
Biodiv Sci ›› 2013, Vol. 21 ›› Issue (3): 249-259. DOI: 10.3724/SP.J.1003.2013.11240 cstr: 32101.14.SP.J.1003.2013.11240
Special Issue: 生物多样性与生态系统功能; 生物入侵
• Orginal Article • Previous Articles Next Articles
Sikai Wang, Qiang Sheng, Tianjiang Chu, Bo Li, Jiakuan Chen, Jihua Wu*( )
)
Received:2012-12-05
Accepted:2013-02-25
Online:2013-05-20
Published:2013-06-05
Contact:
Wu Jihua
Sikai Wang,Qiang Sheng,Tianjiang Chu,Bo Li,Jiakuan Chen,Jihua Wu. Impact of invasive plants on food webs and pathways[J]. Biodiv Sci, 2013, 21(3): 249-259.
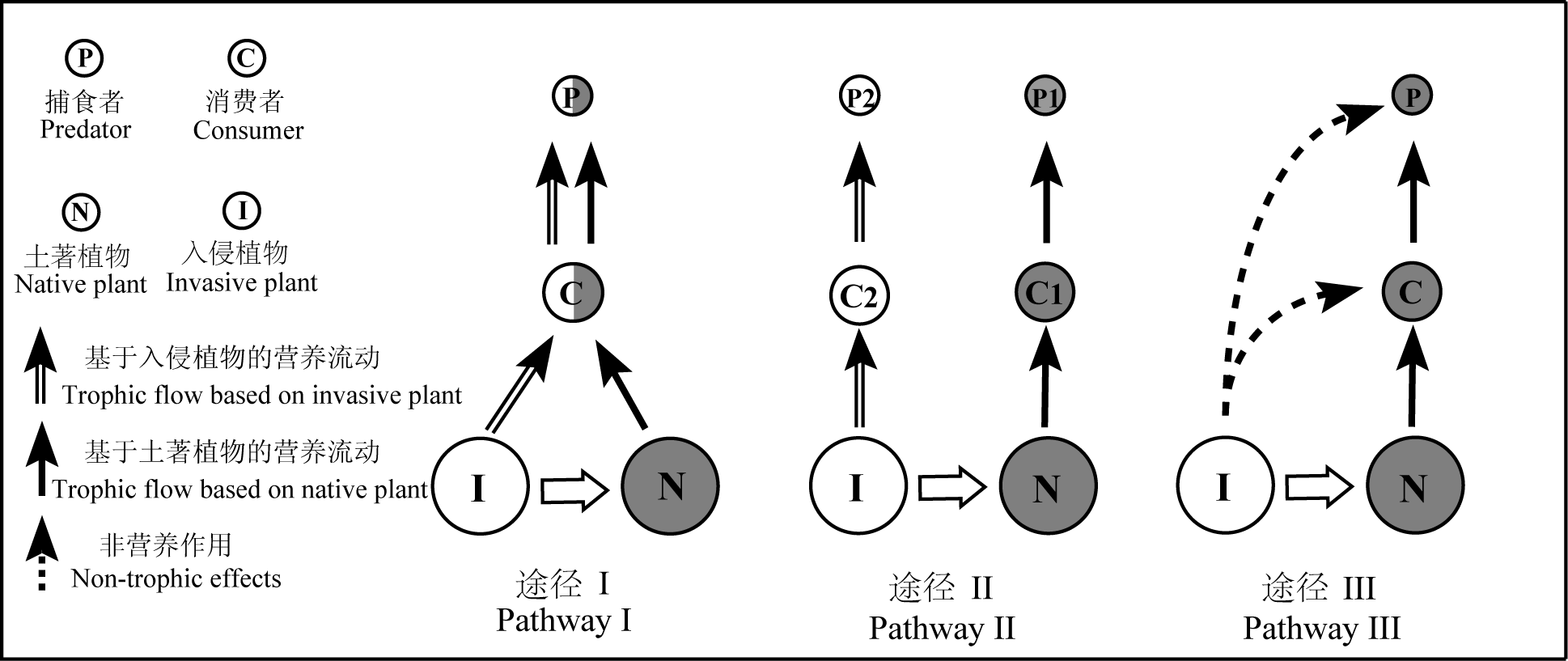
Fig. 1 Three influence pathways of plant invasion on food webs. Pathway I represents that the invasive plant could be incorporated into the native food web. Pathway II represents that the invasive plants change the original path and generate a new energy channel. Pathway III shows the non-trophic effects of invasive plant on food webs. C1 and P1 represent native consumer and predator; C2 and P2 represent new consumer and predator. The gray filled circles and the empty circles represent the components of food web before and after plant invasion, respectively.
| 生态系统 Ecosystem | 地点 Sites | 入侵物种 Invasive species | 影响结果 Effects | 途径 Pathway | 参考文献 References |
|---|---|---|---|---|---|
| 盐沼 | 中国长江口 | 互花米草 Spartina alterni?ora | 入侵植物碎屑通过土著细菌性线虫的取食, 进入到土著食物网中。 | I | Chen et al., 2007 |
| 盐沼 | 中国长江口 | 互花米草 S. alterni?ora | 互花米草对鮻鱼(Chelon haematocheilus)和斑尾复鰕虎鱼(Synechogobius ommaturus)的食物贡献率超过50%, 通过游泳动物的取食进入到土著食物网中。 | I | Quan et al., 2007 |
| 盐沼 | 中国长江口 | 互花米草 S. alterni?ora | 互花米草能够通过直翅目、双翅目等广食性草食者的取食进入到土著食物网中(途径I); 而部分专食性草食者由于不能取食互花米草而离开(途径II)。 | I、II | Wu et al., 2009 |
| 盐沼 | 中国长江口 | 互花米草 S. alterni?ora | 互花米草生境中具有更高的土壤含水量和更低的光照强度, 能够为土著消费者无齿相手蟹(Sesarma dehaani)提供阴凉潮湿的环境, 有效地缓解了失水胁迫, 从而对土著食物网中的群落结构和物种组成产生影响。 | III | Wang et al., 2008 |
| 盐沼 | 西班牙西北部加利西亚 | 狐米草 S. patens | 入侵植物不能被土著草食性昆虫取食, 但是可以通过腐食者进入到土著碎屑食物网, 改变了食物网有机质的循环路径。 | II | Page et al., 2010 |
| 盐沼 | 美国新泽西 | 芦苇 Phragmites australis | 芦苇入侵使得原先取食土著植物互花米草的节肢动物改为取食碎屑和底栖微藻。 | II | Gratton & Denno, 2006 |
| 盐沼 | 美国提瓦那河口 | 多枝柽柳 Tamarisk ramosissima | 多枝柽柳碎屑能够被土壤表层大型无脊椎动物取食利用按照原路径进入土著食物网中(途径I); 而土壤下层无脊椎动物不能够利用入侵植物碎屑导致其丰度降低(途径II)。 | I、II | Whitcraft et al., 2008 |
| 盐沼 | 美国旧金山湾 | 互花米草 S. alterni?ora | 入侵植物互花米草能够被光蝉(Prokelisia marginata)等土著广食性草食者取食, 进入到土著食物网中。 | I | Daehler & Strong, 1994 |
| 盐沼 | 美国旧金山湾 | 穗状狐尾藻 Myriophyllum spicatum | 入侵植物复杂的形态结构为一些底栖无脊椎动物或者幼鱼提供了庇护场所, 能够阻止更高级消费者对这些动物的取食。 | III | Chaplin & Valentine, 2009 |
| 盐沼 | 美国旧金山湾 | 互花米草 S. alterni?ora | 入侵植物产生的大量碎屑没有被土著底栖动物取食利用(途径II); 同时入侵植物作为生态系统工程师能够改变植物结构、沉积物性质、光照、水文等环境特点, 对土著生物群落与食物网产生影响(途径III: 植物形态结构、环境理化性质)。 | II、III | Brusati & Grosholz, 2009 |
| 盐沼 | 美国旧金山湾 | 互花米草 S. alterni?ora | 入侵植物不能被土著消费者双壳类白缨蛤(Macoma petaluma)取食利用,导致双壳类消费者丰度与丰富度降低(途径II); 另外互花米草提高盐沼的海拔高度, 降低水流流速与光照渗透强度, 通过生境的变化影响底栖动物食物网(途径III: 环境理化性质)。 | II、III | Brusati & Grosholz, 2007 |
| 盐沼 | 美国东部 | 芦苇 P. australis | 来自芦苇的初级生产力能够进入到土著食物网中, 为河口游泳动物提供食物。 | I | Weis & Weis, 2003 |
| 盐沼 | 法国西北部 | Elymus athericus | 披碱草属植物E. athericus的碎屑分解较慢, 改变了微生境条件, 为蜘蛛提供了适合结网的生境和庇护所。 | III | Petillon et al., 2005 |
| 溪流 | 澳大利亚昆士兰 | 巴拉草 Brachiaria mutica | 入侵植物巴拉草不能被蜉蝣目动物Baetid bymphs、蚋科(Simuliidae)、米虾属(Caridina)、沼虾属(Macrobrachium)等土著水生动物取食, 不能融入到土著水生食物网中。 | II | Bunn et al., 1997 |
| 溪流 | 爱尔兰西部 | 彭土杜鹃 Rhododendron ponticum | 入侵植物彭土杜鹃的可利用性较差, 同时其产生的大量碎屑抑制了藻类的生长, 导致无脊椎动物丰度的降低, 食物来源类型也从基于藻类转向基于颗粒有机质为主, 造成食物网群落结构以及流通路径的改变。 | II | Hladyz et al., 2011 |
| 森林 | 英国雅芳希斯 国家公园 | 北美白珠树 Gaultheria shallon 寻石楠 Calluna vulgaris | 入侵植物能够通过广食性草食者进入到土著食物网(途径I); 而专食性消费者不能适应入侵植物而选择离开入侵区域(途径II)。 | I、II | Carvalheiro et al., 2010 |
| 森林 | 美国芝加哥 Morton植物园 | 金银忍冬 Lonicera maackii 药鼠李 Rhamnus cathartica | 入侵植物的枝条结构适合鸟类筑巢, 也适合捕食者穿行, 使知更鸟鸟巢更容易遭受捕食者的破坏, 繁殖成功率降低, 因此入侵植物通过形态结构的变化对食物网产生了非营养作用。 | III | Schmidt & Whelan, 1999 |
| 森林 | 美国密西根 | 小蔓长春花 Vinca minor | 入侵植物改变了森林地被物的物理特性, 茂密的入侵植物限制了蜘蛛的活动能力, 造成蜘蛛丰度、均匀度和多样性的降低, 进而通过捕食作用影响昆虫食物网结构。 | III | Bultman & DeWitt, 2008 |
| 森林 | 美国 | 多种外来树种 | 鳞翅目昆虫对入侵植物没有形成生理和行为上的适应, 不能够利用外来植物作为营养来源和宿主, 导致生物量和多样性下降, 进而导致更高营养级的食虫动物种类和数量下降。 | II | Tallamy & Shropshire, 2009 |
| 森林 | 美国东部 | 葱芥 Alliaria petiolata | 入侵植物葱芥能够向土壤中渗出植物毒素芥子油苷(glucosinolate), 抑制丛枝菌根真菌的生长, 从而影响到土著植物的生长繁殖。 | III | Stinson et al., 2006 |
