排列; 2: 被宿存苞片包被的外围果; 3: 去除苞片后的外围果; 4: 过渡果; 5: 中央果(谭黎霞绘)
Fig. 1 Morphology of three types of achenes in Garhadiolus papposus. 1, Infructescence (longitudinal section); 2, Peripheral achene subtended by phyllary; 3, Peripheral achene without phyllary; 4, Intermediate achene; 5, Central achene (drawn by Lixia Tan)
| [1] | An ZX (安争夕), Shen GM (沈观冕), Zhai DT (翟大彤) (1999) Asteraceae. In:Flora Xinjiangensis (新疆植物志). Tomus 5,p. 371. Xinjiang Science, Technology and Hygiene Publishing House, Urumqi. (in Chinese) |
| [2] |
Arroyo MTK, Chacon P, Cavieres LA (2006) Relationship between seed bank expression, adult longevity and aridity in species of Chaetanthera (Asteraceae) in central Chile. Annals of Botany, 98,591-600.
DOI URL PMID |
| [3] | Bachmann K, Chambers KL, Price HJ (1984) Genetic components of heterocarpy in Microseries hybrid B87 (Asteraceae, Lactuceae ) . Plant Systematics and Evolution, 148,149-164. |
| [4] | Baskin JM, Baskin CC (1976) Germination dimorphism in Heterotheca subaxillaris var . subaxillari. Bulletin of the Torrey Botanical Club, 103,201-206. |
| [5] | Berger A (1985) Seed dimorphism and germination behaviour in Salicornia patula. Vegetatio, 61,137-143. |
| [6] | Brändel M (2004) Dormancy and germination of heteromorphic achenes of Bidens frondosa. Flora, 199,228-233. |
| [7] |
Brändel M (2007) Ecology of achene dimorphism in Leontodon saxatilis. Annals of Botany, 100,1189-1197.
DOI URL PMID |
| [8] | Ellner SP, Shmida A (1984) Seed dispersal in relation to habitat in the genus Picris (Compositae) in Mediterranean and arid regions. Israel Journal of Botany, 33,25-39. |
| [9] | Eriksson A, Eriksson O (1997) Seedling recruitment in semi-natural pastures: the effects of disturbance, seed size, phenology and seed bank. Nordic Journal of Botany, 17,469-482. |
| [10] | Forsyth C, Brown NAC (1982) Germination of the dimorphic fruits of Bidens pilosa L. New Phytologist, 90,151-164. |
| [11] | Gibson JP (2001) Ecological and genetic comparison between ray and disc achene pools of the heteromorphic species Prionopsis ciliata (Asteraceae). International Journal of Plant Sciences, 162,137-145. |
| [12] | Gravuer K, von Wettberg EJ, Schmitt J (2003) Dispersal biology of Liatris scariosa var . novae-angliae (Asteraceae), a rare New England grassland perennial. American Journal of Botany, 90,1159-1167. |
| [13] | Harper JL, Obeid M (1967) Influence of seed size and depth of sowing on the establishment and growth of varieties of fiber and oil seed flax. Crop Science, 7,527-532. |
| [14] | Harper JL (1977) Population Biology of Plants. Academic Press, New York. |
| [15] | Imbert E (1999) The effects of achene dimorphism on the dispersal in time and space in Crepis sancta (Asteraceae). Canadian Journal of Botany, 77,508-513. |
| [16] |
Imbert E (2001) Capitulum characters in a seed heteromorphic plant, Crepis sancta (Asteraceae): variance partitioning and inference for the evolution of dispersal rate. Heredity, 86,78-86.
DOI URL PMID |
| [17] | Imbert E (2002) Ecological consequences and ontogeny of seed heteromorphism. Perspectives in Plant Ecology,Evolution and Systematics, 5,13-36. |
| [18] | Koller D, Roth N (1964) Studies on the ecological and physiological significance of amphicarpy in Gyminarrhena micrantha (Compositae). American Journal of Botany, 51,26-35. |
| [19] | Mandák B, Pyśek P (2001) Fruit dispersal and seed banks in Atriplex sagittata: the role of heterocarpy . Journal of Ecology, 89,159-165. |
| [20] | Mao ZM (毛祖美), Zhang DM (张佃民) (1994) The conspectus of ephemeral flora in northern Xinjiang. Arid Zone Research (干旱区研究), 11(3),1-26. (in Chinese) |
| [21] | Maxwell CD, Zobel A, Woodfine D (1994) Somatic polymorphism in the achenes of Tragopogon dubious. Canadian Journal of Botany, 72,1282-1288. |
| [22] | McDonough WT (1975) Germination polymorphism in Grindelia squarrosa. Northwest Science, 49,190-200. |
| [23] |
McEvoy PB (1984) Dormancy and dispersal in dimorphic achenes of tansy ragwort, Senecio jacobaea L. (Compositae). Oecologia, 61,160-168.