| 森林 | 南非 | 金合欢属伞树 Acacia saligna | 一方面入侵植物种子能够被啮齿类纹鼠(Rhabdomys pumilio)大量取食, 从而进入到土著食物网中(途径I); 另一方面入侵植物对生境的改变不适合纹鼠挖洞筑巢, 使纹鼠种群密度降低, 导致食物网中物种组成的改变(途径III: 环境理化性质)。 | I、III | Holmes, 1990 |
| 湖泊 | 美国东北部 | 穗状狐尾藻 Myriophyllum spicatum | 入侵植物能够被鞘翅目动物Euhrychiopsis lecontei取食, 根茎生物量降低一半, 土著消费者通过下行效应抑制了入侵植物的扩散。 | I | Creed & Sheldon, 1995; Sheldon & Creed, 1995 |
| 湖泊 | 美国密歇根 | 穗状狐尾藻 M. spicatum | 穗状狐尾藻为多裂形植物, 能够为附生在植物表面的大型无脊椎动物提供更多的觅食场所以及更多的遮蔽面积, 因而在多裂型植物周围有更多的无脊椎动物密度和生物量。 | III | Cheruvelil et al., 2002 |
| 湖泊 | 新西兰, 瓦那卡湖Wanaka | 卷叶蜈蚣草 Lagarosiphon major 伊乐藻 Elodea canadensis | 入侵区无脊椎动物群落密度和多样性比非入侵区域大, 优势物种发生改变, 食物网碳流通路径从基于沉积颗粒有机质为主转向基于水生植物为主。 | II | Kelly & Hawes, 2005 |
| 红树林 | 美国夏威夷、波多黎各岛 | 美洲红树 Rhizophora mangle | 美洲红树具有较高的单宁含量, 能够破坏许多生物正常的消化吸收功能, 较高的C:N比和木质素含量使得其叶片的营养价值较低。杂食者蓝蟹(Callinectes sapidus)、悬浮取食者贻贝(Mytilus sp.)、表层取食者滨螺(Littoraria sp.)等消费者无法适应这种植物碎屑, 其入侵区消费者种类与数量降低。 | II | Demopoulos et al., 2007 |
| 河流泛滥区 | 新西兰东南部 | 大叶醉鱼草 Buddleja davidii | 入侵植物使得根系周围土壤C、土壤细菌生物量增加, 土壤微生物群落结构发生改变, 土壤食微线虫和肉食性线虫丰度增加, 通过营养关系改变了食物网结构。 | II | Peltzer et al., 2009 |
| 海岸沙滩 | 西班牙西北部Galician海岸 | 海黍子马尾藻 Sargassum muticum | 入侵植物在冬季能够被底栖无脊椎动物取食, 通过途径I进入到土著食物网中; 而在春季由于土著藻类的大量生长繁殖, 入侵植物不能被取食, 导致底栖无脊椎动物的减少。入侵植物在春季通过途径II改变食物网。 | I、II | Rossi et al., 2010 |
| 海岸草地 | 德国波罗的海群岛 | 曲柄藓属植物Campylopus intro?exus | 曲柄藓属植物C. intro?exus入侵区杂草较少, 限制了草食性昆虫的生长, 导致入侵区步甲科物种丰度的降低(途径II); 另外C. intro?exu的形态结构不适合蜘蛛结网, 还通过改变周围生境中的温度和湿度降低了步甲科物种与蜘蛛的丰富度, 通过非营养作用改变食物网的物种组成(途径III: 形态结构与理化性质)。 | II、III | Schirmel et al., 2011 |
| 海岸草地 | 美国加利福尼亚沿岸 | 马德雀麦 Bromus madritensis 裂稃燕麦 Avena barbata | 入侵植物碎屑增加土壤湿度, 促进本地植物的生长, 增加灌丛草食动物叶蝉和捕食者蜘蛛的丰度, 从而对食物网产生影响。 | III | Wolkovich, 2010 |
| 海岸草地 | 美国加利福尼亚沿岸 | 莫邪菊 Carpobrotus edulis | 入侵植物的果实能够被土著小型哺乳动物加利福尼亚兔(Lepus californicus)以及骡鹿(Odocoileus hemionus)取食, 种子能够通过动物粪便传播扩散。 | I | Vila & D'Antonio, 1998 |
| 海岸草地 | 美国加利福尼亚沿岸 | 二穗短柄草、马德雀麦等Brachypodium distachyon B. madritensis etc. | 植物入侵产生的大量凋落物使得土著节肢动物蚂蚁和螨虫等物种的丰度降低, 通过营养级联影响到入侵生态系统食物网结构。 | II | Wolkovich et al., 2009 |
| 岛屿 | 新西兰东南部 | 花葵 Lavatera arborea | 入侵植物成为岛屿陆地生态系统的重要组成部分, 从传粉者到顶级捕食者多个营养级物种都有来自花葵的同位素标记, 入侵植物已经完全融入到土著食物网中。 | I | Hawke & Clark, 2010 |
| 岛屿 | 大西洋北部Azores群岛 | 日本柳杉、波叶海桐等Cryptomeria japonica, Pittosporum undulatum etc. | 植物入侵导致取食土著植物的大型昆虫消失, 外来小型昆虫进入, 形成新的食物网流通路径。 | II | Heleno et al., 2009 |
| 池塘 | 控制实验 | 多种水生植物 | 外来入侵植物能够被土著广食性草食者取食, 通过营养作用进入到土著食物网中。 | I | Parker & Hay, 2005 |
| 池塘 | 控制实验 | 黑藻 Hydrilla verticillata | 植物入侵导致入侵区域的空间异质性降低, 改变了大型无脊椎动物的类群, 造成食物网中物种组成的变化。 | III | Theel et al., 2008 |
| 池塘 | 美国西北部 | 千屈菜和鹬草 Lythrum salicaria and Phalaris arundinacea | 土著草食者飞蛾等不能够取食入侵植物而选择离开入侵区域, 造成入侵区食物网中物种组成的变化。 | II | Schooler et al., 2009 |
| 池塘 | 北美 | 千屈菜 L. salicaria | 入侵植物叶片由于具有较高的单宁含量, 可利用性较低, 不能被土著消费者美洲蟾蜍(Bufo americanus)的蝌蚪取食, 导致入侵区美洲蟾蜍的成活率显著降低。 | II | Brown et al., 2006 |
| 草地 | 美国蒙大拿州 | 斑点矢车菊 Centaurea maculosa | 通过引入矢车菊的天敌果蝇来控制植物入侵, 但是果蝇数量的迅速增加导致其捕食者鹿鼠增加, 鹿鼠又取食土著植物种子,从而形成新的食物网结构。 | II | Ortega et al., 2004 |
| 草地 | 控制实验 | 斑点矢车菊 C. maculosa | 入侵植物较高的植物密度以及复杂的形态结构提高了卷叶蛛(Dictyna)的密度与结网面积, 降低了卷叶蛛的猎物实蝇科昆虫Urophora affinis的密度, 入侵植物通过形态特征的变化以及对环境条件的改变影响昆虫食物网。 | III | Pearson, 2010 |
| 草地 | 北美西部 | 斑点矢车菊 C. maculosa | 矢车菊的形态结构方便蜘蛛结网, 入侵区蜘蛛网大小与捕获量分别是未入侵区的2.9-4倍和2-2.3倍, 入侵植物形态结构通过捕食者产生下行效应, 影响昆虫食物网的物种组成。 | III | Pearson, 2009 |
| 草地 | 澳大利亚北部 | 大含羞草 Mimosa pigra | 在入侵植物含羞草上发现有50科,101属,共114种植食性昆虫, 入侵植物既能够被土著草食者取食进入到土著食物网中(途径I), 也能够被外来草食者利用形成新的食物网结构(途径II)。 | I、II | Wilson et al., 1990 |
| 草地 | 荷兰 | 菊科和十字花科草本植物 | 植物入侵区域虽然根系生物量较多, 但是却无法被土著消费者取食, 导致入侵植物根系周围线虫多样性和生物量显著降低。 | II | Morrien et al., 2012 |
| 草地 | 英国西部 | 凤仙花 Impatiens glandulifera | 外来植物凤仙花具有较高质量的花蜜, 能够吸引较多的传粉者, 入侵区传粉昆虫丰富度与丰度显著提高, 同时也促进了入侵植物的成功授粉。 | III | Lopezaraiza-Mikel et al., 2007 |
| 草地 | 美国犹他州东南部 | 旱雀麦 Bromus tectorum | 线虫、微型节肢动物等土壤动物不能取食入侵植物, 而选择离开入侵区域, 导致土壤食物网结构与物种组成发生变化。 | II | Belnap et al., 2005 |
| 草地 | 波兰南部 | 一枝黄花 Solidago spp. | 入侵植物造成蝴蝶、昆虫等无脊椎动物丰度降低, 引起以这些动物为食物的鸟类种类和数量的减少(途径II)。另外, 入侵植物的形态结构不适合鸟类筑巢并且入侵区缺少鸟类筑巢所需的材料, 导致鸟类繁殖成功率的下降(途径III: 植物形态结构)。 | II、III | Skorka et al., 2010 |
| 草地 | 美国落基山国家公园 | 乳浆大戟和丝路蓟 Euphorbia esula and Cirsium arvense | 入侵植物不能被土著土壤动物取食, 土壤动物由于缺少食物资源而离开入侵区域(途径II);同时植物入侵使得裸地斑块增加, 改变了土壤有机质含量、pH值以及多种微量元素含量, 导致土著土壤食物网中蜱螨目等小型节肢动物丰度在入侵区显著降低(途径III: 理化性质)。 | II、III | Pritekel et al., 2006 |
| 草地 | 控制实验 | 多种外来植物 | 外来入侵植物降低土著昆虫生物量, 通过营养级联作用影响较高营养级的食虫动物丰度, 严重扰乱了入侵区陆地食物网结构。 | II | Tallamy et al., 2010 |
| 水生系统 | 控制实验 | 凤眼蓝、芋等多种水生植物 Eichhornia crassipes, Colocasia esculenta | 土著广食性草食者苹螺(Pomacea sp.)以及Procambarus spiculifer等对外来入侵植物的取食是土著植物的2-17倍, 入侵植物能够被土著系统中的广食性草食者取食利用, 融入土著食物网中, 而广食性草食者的取食抑制了入侵植物的扩散。 | I | Morrison & Hay, 2011 |
| 水生系统 | 综述 | 大型水生植物 | 大型水生植物的入侵导致生境复杂性增加、水体缺氧, 释放化学物质产生异株相克化学作用, 改变土著动植物群落结构及物种组成(途径III: 植物形态特征和环境理化性质); 另一方面能够降低食物质量, 导致土著鱼类和大型无脊椎动物丰度的降低(途径II)。 | II、III | Schultz & Dibble, 2012 |
| 多种系统 | 捷克 | 13种入侵植物 | 入侵植物的株高、覆盖度及根系的密集度等特点改变了入侵区群落结构物种组成与物种丰富度。 | III | Hejda et al., 2009 |
| 多种系统 | 综述 | 100多种入侵植物 | 外来植物对土著草食者还没有形成防御策略, 更容易受到土著草食者的取食, 进入土著食物网。 | I | Parker et al., 2006 |
Appendix I Cases showing influence pathways of invasive plants on food webs
| 生态系统 Ecosystem | 地点 Sites | 入侵物种 Invasive species | 影响结果 Effects | 途径 Pathway | 参考文献 References |
|---|---|---|---|---|---|
| 盐沼 | 中国长江口 | 互花米草 Spartina alterni?ora | 入侵植物碎屑通过土著细菌性线虫的取食, 进入到土著食物网中。 | I | Chen et al., 2007 |
| 盐沼 | 中国长江口 | 互花米草 S. alterni?ora | 互花米草对鮻鱼(Chelon haematocheilus)和斑尾复鰕虎鱼(Synechogobius ommaturus)的食物贡献率超过50%, 通过游泳动物的取食进入到土著食物网中。 | I | Quan et al., 2007 |
| 盐沼 | 中国长江口 | 互花米草 S. alterni?ora | 互花米草能够通过直翅目、双翅目等广食性草食者的取食进入到土著食物网中(途径I); 而部分专食性草食者由于不能取食互花米草而离开(途径II)。 | I、II | Wu et al., 2009 |
| 盐沼 | 中国长江口 | 互花米草 S. alterni?ora | 互花米草生境中具有更高的土壤含水量和更低的光照强度, 能够为土著消费者无齿相手蟹(Sesarma dehaani)提供阴凉潮湿的环境, 有效地缓解了失水胁迫, 从而对土著食物网中的群落结构和物种组成产生影响。 | III | Wang et al., 2008 |
| 盐沼 | 西班牙西北部加利西亚 | 狐米草 S. patens | 入侵植物不能被土著草食性昆虫取食, 但是可以通过腐食者进入到土著碎屑食物网, 改变了食物网有机质的循环路径。 | II | Page et al., 2010 |
| 盐沼 | 美国新泽西 | 芦苇 Phragmites australis | 芦苇入侵使得原先取食土著植物互花米草的节肢动物改为取食碎屑和底栖微藻。 | II | Gratton & Denno, 2006 |
| 盐沼 | 美国提瓦那河口 | 多枝柽柳 Tamarisk ramosissima | 多枝柽柳碎屑能够被土壤表层大型无脊椎动物取食利用按照原路径进入土著食物网中(途径I); 而土壤下层无脊椎动物不能够利用入侵植物碎屑导致其丰度降低(途径II)。 | I、II | Whitcraft et al., 2008 |
| 盐沼 | 美国旧金山湾 | 互花米草 S. alterni?ora | 入侵植物互花米草能够被光蝉(Prokelisia marginata)等土著广食性草食者取食, 进入到土著食物网中。 | I | Daehler & Strong, 1994 |
| 盐沼 | 美国旧金山湾 | 穗状狐尾藻 Myriophyllum spicatum | 入侵植物复杂的形态结构为一些底栖无脊椎动物或者幼鱼提供了庇护场所, 能够阻止更高级消费者对这些动物的取食。 | III | Chaplin & Valentine, 2009 |
| 盐沼 | 美国旧金山湾 | 互花米草 S. alterni?ora | 入侵植物产生的大量碎屑没有被土著底栖动物取食利用(途径II); 同时入侵植物作为生态系统工程师能够改变植物结构、沉积物性质、光照、水文等环境特点, 对土著生物群落与食物网产生影响(途径III: 植物形态结构、环境理化性质)。 | II、III | Brusati & Grosholz, 2009 |
| 盐沼 | 美国旧金山湾 | 互花米草 S. alterni?ora | 入侵植物不能被土著消费者双壳类白缨蛤(Macoma petaluma)取食利用,导致双壳类消费者丰度与丰富度降低(途径II); 另外互花米草提高盐沼的海拔高度, 降低水流流速与光照渗透强度, 通过生境的变化影响底栖动物食物网(途径III: 环境理化性质)。 | II、III | Brusati & Grosholz, 2007 |
| 盐沼 | 美国东部 | 芦苇 P. australis | 来自芦苇的初级生产力能够进入到土著食物网中, 为河口游泳动物提供食物。 | I | Weis & Weis, 2003 |
| 盐沼 | 法国西北部 | Elymus athericus | 披碱草属植物E. athericus的碎屑分解较慢, 改变了微生境条件, 为蜘蛛提供了适合结网的生境和庇护所。 | III | Petillon et al., 2005 |
| 溪流 | 澳大利亚昆士兰 | 巴拉草 Brachiaria mutica | 入侵植物巴拉草不能被蜉蝣目动物Baetid bymphs、蚋科(Simuliidae)、米虾属(Caridina)、沼虾属(Macrobrachium)等土著水生动物取食, 不能融入到土著水生食物网中。 | II | Bunn et al., 1997 |
| 溪流 | 爱尔兰西部 | 彭土杜鹃 Rhododendron ponticum | 入侵植物彭土杜鹃的可利用性较差, 同时其产生的大量碎屑抑制了藻类的生长, 导致无脊椎动物丰度的降低, 食物来源类型也从基于藻类转向基于颗粒有机质为主, 造成食物网群落结构以及流通路径的改变。 | II | Hladyz et al., 2011 |
| 森林 | 英国雅芳希斯 国家公园 | 北美白珠树 Gaultheria shallon 寻石楠 Calluna vulgaris | 入侵植物能够通过广食性草食者进入到土著食物网(途径I); 而专食性消费者不能适应入侵植物而选择离开入侵区域(途径II)。 | I、II | Carvalheiro et al., 2010 |
| 森林 | 美国芝加哥 Morton植物园 | 金银忍冬 Lonicera maackii 药鼠李 Rhamnus cathartica | 入侵植物的枝条结构适合鸟类筑巢, 也适合捕食者穿行, 使知更鸟鸟巢更容易遭受捕食者的破坏, 繁殖成功率降低, 因此入侵植物通过形态结构的变化对食物网产生了非营养作用。 | III | Schmidt & Whelan, 1999 |
| 森林 | 美国密西根 | 小蔓长春花 Vinca minor | 入侵植物改变了森林地被物的物理特性, 茂密的入侵植物限制了蜘蛛的活动能力, 造成蜘蛛丰度、均匀度和多样性的降低, 进而通过捕食作用影响昆虫食物网结构。 | III | Bultman & DeWitt, 2008 |
| 森林 | 美国 | 多种外来树种 | 鳞翅目昆虫对入侵植物没有形成生理和行为上的适应, 不能够利用外来植物作为营养来源和宿主, 导致生物量和多样性下降, 进而导致更高营养级的食虫动物种类和数量下降。 | II | Tallamy & Shropshire, 2009 |
| 森林 | 美国东部 | 葱芥 Alliaria petiolata | 入侵植物葱芥能够向土壤中渗出植物毒素芥子油苷(glucosinolate), 抑制丛枝菌根真菌的生长, 从而影响到土著植物的生长繁殖。 | III | Stinson et al., 2006 |
| 森林 | 南非 | 金合欢属伞树 Acacia saligna | 一方面入侵植物种子能够被啮齿类纹鼠(Rhabdomys pumilio)大量取食, 从而进入到土著食物网中(途径I); 另一方面入侵植物对生境的改变不适合纹鼠挖洞筑巢, 使纹鼠种群密度降低, 导致食物网中物种组成的改变(途径III: 环境理化性质)。 | I、III | Holmes, 1990 |
| 湖泊 | 美国东北部 | 穗状狐尾藻 Myriophyllum spicatum | 入侵植物能够被鞘翅目动物Euhrychiopsis lecontei取食, 根茎生物量降低一半, 土著消费者通过下行效应抑制了入侵植物的扩散。 | I | Creed & Sheldon, 1995; Sheldon & Creed, 1995 |
| 湖泊 | 美国密歇根 | 穗状狐尾藻 M. spicatum | 穗状狐尾藻为多裂形植物, 能够为附生在植物表面的大型无脊椎动物提供更多的觅食场所以及更多的遮蔽面积, 因而在多裂型植物周围有更多的无脊椎动物密度和生物量。 | III | Cheruvelil et al., 2002 |