DOI URL PMID |
| [24] | Rocha OJ (1996) The effects of achene heteromorphism on the dispersal capacity of Bidens pilosa L. International Journal of Plant Sciences, 157,316-322. |
| [25] | Ruiz de Clavijo E (1994) Heterocarpy and seed polymorphism in Ceratocapons heterocarpa (Fumariaceae). International Journal of Plant Sciences, 155,196-202. |
| [26] | Ruiz de Clavijo E, Jiménez MJ (1998) The influence of achene type and plant density on growth and biomass allocation in the heterocarpic annual Catananche lutea (Asteraceae). International Journal of Plant Sciences, 159,637-647. |
| [27] | Shi Z (石铸) (1997) Asteraceae. In: Flora Reipublicae Popularis Sinicae (中国植物志), Tomus 80(1), pp. 260-265. Science Press, Beijing. (in Chinese) |
| [28] | Stebbins GJ (1974) Flowering Plants Evolution Above the Species Level. Harvard University Press,Cambridge, Massachusetts. |
| [29] | Tanowitz BD, Salopek PF, Mahall BE (1987) Differential germination of ray and disc achenes in Hemizonia increscens (Asteraceae). American Journal of Botany, 74,303-312. |
| [30] | Telenius A (1992) Seed heteromorphism in a population of Spergularia media in relation to the ambient vegetation density. Acta Botanica Neerlandica, 41,305-318. |
| [31] |
Telenius A, Torstensson P (1989) The seed dimorphism of Spergularia marina in relation to dispersal by wind and water. Oecologia, 80,206-210.
DOI URL PMID |
| [32] | Travesetet AN, Riera A, Mas RE (2001) Ecology of fruit-colour polymorphism in Myrtus communis and differential effects of birds and mammals on seed germination and seedling growth. Journal of Ecology, 89,749-760. |
| [33] | Ungar IA (1979) Seed dimorphism in Salicornia europaea L. Botanical Gazette, 140,102-108. |
| [34] | van Mölken T, Jorritsma-Wienk LD, van Hoek PHW, de Kroon H (2005) Only seed size matters for germination in different populations of the dimorphic Tragopogon pratensis subsp. pratensis (Asteraceae). American Journal of Botany, 92,432-437. |
| [35] | Venable DL (1985) The evolutionary ecology of seed heteromorphism. The American Naturalist, 126,577-595. |
| [36] | Venable DL, Burquez AM, Corral G, Morales E, Espinosa F (1987) The ecology of seed heteromorphism in Heterosperma pinnatum in central Mexico. Ecology, 68,65-76. |
| [37] |
Venable DL, Lawlor L (1980) Delayed germination and dispersal in desert annuals: escape in space and time. Oecologia, 46,272-282.
DOI URL PMID |
| [38] |
Venable DL, Levin DA (1985) Ecology of achene dimorphism in Heterotheca latifolia. I. Achene structure, germination and dispersal. Journal of Ecology, 73,113-145.
DOI URL |
| [39] |
Zhang J (1993) Seed dimorphism in relation to germination and growth of Cakile edentula. Canadian Journal of Botany, 71,1231-1235.