| 湖泊 | 新西兰, 瓦那卡湖Wanaka | 卷叶蜈蚣草 Lagarosiphon major 伊乐藻 Elodea canadensis | 入侵区无脊椎动物群落密度和多样性比非入侵区域大, 优势物种发生改变, 食物网碳流通路径从基于沉积颗粒有机质为主转向基于水生植物为主。 | II | Kelly & Hawes, 2005 |
| 红树林 | 美国夏威夷、波多黎各岛 | 美洲红树 Rhizophora mangle | 美洲红树具有较高的单宁含量, 能够破坏许多生物正常的消化吸收功能, 较高的C:N比和木质素含量使得其叶片的营养价值较低。杂食者蓝蟹(Callinectes sapidus)、悬浮取食者贻贝(Mytilus sp.)、表层取食者滨螺(Littoraria sp.)等消费者无法适应这种植物碎屑, 其入侵区消费者种类与数量降低。 | II | Demopoulos et al., 2007 |
| 河流泛滥区 | 新西兰东南部 | 大叶醉鱼草 Buddleja davidii | 入侵植物使得根系周围土壤C、土壤细菌生物量增加, 土壤微生物群落结构发生改变, 土壤食微线虫和肉食性线虫丰度增加, 通过营养关系改变了食物网结构。 | II | Peltzer et al., 2009 |
| 海岸沙滩 | 西班牙西北部Galician海岸 | 海黍子马尾藻 Sargassum muticum | 入侵植物在冬季能够被底栖无脊椎动物取食, 通过途径I进入到土著食物网中; 而在春季由于土著藻类的大量生长繁殖, 入侵植物不能被取食, 导致底栖无脊椎动物的减少。入侵植物在春季通过途径II改变食物网。 | I、II | Rossi et al., 2010 |
| 海岸草地 | 德国波罗的海群岛 | 曲柄藓属植物Campylopus intro?exus | 曲柄藓属植物C. intro?exus入侵区杂草较少, 限制了草食性昆虫的生长, 导致入侵区步甲科物种丰度的降低(途径II); 另外C. intro?exu的形态结构不适合蜘蛛结网, 还通过改变周围生境中的温度和湿度降低了步甲科物种与蜘蛛的丰富度, 通过非营养作用改变食物网的物种组成(途径III: 形态结构与理化性质)。 | II、III | Schirmel et al., 2011 |
| 海岸草地 | 美国加利福尼亚沿岸 | 马德雀麦 Bromus madritensis 裂稃燕麦 Avena barbata | 入侵植物碎屑增加土壤湿度, 促进本地植物的生长, 增加灌丛草食动物叶蝉和捕食者蜘蛛的丰度, 从而对食物网产生影响。 | III | Wolkovich, 2010 |
| 海岸草地 | 美国加利福尼亚沿岸 | 莫邪菊 Carpobrotus edulis | 入侵植物的果实能够被土著小型哺乳动物加利福尼亚兔(Lepus californicus)以及骡鹿(Odocoileus hemionus)取食, 种子能够通过动物粪便传播扩散。 | I | Vila & D'Antonio, 1998 |
| 海岸草地 | 美国加利福尼亚沿岸 | 二穗短柄草、马德雀麦等Brachypodium distachyon B. madritensis etc. | 植物入侵产生的大量凋落物使得土著节肢动物蚂蚁和螨虫等物种的丰度降低, 通过营养级联影响到入侵生态系统食物网结构。 | II | Wolkovich et al., 2009 |
| 岛屿 | 新西兰东南部 | 花葵 Lavatera arborea | 入侵植物成为岛屿陆地生态系统的重要组成部分, 从传粉者到顶级捕食者多个营养级物种都有来自花葵的同位素标记, 入侵植物已经完全融入到土著食物网中。 | I | Hawke & Clark, 2010 |
| 岛屿 | 大西洋北部Azores群岛 | 日本柳杉、波叶海桐等Cryptomeria japonica, Pittosporum undulatum etc. | 植物入侵导致取食土著植物的大型昆虫消失, 外来小型昆虫进入, 形成新的食物网流通路径。 | II | Heleno et al., 2009 |
| 池塘 | 控制实验 | 多种水生植物 | 外来入侵植物能够被土著广食性草食者取食, 通过营养作用进入到土著食物网中。 | I | Parker & Hay, 2005 |
| 池塘 | 控制实验 | 黑藻 Hydrilla verticillata | 植物入侵导致入侵区域的空间异质性降低, 改变了大型无脊椎动物的类群, 造成食物网中物种组成的变化。 | III | Theel et al., 2008 |
| 池塘 | 美国西北部 | 千屈菜和鹬草 Lythrum salicaria and Phalaris arundinacea | 土著草食者飞蛾等不能够取食入侵植物而选择离开入侵区域, 造成入侵区食物网中物种组成的变化。 | II | Schooler et al., 2009 |
| 池塘 | 北美 | 千屈菜 L. salicaria | 入侵植物叶片由于具有较高的单宁含量, 可利用性较低, 不能被土著消费者美洲蟾蜍(Bufo americanus)的蝌蚪取食, 导致入侵区美洲蟾蜍的成活率显著降低。 | II | Brown et al., 2006 |
| 草地 | 美国蒙大拿州 | 斑点矢车菊 Centaurea maculosa | 通过引入矢车菊的天敌果蝇来控制植物入侵, 但是果蝇数量的迅速增加导致其捕食者鹿鼠增加, 鹿鼠又取食土著植物种子,从而形成新的食物网结构。 | II | Ortega et al., 2004 |
| 草地 | 控制实验 | 斑点矢车菊 C. maculosa | 入侵植物较高的植物密度以及复杂的形态结构提高了卷叶蛛(Dictyna)的密度与结网面积, 降低了卷叶蛛的猎物实蝇科昆虫Urophora affinis的密度, 入侵植物通过形态特征的变化以及对环境条件的改变影响昆虫食物网。 | III | Pearson, 2010 |
| 草地 | 北美西部 | 斑点矢车菊 C. maculosa | 矢车菊的形态结构方便蜘蛛结网, 入侵区蜘蛛网大小与捕获量分别是未入侵区的2.9-4倍和2-2.3倍, 入侵植物形态结构通过捕食者产生下行效应, 影响昆虫食物网的物种组成。 | III | Pearson, 2009 |
| 草地 | 澳大利亚北部 | 大含羞草 Mimosa pigra | 在入侵植物含羞草上发现有50科,101属,共114种植食性昆虫, 入侵植物既能够被土著草食者取食进入到土著食物网中(途径I), 也能够被外来草食者利用形成新的食物网结构(途径II)。 | I、II | Wilson et al., 1990 |
| 草地 | 荷兰 | 菊科和十字花科草本植物 | 植物入侵区域虽然根系生物量较多, 但是却无法被土著消费者取食, 导致入侵植物根系周围线虫多样性和生物量显著降低。 | II | Morrien et al., 2012 |
| 草地 | 英国西部 | 凤仙花 Impatiens glandulifera | 外来植物凤仙花具有较高质量的花蜜, 能够吸引较多的传粉者, 入侵区传粉昆虫丰富度与丰度显著提高, 同时也促进了入侵植物的成功授粉。 | III | Lopezaraiza-Mikel et al., 2007 |
| 草地 | 美国犹他州东南部 | 旱雀麦 Bromus tectorum | 线虫、微型节肢动物等土壤动物不能取食入侵植物, 而选择离开入侵区域, 导致土壤食物网结构与物种组成发生变化。 | II | Belnap et al., 2005 |
| 草地 | 波兰南部 | 一枝黄花 Solidago spp. | 入侵植物造成蝴蝶、昆虫等无脊椎动物丰度降低, 引起以这些动物为食物的鸟类种类和数量的减少(途径II)。另外, 入侵植物的形态结构不适合鸟类筑巢并且入侵区缺少鸟类筑巢所需的材料, 导致鸟类繁殖成功率的下降(途径III: 植物形态结构)。 | II、III | Skorka et al., 2010 |
| 草地 | 美国落基山国家公园 | 乳浆大戟和丝路蓟 Euphorbia esula and Cirsium arvense | 入侵植物不能被土著土壤动物取食, 土壤动物由于缺少食物资源而离开入侵区域(途径II);同时植物入侵使得裸地斑块增加, 改变了土壤有机质含量、pH值以及多种微量元素含量, 导致土著土壤食物网中蜱螨目等小型节肢动物丰度在入侵区显著降低(途径III: 理化性质)。 | II、III | Pritekel et al., 2006 |
| 草地 | 控制实验 | 多种外来植物 | 外来入侵植物降低土著昆虫生物量, 通过营养级联作用影响较高营养级的食虫动物丰度, 严重扰乱了入侵区陆地食物网结构。 | II | Tallamy et al., 2010 |
| 水生系统 | 控制实验 | 凤眼蓝、芋等多种水生植物 Eichhornia crassipes, Colocasia esculenta | 土著广食性草食者苹螺(Pomacea sp.)以及Procambarus spiculifer等对外来入侵植物的取食是土著植物的2-17倍, 入侵植物能够被土著系统中的广食性草食者取食利用, 融入土著食物网中, 而广食性草食者的取食抑制了入侵植物的扩散。 | I | Morrison & Hay, 2011 |
| 水生系统 | 综述 | 大型水生植物 | 大型水生植物的入侵导致生境复杂性增加、水体缺氧, 释放化学物质产生异株相克化学作用, 改变土著动植物群落结构及物种组成(途径III: 植物形态特征和环境理化性质); 另一方面能够降低食物质量, 导致土著鱼类和大型无脊椎动物丰度的降低(途径II)。 | II、III | Schultz & Dibble, 2012 |
| 多种系统 | 捷克 | 13种入侵植物 | 入侵植物的株高、覆盖度及根系的密集度等特点改变了入侵区群落结构物种组成与物种丰富度。 | III | Hejda et al., 2009 |
| 多种系统 | 综述 | 100多种入侵植物 | 外来植物对土著草食者还没有形成防御策略, 更容易受到土著草食者的取食, 进入土著食物网。 | I | Parker et al., 2006 |
| 12 | Carvalheiro LG, Buckley YM, Memmott J (2010) Diet breadth influences how the impact of invasive plants is propagated through food webs.Ecology, 91, 1063-1074. |
| 13 | Cebrian E, Ballesteros E, Linares C, Tomas F (2011) Do native herbivores provide resistance to Mediterranean marine bioinvasions? A seaweed example.Biological Invasions, 13, 1397-1408. |
| 14 | Chen HL, Li B, Hu JB, Chen JK, Wu JH (2007) Effects of Spartina alterniflora invasion on benthic nematode communities in the Yangtze Estuary.Marine Ecology Progress Series, 336, 99-110. |
| 15 | Cheruvelil KS, Soranno PA, Madsen JD, Roberson MJ (2002) Plant architecture and epiphytic macroinvertebrate communities: the role of an exotic dissected macrophyte.Journal of the North American Benthological Society, 21, 261-277. |
| 16 | Chittka L, Schurkens S (2001) Successful invasion of a floral market-an exotic Asian plant has moved in on Europe’s river-banks by bribing pollinators.Nature, 411, 653-653. |
| 17 | Chown SL, Huiskes AHL, Gremmen NJM, Lee JE, Terauds A, Crosbie K, Frenot Y, Hughes KA, Imura S, Kiefer K, Lebouvier M, Raymond B, Tsujimoto M, Ware C, Van de Vijver B, Bergstrom DM (2012) Continent-wide risk assessment for the establishment of nonindigenous species in Antarctica.Proceedings of the National Academy of Sciences,USA, 109, 4938-4943. |
| 18 | Christian JM, Wilson SD (1999) Long-term ecosystem impacts of an introduced grass in the northern Great Plains.Ecology, 80, 2397-2407. |
| 19 | Cohen JS, Ng S, Blossey B (2012) Quantity counts: amount of litter determines tadpole performance in experimental microcosms.Journal of Herpetology, 46, 85-90. |
| 20 | Creed RP, Sheldon SP (1995) Weevils and watermilfoil: did a north-American herbivore cause the decline of an exotic plant ? Ecological Applications, 5, 1113-1121. |
| 21 | Crooks JA (2002) Characterizing ecosystem-level consequences of biological invasions: the role of ecosystem engineers.Oikos, 97, 153-166. |
| 22 | Daehler CC, Strong DR (1994) Variable reproductive output among clones of Spartina alterniflora (Poaceae) invading San Francisco Bay, California: the influence of herbivory, pollination, and establishment site.American Journal of Botany, 81, 307-313. |
| 23 | Dietzsch AC, Stanley DA, Stout JC (2011) Relative abundance of an invasive alien plant affects native pollination processes.Oecologia, 167, 469-479. |
| 24 | Dunne JA, Williams RJ, Martinez ND (2002) Food-web structure and network theory: The role of connectance and size.Proceedings of the National Academy of Sciences, USA, 99, 12917-12922. |
| 25 | Ehrenfeld JG (2010) Ecosystem consequences of biological invasions.Annual Review of Ecology, Evolution, and Systematics, 41, 59-80. |
| 26 | Elton CS (1927) Animal Ecology. Republished 2001. University of Chicago Press, Chicago. |
| 27 | Fridley JD (2012) Extended leaf phenology and the autumn niche in deciduous forest invasions.Nature, 485, 359-362. |
| 28 | Gilbert AJ (2009) Connectance indicates the robustness of food webs when subjected to species loss.Ecological Indicators, 9, 72-80. |
| 29 | Gratton C, Denno RF (2006) Arthropod food web restoration following removal of an invasive wetland plant.Ecological Applications, 16, 622-631. |
| 30 | Gross H, Goeger DE, Hills P, Mooberry SL, Ballantine DL, Murray TF, Valeriote FA, Gerwick WH (2006) Lophocladines, bioactive alkaloids from the red alga Lophocladia sp.Journal of Natural Products, 69, 640-644. |
| 31 | Harvey JA, Bukovinszky T, van der Putten WH (2010) Interactions between invasive plants and insect herbivores: a plea for a multitrophic perspective.Biological Conservation, 143, 2251-2259. |
| 32 | Heleno RH, Ceia RS, Ramos JA, Memmott J (2009) Effects of alien plants on insect abundance and biomass: a food-web approach.Conservation Biology, 23, 410-419. |
| 33 | Holmes PM (1990) Dispersal and predation in alien Acacia.Oecologia, 83, 288-290. |
| 34 | Kaiser-Bunbury CN, Valentin T, Mougal J, Matatiken D, Ghazoul J (2011) The tolerance of island plant-pollinator networks to alien plants.Journal of Ecology, 99, 202-213. |
| 35 | Kardol P, Wardle DA (2010) How understanding aboveground-belowground linkages can assist restoration ecology.Trends in Ecology and Evolution, 25, 670-679. |