DOI URL |
| [40] | Zhao XJ (赵学杰), Tan DY (谭敦炎), Li XR (李新蓉) (2008) The breeding system of two ephemeral species of Lappula, with reference to their adaptations to the Junggar Desert environment, northwest China. Journal of Desert Research (中国沙漠), 28,294-300. (in Chinese with English abstract) |
| [41] | Zheng GC (郑国锠), Gu ZP (谷祝平) (1993) Biological Microtechnology(生物显微技术). China Higher Education Press,Beijing. (in Chinese) |
| [42] | Zheng JM (郑景明), Sang WG (桑卫国), Ma KP (马克平) (2004) Advances in model construction of anemochoric seed long-distance dispersal. Acta Phytoecologica Sinica(植物生态学报), 28,414-425. (in Chinese with English abstract) |
| [43] | Zhou TY (周太炎), Guo RL (郭荣麟), Lan YZ (蓝永珍), Lu LL (陆莲立) (1987) Cruciferae. In:Flora Reipublicae Popularis Sinicae(中国植物志), Tomus 33, pp.351-353. Science Press,Beijing. (in Chinese) |
| [44] | Zhu SX (朱世新) (2004) Systematics of Chaetoseris and Stenoseris (Compositae-Lactuceae) (毛鳞菊属和细莴苣属的系统学研究). PhD dissertation, Institute of Botany, Chinese Academy of Sciences, Beijing. (in Chinese with English abstract) |
| [1] | 王林, 尹梓杨, 黄慧芳, 王静. 基于Carter-Morley Jones蛋形模型的参数估计新方法[J]. 生物多样性, 2025, 33(1): 24203-. |
| [2] | 陈静, 张丙昌, 刘燕晋, 武杰, 赵康, 明姣. 荒漠生物结皮细鞘丝藻类(Leptolyngbya-like)蓝藻多样性[J]. 生物多样性, 2024, 32(9): 24186-. |
| [3] | 王江, 赵一凡, 屈彦福, 张财文, 张亮, 陈传武, 王彦平. 中国蛇类形态、生活史和生态学特征数据集[J]. 生物多样性, 2023, 31(7): 23126-. |
| [4] | 张世航, 陶冶, 陈玉森, 郭浩, 陆永兴, 郭星, 刘朝红, 周晓兵, 张元明. 准噶尔荒漠土壤多功能性的空间变异特征及其驱动因素[J]. 生物多样性, 2022, 30(8): 22097-. |
| [5] | 钟雨茜, 陈传武, 王彦平. 中国蜥蜴类生活史和生态学特征数据集[J]. 生物多样性, 2022, 30(4): 22071-. |
| [6] | 丁晨晨, 梁冬妮, 信文培, 李春旺, 蒋志刚. 中国哺乳动物形态、生活史和生态学特征数据集[J]. 生物多样性, 2022, 30(2): 21520-. |
| [7] | 王彦平, 宋云枫, 钟雨茜, 陈传武, 赵郁豪, 曾頔, 吴亦如, 丁平. 中国鸟类的生活史和生态学特征数据集[J]. 生物多样性, 2021, 29(9): 1149-1153. |
| [8] | 俞正森, 宋娜, 本村浩之, 高天翔. 中国银口天竺鲷属鱼类的分类厘定[J]. 生物多样性, 2021, 29(7): 971-979. |
| [9] | 郭志文, 郑景明. 用植物生活史性状预测种子扩散方式[J]. 生物多样性, 2017, 25(9): 966-971. |
| [10] | 郑硕理, 田晓玲, 黄承玲, 王灵军, 冯元, 张敬丽. 结合分子手段和形态分析验证大白杜鹃与马缨杜鹃的自然杂交[J]. 生物多样性, 2017, 25(6): 627-637. |
| [11] | 王雨, 张会勇, 项鹏, 叶又茵, 林更铭, 杨清良, 林茂. 颗石藻颗石粒形态的原子力显微观测方法: 以赫氏艾密里藻为例[J]. 生物多样性, 2016, 24(7): 847-854. |
| [12] | 魏雪苹, 张宪春. 蕨类植物不同孢子裂缝类型在中国的分布格局[J]. 生物多样性, 2016, 24(10): 1129-1134. |
| [13] | 姚蓓, 余建平, 刘晓娟, 米湘成, 马克平. 亚热带常绿阔叶林种子性状对木本植物聚集格局的影响[J]. 生物多样性, 2015, 23(2): 157-166. |
| [14] | 钟云芳, 张哲, 宋希强, 周兆德. 海南凤仙花不同海拔种群的传粉生物学[J]. 生物多样性, 2014, 22(4): 467-475. |
| [15] | 徐兆礼. 中国近海浮游动物多样性研究的过去和未来[J]. 生物多样性, 2011, 19(6): 635-645. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
备案号:京ICP备16067583号-7
Copyright © 2026 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn
![]()