| 36 | Kelly DJ, Hawes I (2005) Effects of invasive macrophytes on littoral-zone productivity and foodweb dynamics in a New Zealand high-country lake.Journal of the North American Benthological Society, 24, 300-320. |
| 37 | Kimball ME, Able KW, Grothues TM (2010) Evaluation of long-term response of intertidal creek nekton to Phragmites australis (Common Reed) removal in oligohaline Delaware Bay salt marshes.Restoration Ecology, 18, 772-779. |
| 38 | Klironomos JN (2002) Feedback with soil biota contributes to plant rarity and invasiveness in communities.Nature, 417, 67-70. |
| 39 | Kovalenko KE, Dibble ED (2011) Effects of invasive macrophyte on trophic diversity and position of secondary consumers.Hydrobiologia, 663, 167-173. |
| 40 | Lambrinos JG (2004) How interactions between ecology and evolution influence contemporary invasion dynamics.Ecology, 85, 2061-2070. |
| 41 | Levin LA, Neira C, Grosholz ED (2006) Invasive cordgrass modifies wetland trophic function.Ecology, 87, 419-432. |
| 42 | Levine JM (2000) Species diversity and biological invasions: relating local process to community pattern.Science, 288, 852-854. |
| 43 | Liang X, Zheng H, He CQ, Xu QY, Zhan YW, Lei YR, Du W, Yang JN (2012) Allelopathic effects of invasive Spartina alterniflora root exudates in soil on the offspring (seeds) of Scirpus mariqueter.Allelopathy Journal, 29, 251-262. |
| 44 | Liao CZ, Luo YQ, Fang CM, Chen JK, Li B (2008a) Litter pool sizes, decomposition, and nitrogen dynamics in Spartina alterniflora-invaded and native coastal marshlands of the Yangtze Estuary.Oecologia, 156, 589-600. |
| 45 | Liao CZ, Peng RH, Luo YQ, Zhou XH, Wu XW, Fang CM, Chen JK, Li B (2008b) Altered ecosystem carbon and nitrogen cycles by plant invasion: a meta-analysis.New Phytologist, 177, 706-714. |
| 1 | Allesina S, Tang S (2012) Stability criteria for complex ecosystems.Nature, 483, 205-208. |
| 2 | Baiser B, Russell GJ, Lockwood JL (2010) Connectance determines invasion success via trophic interactions in model food webs.Oikos, 119, 1970-1976. |
| 46 | Liao CZ, Luo YQ, Jiang LF, Zhou XH, Wu XW, Fang CM, Chen JK, Li B (2007) Invasion of Spartina alterniflora enhanced ecosystem carbon and nitrogen stocks in the Yangtze Estuary, China.Ecosystems, 10, 1351-1361. |
| 47 | Lonsdale WM (1999) Global patterns of plant invasions and the concept of invasibility.Ecology, 50, 1522-1536. |
| 3 | Bartuszevige AM, Gorchov DL, Raab L (2006) The relative importance of landscape and community features in the invasion of an exotic shrub in a fragmented landscape.Ecography, 29, 213-222. |
| 4 | Benoit LK, Askins RA (1999) Impact of the spread of Phragmites on the distribution of birds in Connecticut tidal marshes.Wetlands, 19, 194-208. |
| 5 | Bersier LF, Banašek-Richter C, Cattin MF (2002) Quantitative descriptors of food-web matrices.Ecology, 83, 2394-2407. |
| 6 | Bradford MA, Strickland MS, DeVore JL, Maerz JC (2012) Root carbon flow from an invasive plant to belowground foodwebs.Plant and Soil, 359, 233-244. |
| 7 | Briggs CJ, Borer ET (2005) Why short-term experiments may not allow long-term predictions about intraguild predation.Ecological Applications, 15, 1111-1117. |
| 48 | Lopezaraiza-Mikel ME, Hayes RB, Whalley MR, Memmott J (2007) The impact of an alien plant on a native plant-pollinator network: an experimental approach.Ecology Letters, 10, 539-550. |
| 49 | Maron JL, Vilà M (2001) When do herbivores affect plant invasion? Evidence for the natural enemies and biotic resistance hypotheses.Oikos, 95, 361-373. |
| 50 | Moragues E, Traveset A (2005) Effect of Carpobrotus spp. on the pollination success of native plant species of the Balearic Islands.Biological Conservation, 122, 611-619. |
| 8 | Bukovinszky T, van Veen FJF, Jongema Y, Dicke M (2008) Direct and indirect effects of resource quality on food web structure.Science, 319, 804-807. |
| 9 | Bultman TL, DeWitt DJ (2008) Effect of an invasive ground cover plant on the abundance and diversity of a forest floor spider assemblage.Biological Invasions, 10, 749-756. |
| 51 | Morin PJ (2011) Food webs. In: Community Ecology, 2nd edn. (ed. Morin PJ) pp. 136-165. Blackwell Publishing, Chichester, UK. |
| 52 | Morrison WE, Hay ME (2011) Herbivore preference for native vs. exotic plants: generalist herbivores from multiple continents prefer exotic plants that are evolutionarily naive.PLoS ONE, 6, e17227. |
| 10 | Cappuccino N, Arnason JT (2006) Novel chemistry of invasive exotic plants.Biology Letters, 2, 189-193. |
| 11 | Carroll SP, Loye JE, Dingle H, Mathieson M, Famula TR, Zalucki MP (2005) And the beak shall inherit-evolution in response to invasion.Ecology Letters, 8, 944-951. |
| 53 | Naeem S, Knops JMH, Tilman D, Howe KM, Kennedy T, Gale S (2000) Plant diversity increases resistance to invasion in the absence of covarying extrinsic factors.Oikos, 91, 97-108. |
| 54 | Ortega YK, Pearson DE, McKelvey KS (2004) Effects of biological control agents and exotic plant invasion on deer mouse populations.Ecological Applications, 14, 241-253. |
| 55 | Page HM, Lastra M, Rodil IF, Briones MJI, Garrido J (2010) Effects of non-native Spartina patens on plant and sediment organic matter carbon incorporation into the local invertebrate community.Biological Invasions, 12, 3825-3838. |
| 56 | Palmer MA, Ambrose RF, Poff NL (1997) Ecological theory and community restoration ecology.Restoration Ecology, 5, 291-300. |
| 57 | Parker JD, Burkepile DE, Hay ME (2006) Opposing effects of native and exotic herbivores on plant invasions.Science, 311, 1459-1461. |
| 58 | Pearson DE (2009) Invasive plant architecture alters trophic interactions by changing predator abundance and behavior.Oecologia, 159, 549-558. |
| 59 | Pearson DE (2010) Trait- and density-mediated indirect interactions initiated by an exotic invasive plant autogenic ecosystem engineer.The American Naturalist, 176, 394-403. |
| 60 | Pejchar L, Mooney HA (2009) Invasive species, ecosystem services and human well-being.Trends in Ecology and Evolution, 24, 497-504. |
| 61 | Peltzer DA, Bellingham PJ, Kurokawa H, Walker LR, Wardle DA, Yeates GW (2009) Punching above their weight: low-biomass non-native plant species alter soil properties during primary succession.Oikos, 118, 1001-1014. |
| 62 | Pimm SL (1979) The structure of food webs.Theoretical Population Biology, 16, 144-158. |
| 63 | Pimm SL, Lawton JH, Cohen JE (1991) Food web patterns and their consequences.Nature, 350, 669-674. |
| 64 | Pritekel C, Whittemore-Olson A, Snow N, Moore JC (2006) Impacts from invasive plant species and their control on the plant community and belowground ecosystem at Rocky Mountain National Park, USA.Applied Soil Ecology, 32, 132-141. |
| 65 | Richards CL, Bossdorf O, Muth NZ, Gurevitch J, Pigliucci M (2006) Jack of all trades, master of some? On the role of phenotypic plasticity in plant invasions.Ecology Letters, 9, 981-993. |
| 66 | Rinella MJ, Mangold JM, Espeland EK, Sheley RL, Jacobs JS (2012) Long-term population dynamics of seeded plants in invaded grasslands.Ecological Applications, 22, 1320-1329. |
| 67 | Rooney N, McCann KS (2012) Integrating food web diversity, structure and stability.Trends in Ecology and Evolution, 27, 40-46. |
| 68 | Rossi F, Olabarria C, Incera M, Garrido J (2010) The trophic significance of the invasive seaweed Sargassum muticum in sandy beaches.Journal of Sea Research, 63, 52-61. |
| 69 | Schmidt KA, Whelan CJ (1999) Effects of exotic Lonicera and Rhamnus on songbird nest predation.Conservation Biology, 13, 1502-1506. |
| 70 | Stinson KA, Campbell SA, Powell JR, Wolfe BE, Callaway RM, Thelen GC, Hallett SG, Prati D, Klironomos JN (2006) Invasive plant suppresses the growth of native tree seedlings by disrupting belowground mutualisms.PLoS Biology, 4, e140. |
| 71 | Sugiura S (2010) Species interactions-area relationships: biological invasions and network structure in relation to island area.Proceedings of the Royal Society B: Biological Sciences, 277, 1807-1815. |
| 72 | Sultan SE (2001) Phenotypic plasticity for fitness components in Polygonum species of contrasting ecological breadth.Ecology, 82, 328-343. |
| 73 | Wang JQ, Zhang XQ, Nie M, Fu CZ, Chen JK, Li B (2008) Exotic Spartina alterniflora provides compatible habitats for native estuarine crab Sesarma dehaani in the Yangtze River Estuary.Ecological Engineering, 34, 57-64. |
| 74 | Williams NM, Minckley RL, Silveira FA (2001) Variation in native bee faunas and its implications for detecting community changes.Conservation Ecology, 5, 7. |
| 75 | Wilson CG, Flanagan GJ, Gillett JD (1990) The phytophagous insect fauna of the introduced shrub Mimosa pigra in northern Australia and its relevance to biological-control.Environmental Entomology, 19, 776-784. |
| 76 | Wolkovich EM (2010) Nonnative grass litter enhances grazing arthropod assemblages by increasing native shrub growth.Ecology, 91, 756-766. |
| 77 | Woodcock BA, Bullock JM, Mortimer SR, Pywell RF (2012) Limiting factors in the restoration of UK grassland beetle assemblages.Biological Conservation, 146, 136-143. |
| 78 | Wu YT, Wang CH, Zhang XD, Zhao B, Jiang LF, Chen JK, Li B (2009) Effects of saltmarsh invasion by Spartina alterniflora on arthropod community structure and diets.Biological Invasions, 11, 635-649. |
| 79 | Xu CY, Griffin KL, Schuster WSF (2007) Leaf phenology and seasonal variation of photosynthesis of invasive Berberis thunbergii (Japanese barberry) and two co-occurring native understory shrubs in a northeastern United States deciduous forest.Oecologia, 154, 11-21. |
| 80 | Zedler JB (2000) Progress in wetland restoration ecology.Trends in Ecology and Evolution, 15, 402-407. |
| [1] | Chen Dingsong, Liu Zikai, He Ziyang, Chen Weidong. Advances in tardigrade diversity, distribution characteristics and ecological functions [J]. Biodiv Sci, 2025, 33(2): 24406-. |
| [2] | Xingda Yao, Kaiyuan Zhu, Yihan Lan, Shuxia Zhan, Tana Wuyun, Jian Li, Wenting Xu. Effects of alien invasive plants on spontaneous plant communities in Hangzhou Xixi National Wetland Park [J]. Biodiv Sci, 2025, 33(12): 25212-. |
| [3] | Rongjiang Zhao, Jihua Wu, Weiming He, Caiyun Zhao, Bo Zhou, Bo Li, Qiang Yang. Soil biodiversity and exotic plant invasions: Progress and perspectives [J]. Biodiv Sci, 2024, 32(11): 24243-. |
| [4] | Yongjie Niu, Quanhui Ma, Yu Zhu, Hairong Liu, Jiale Lü, Yuanchun Zou, Ming Jiang. Research progress on the impact of nitrogen deposition on grassland insect diversity [J]. Biodiv Sci, 2023, 31(9): 23130-. |
| [5] | Yong Li, Sanqing Li, Huan Wang. A dataset of wild vascular plants and their distributions in Tianjin, China [J]. Biodiv Sci, 2023, 31(9): 23128-. |
| [6] | Yu Xiao, Yuran Li, Hexiang Duan, Zhengtao Ren, Shengbi Feng, Zhicheng Jiang, Jiahua Li, Pin Zhang, Jinming Hu, Yupeng Geng. Invasion status and control measures for alien plants within the Gaoligong Mountains [J]. Biodiv Sci, 2023, 31(5): 23011-. |
| [7] | Yaochu Sun, Yuanfei Pan, Mu Liu, Xiaoyun Pan. The specialist-to-generalist ratio affects growth and defense strategy of invasive plant Alternanthera philoxeroides [J]. Biodiv Sci, 2023, 31(4): 22632-. |
| [8] | Xia Cui, Quanru Liu, Chaoran Wu, Yufei He, Jinshuang Ma. The alien invasive plants in Beijing-Tianjin-Hebei [J]. Biodiv Sci, 2022, 30(8): 21497-. |
| [9] | Qinwen Lin, Cui Xiao, Jinshuang Ma. A dataset on catalogue of alien plants in China [J]. Biodiv Sci, 2022, 30(5): 22127-. |
| [10] | Chaodan Guo, Caiyun Zhao, Feifei Li, Junsheng Li. Comparative study of invasive and native herbs in natural forests and plantation forests: With Nonggang National Nature Reserve as an example [J]. Biodiv Sci, 2022, 30(4): 21356-. |
| [11] | Haifeng Yao, Saichao Zhang, Huayuan Shangguan, Zhipeng Li, Xin Sun. Effects of urbanization on soil fauna community structure and diversity [J]. Biodiv Sci, 2022, 30(12): 22547-. |
| [12] | Baomin Yao, Qing Zeng, Limei Zhang. Research progress on the biodiversity and ecological function of soil protists [J]. Biodiv Sci, 2022, 30(12): 22353-. |
| [13] | Huiling Hu, Zhiyuan Yao, Shibin Gao, Bo Zhu. Nematode response to long-term fertilization in purple soil [J]. Biodiv Sci, 2022, 30(12): 22189-. |
| [14] | Ya Wang, Weiqian Wang, Qinke Wang, Xiaoxia Li, Yan Liu, Qiaoqiao Huang. Effects of soil nutrients on reproductive traits of invasive and native annual Asteraceae plants [J]. Biodiv Sci, 2021, 29(1): 1-9. |
| [15] | Xue Han, Jinquan Su, Nana Yao, Baoming Chen. Advances in root foraging behavior of exotic invasive plants [J]. Biodiv Sci, 2020, 28(6): 727-733. |
| Viewed | ||||||
|
Full text |
|
|||||
|
Abstract |
|
|||||
Copyright © 2026 Biodiversity Science
Editorial Office of Biodiversity Science, 20 Nanxincun, Xiangshan, Beijing 100093, China
Tel: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn