SLOSS争论研究进展: 分析方法、理论机制及保护实践
章洋, 王彦平
生物多样性
2025, 33 ( 7):
25081-.
DOI: 10.17520/biods.2025081
生物多样性保护领域中的SLOSS (single large or several small)争论概述了在总面积相等或资源有限的情况下, 应该优先保护一个大的保护区(SL)还是几个小的保护区(SS)来最大化保护物种多样性的问题。自20世纪70年代以来, 该问题一直备受生态学家和保护生物学家的关注, 其研究对于保护区设计以及生物多样性保护具有重要意义。随着研究方法的发展和交叉学科的兴起, SLOSS争论在分析方法和理论机制方面都取得了重要的阶段性进展。本文通过文献检索和归纳总结对SLOSS争论的研究现状进行了综述。首先, 我们回顾了SLOSS争论的起源及其对生物多样性保护的重要性。其次, 本文概括评估了SLOSS争论在实证研究和理论方面的分析方法, 包括零模型法、经典的物种累积曲线法以及由该方法所计算出来的饱和指数。随后, 本文还回顾了以往研究者提出的改进的饱和指数, 并介绍了我们所开发的优化后的饱和指数。此外, 本文探讨了几种影响SLOSS争论的理论机制, 如由灭绝率和定殖率的变化所主导的灭绝-定殖动态理论机制及其相应的检验方法。最后, 本文对该领域进行了总结和展望并为未来的发展方向提出了一些针对性建议, 包括采用多种保护策略的重要性以及在保护区规划中经济和生态因素的结合的必要性。本文整合了SLOSS争论中常用的分析方法的适用场景与局限性, 比较了不同方法在划分保护优先级时的优势和争议点, 可为研究者在方法选择及保护规划权衡方面提供参考。
View image in article
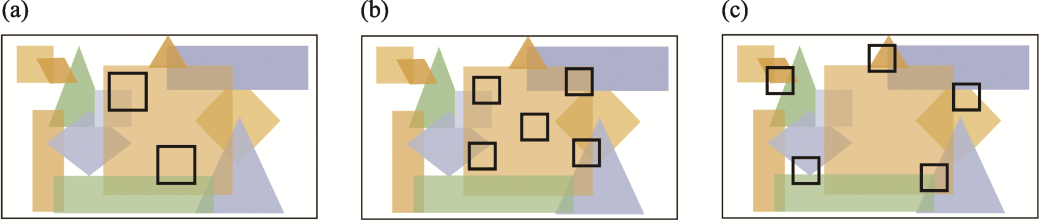
图5
当物种呈聚集分布时, 少量大斑块相交的物种数量(a)少于几个小斑块(b)和(c)。不同的颜色及相应的形状代表生境丧失前单个连续栖息地中的不同物种。黑色方框代表因生境丧失而形成的斑块。当少数大斑块和几个小斑块的斑块边缘之间的最大距离相等或类似时, 几个小斑块集能更均匀地覆盖在整个景观中, 因此相交的物种数更多(Fahrig et al, 2022)。(a)中2个大斑块与3个物种相交, 然而(b)中5个小斑块与6个物种相交。在斑块之间相距较远的情况下, 这种效应还会放大: (c)中5个小斑块相交了11个物种(该图改自Fahrig等, 2022)。
正文中引用本图/表的段落
事实上, 大多数物种往往呈现聚集分布的趋势(Seidler & Plotkin, 2006; Morlon et al, 2008)。这种趋势由多种因素引起, 包括抵御入侵种或共同寻找食物资源时的集群以减少死亡率, 比如冬季哺乳动物个体之间的社会联系来定位觅食区域或对抗(Peignier et al, 2019), 雀形目中雌鸟的筑巢位置更靠近边界(Ramsay et al, 1999)等, 这些统称为同种吸引(Vité & Francke, 1976; Schuck-Paim & Alonso, 2001)和恋巢性(Weatherhead & Forbes, 1994)。然而, 动物倾向于聚集分布的主要驱动力或许来自它们对部分微生境(局部地形、小气候)的响应(详见Fahrig等(2022)的综述论文)。当区域因栖息地破碎化或土地清理等过程形成几个小斑块时, 与少数大斑块相比, 前者将与那些在斑块形成之前就已经存在的物种更广泛地相交(图5), 这增加了SS中的β多样性, 因此可以预测SS > SL。Saravia等(2025)的最新研究在运用多物种空间显式模型探讨SLOSS问题时就明确考了虑物种数量稳定后的栖息地破碎化, 这种情况被认为比所有物种最初以随机模式出现在破碎的栖息地中更现实(Rybicki et al, 2020)。正如立方体假说中所提到的那样, 物种在破碎化之前的空间分布格局越异质性, 这种类比就越重要(Saravia et al, 2025)。
灭绝和定殖对SLOSS模式的影响在早期就已经得到了广泛的认识.以往的研究发现, 当灭绝率的变化主导了灭绝-定殖之间的权衡时应该发现SL > SS (McCarthy et al, 2005; Jagers & Harding, 2009).这是可以预期的, 由于几个小斑块总的边缘更长, 导致小斑块中的物种迁入率更大.因此, 扩散能力强的物种容易从原生斑块迁出, 而基质的不利性导致了迁移过程中非常高的死亡率(Flather & Bevers, 2002; Martin & Fahrig, 2016), 这降低了物种定殖的成功率, 从而使得SS中的物种数小于SL.此外, 不同的物种具有不同的最小斑块面积需求, 对于那些体型较大的动物来说, 它们对最小面积的需求也更高.因此, 当小型斑块不能满足物种的最小面积需求或维持物种生存的最小栖息地数量远远不足时, 由于选择性灭绝, SS中的物种灭绝率将高于SL, 这也会导致SL > SS (McCarthy et al, 2006; Tj?rve, 2010). ... A general framework for the distance-decay of similarity in ecological communities 2 2008 ... 事实上, 大多数物种往往呈现聚集分布的趋势(Seidler & Plotkin, 2006; Morlon et al, 2008).这种趋势由多种因素引起, 包括抵御入侵种或共同寻找食物资源时的集群以减少死亡率, 比如冬季哺乳动物个体之间的社会联系来定位觅食区域或对抗(Peignier et al, 2019), 雀形目中雌鸟的筑巢位置更靠近边界(Ramsay et al, 1999)等, 这些统称为同种吸引(Vité & Francke, 1976; Schuck-Paim & Alonso, 2001)和恋巢性(Weatherhead & Forbes, 1994).然而, 动物倾向于聚集分布的主要驱动力或许来自它们对部分微生境(局部地形、小气候)的响应(详见Fahrig等(2022)的综述论文).当区域因栖息地破碎化或土地清理等过程形成几个小斑块时, 与少数大斑块相比, 前者将与那些在斑块形成之前就已经存在的物种更广泛地相交(图5), 这增加了SS中的β多样性, 因此可以预测SS > SL.Saravia等(2025)的最新研究在运用多物种空间显式模型探讨SLOSS问题时就明确考了虑物种数量稳定后的栖息地破碎化, 这种情况被认为比所有物种最初以随机模式出现在破碎的栖息地中更现实(Rybicki et al, 2020).正如立方体假说中所提到的那样, 物种在破碎化之前的空间分布格局越异质性, 这种类比就越重要(Saravia et al, 2025). ...
灭绝和定殖对SLOSS模式的影响在早期就已经得到了广泛的认识.以往的研究发现, 当灭绝率的变化主导了灭绝-定殖之间的权衡时应该发现SL > SS (McCarthy et al, 2005; Jagers & Harding, 2009).这是可以预期的, 由于几个小斑块总的边缘更长, 导致小斑块中的物种迁入率更大.因此, 扩散能力强的物种容易从原生斑块迁出, 而基质的不利性导致了迁移过程中非常高的死亡率(Flather & Bevers, 2002; Martin & Fahrig, 2016), 这降低了物种定殖的成功率, 从而使得SS中的物种数小于SL.此外, 不同的物种具有不同的最小斑块面积需求, 对于那些体型较大的动物来说, 它们对最小面积的需求也更高.因此, 当小型斑块不能满足物种的最小面积需求或维持物种生存的最小栖息地数量远远不足时, 由于选择性灭绝, SS中的物种灭绝率将高于SL, 这也会导致SL > SS (McCarthy et al, 2006; Tj?rve, 2010). ... Space use and social association in a gregarious ungulate: Testing the conspecific attraction and resource dispersion hypotheses 1 2019 ... 事实上, 大多数物种往往呈现聚集分布的趋势(Seidler & Plotkin, 2006; Morlon et al, 2008).这种趋势由多种因素引起, 包括抵御入侵种或共同寻找食物资源时的集群以减少死亡率, 比如冬季哺乳动物个体之间的社会联系来定位觅食区域或对抗(Peignier et al, 2019), 雀形目中雌鸟的筑巢位置更靠近边界(Ramsay et al, 1999)等, 这些统称为同种吸引(Vité & Francke, 1976; Schuck-Paim & Alonso, 2001)和恋巢性(Weatherhead & Forbes, 1994).然而, 动物倾向于聚集分布的主要驱动力或许来自它们对部分微生境(局部地形、小气候)的响应(详见Fahrig等(2022)的综述论文).当区域因栖息地破碎化或土地清理等过程形成几个小斑块时, 与少数大斑块相比, 前者将与那些在斑块形成之前就已经存在的物种更广泛地相交(图5), 这增加了SS中的β多样性, 因此可以预测SS > SL.Saravia等(2025)的最新研究在运用多物种空间显式模型探讨SLOSS问题时就明确考了虑物种数量稳定后的栖息地破碎化, 这种情况被认为比所有物种最初以随机模式出现在破碎的栖息地中更现实(Rybicki et al, 2020).正如立方体假说中所提到的那样, 物种在破碎化之前的空间分布格局越异质性, 这种类比就越重要(Saravia et al, 2025). ... Model assessments of the optimal design of nature reserves for maximizing species longevity 1 2000 ... Existing theoretical models and their corresponding target assessments
大多数关于SLOSS争论的理论模型往往只考虑如何使生态效益最大化, 常忽略了经济因素在SLOSS争论中的重要性(Groeneveld, 2005), 包括经济成本、财政支出等, 而经济模型则综合考虑了生态效益和经济效益, 从而能够在资源有限的情况下对其进行合理分配以达到预期目标. ... Why there are more species in several small patches versus few large patches: A multispecies modelling approach 2 2025 ... 事实上, 大多数物种往往呈现聚集分布的趋势(Seidler & Plotkin, 2006; Morlon et al, 2008).这种趋势由多种因素引起, 包括抵御入侵种或共同寻找食物资源时的集群以减少死亡率, 比如冬季哺乳动物个体之间的社会联系来定位觅食区域或对抗(Peignier et al, 2019), 雀形目中雌鸟的筑巢位置更靠近边界(Ramsay et al, 1999)等, 这些统称为同种吸引(Vité & Francke, 1976; Schuck-Paim & Alonso, 2001)和恋巢性(Weatherhead & Forbes, 1994).然而, 动物倾向于聚集分布的主要驱动力或许来自它们对部分微生境(局部地形、小气候)的响应(详见Fahrig等(2022)的综述论文).当区域因栖息地破碎化或土地清理等过程形成几个小斑块时, 与少数大斑块相比, 前者将与那些在斑块形成之前就已经存在的物种更广泛地相交(图5), 这增加了SS中的β多样性, 因此可以预测SS > SL.Saravia等(2025)的最新研究在运用多物种空间显式模型探讨SLOSS问题时就明确考了虑物种数量稳定后的栖息地破碎化, 这种情况被认为比所有物种最初以随机模式出现在破碎的栖息地中更现实(Rybicki et al, 2020).正如立方体假说中所提到的那样, 物种在破碎化之前的空间分布格局越异质性, 这种类比就越重要(Saravia et al, 2025). ...
大多数关于SLOSS争论的理论模型往往只考虑如何使生态效益最大化, 常忽略了经济因素在SLOSS争论中的重要性(Groeneveld, 2005), 包括经济成本、财政支出等, 而经济模型则综合考虑了生态效益和经济效益, 从而能够在资源有限的情况下对其进行合理分配以达到预期目标. ... Introducing the key patch approach for habitat networks with persistent populations: An example for marshland birds 1 2001 ... 空间显式模型的独特性体现在其建模过程中可以联合不同的理论模型从而允许研究者考虑各种动态过程和模拟情景.例如, 通过与基于个体的METAPHOR模型(Verboom et al, 2001; Vos et al, 2001)的结合使用, 空间显式模型可以将保护区的大小、形状、隔离度等这些被认为是决定物种生存的关键因素纳入分析(Ricketts, 2001; Tischendorf et al, 2003; Schippers et al, 2009), 以更真实地反映不同物种在复杂景观中的动态行为.其次, 空间显式模型还可以与物种运动模型相结合, 这对于精确评估生境破碎化景观中的SLOSS问题至关重要.比如, Niebuhr等(2015)将多尺度动物运动模型和空间显式的景观结构结合起来探讨了栖息地搜索行为下的SLOSS争论, 并表明当物种死亡事件为零时, 几个小斑块的景观比单个大斑块景观更适合动物生存.综上, 空间显式模型与其他分析模型整合的方式为解决SLOSS争论以及保护区的设计和管理提供了更为全面和细致的视角.为了更好地理解物种生存力对生境破碎化的响应(Kanagaraj et al, 2013; Niebuhr et al, 2015), 未来在空间显式模型的应用上应该进一步将栖息地质量、基质组成等相关特征考虑在内. ... The aggregation pheromones of bark beetles: Progress and problems 1 1976 ... 事实上, 大多数物种往往呈现聚集分布的趋势(Seidler & Plotkin, 2006; Morlon et al, 2008).这种趋势由多种因素引起, 包括抵御入侵种或共同寻找食物资源时的集群以减少死亡率, 比如冬季哺乳动物个体之间的社会联系来定位觅食区域或对抗(Peignier et al, 2019), 雀形目中雌鸟的筑巢位置更靠近边界(Ramsay et al, 1999)等, 这些统称为同种吸引(Vité & Francke, 1976; Schuck-Paim & Alonso, 2001)和恋巢性(Weatherhead & Forbes, 1994).然而, 动物倾向于聚集分布的主要驱动力或许来自它们对部分微生境(局部地形、小气候)的响应(详见Fahrig等(2022)的综述论文).当区域因栖息地破碎化或土地清理等过程形成几个小斑块时, 与少数大斑块相比, 前者将与那些在斑块形成之前就已经存在的物种更广泛地相交(图5), 这增加了SS中的β多样性, 因此可以预测SS > SL.Saravia等(2025)的最新研究在运用多物种空间显式模型探讨SLOSS问题时就明确考了虑物种数量稳定后的栖息地破碎化, 这种情况被认为比所有物种最初以随机模式出现在破碎的栖息地中更现实(Rybicki et al, 2020).正如立方体假说中所提到的那样, 物种在破碎化之前的空间分布格局越异质性, 这种类比就越重要(Saravia et al, 2025). ... Toward ecologically scaled landscape indices 1 2001 ... 空间显式模型的独特性体现在其建模过程中可以联合不同的理论模型从而允许研究者考虑各种动态过程和模拟情景.例如, 通过与基于个体的METAPHOR模型(Verboom et al, 2001; Vos et al, 2001)的结合使用, 空间显式模型可以将保护区的大小、形状、隔离度等这些被认为是决定物种生存的关键因素纳入分析(Ricketts, 2001; Tischendorf et al, 2003; Schippers et al, 2009), 以更真实地反映不同物种在复杂景观中的动态行为.其次, 空间显式模型还可以与物种运动模型相结合, 这对于精确评估生境破碎化景观中的SLOSS问题至关重要.比如, Niebuhr等(2015)将多尺度动物运动模型和空间显式的景观结构结合起来探讨了栖息地搜索行为下的SLOSS争论, 并表明当物种死亡事件为零时, 几个小斑块的景观比单个大斑块景观更适合动物生存.综上, 空间显式模型与其他分析模型整合的方式为解决SLOSS争论以及保护区的设计和管理提供了更为全面和细致的视角.为了更好地理解物种生存力对生境破碎化的响应(Kanagaraj et al, 2013; Niebuhr et al, 2015), 未来在空间显式模型的应用上应该进一步将栖息地质量、基质组成等相关特征考虑在内. ... A review on the nested distribution pattern (nestedness): Analysis methods, mechanisms and conservation implications 2 2023 ... SLOSS问题的核心在于理解和评估不同保护区配置对物种多样性的影响.自20世纪70年代以来, 生态学家和保护生物学家一直关注于SLOSS争论的不同方面.早期的研究主要集中在各生物类群的的数据分析, 例如通过调查不同生物群系的物种数并运用种-面积关系来探讨保护区大小和数量对物种丰富度的影响(Connor & Abele, 1979).此外, 一些理论学家早就意识到不同区域之间的物种组成是解决SLOSS问题的关键(Simberloff & Abele, 1976; Higgs & Usher, 1980; Tj?rve, 2010), 即如果小保护区中的物种组成完全是大保护区中的物种组成的一个嵌套子集(Patterson & Atmar, 1986; Wang et al, 2023), 那么一个大的保护区会比总面积相等的几个小的保护区保护的物种数更多.主要原因如下: 其一, 如果单个大保护区和几个小保护区的物种组成没有差异(或者说物种组成差异仅由面积决定, 则表明小保护区中的物种完全是大保护区中物种的嵌套子集), 那么SL > SS; 其二, 若几个小保护区与单个连续的大保护区的物种组成存在差异(几个小保护区并不具有嵌套的物种组成), 那么可以从小保护区之间共有物种数的角度来讨论SLOSS问题, 即若保护区之间共有物种数较多, 那么SL > SS, 否则SS > SL (Tj?rve, 2010).然而, Boecklen (1997)利用148组岛屿物种分布数据检验了物种嵌套程度对于塑造SLOSS模式(SS或SL)的作用, 发现嵌套性并不能为解决SLOSS问题提供可靠的证据.进入21世纪以后, 随着生态建模技术的进步, 学者们开始运用理论模型并从多尺度生态过程(如扩散限制、灭绝-定殖理论)的视角出发, 针对如何使生态指标(如物种数量、灭绝率)最优的保护目的来确定保护区的空间配置(Groeneveld, 2005), 包括景观层面的生态学模型、种群动态学模型以及空间模型(Ovaskainen, 2002; Pichancourt et al, 2006; McCarthy et al, 2011), 这些生态学模型的发展推动了SLOSS争论在理论层面的深化.尽管有些研究在不同条件下可能会得出相同的结论, 但SLOSS仍然被描述为一场辩论, 且一直处于争议之中(Le Roux et al, 2015; Kendal et al, 2017; MacDonald et al, 2018). ...
大多数关于SLOSS争论的理论模型往往只考虑如何使生态效益最大化, 常忽略了经济因素在SLOSS争论中的重要性(Groeneveld, 2005), 包括经济成本、财政支出等, 而经济模型则综合考虑了生态效益和经济效益, 从而能够在资源有限的情况下对其进行合理分配以达到预期目标. ... Natal philopatry in passerine birds: Genetic or ecological influences? 1 1994 ... 事实上, 大多数物种往往呈现聚集分布的趋势(Seidler & Plotkin, 2006; Morlon et al, 2008).这种趋势由多种因素引起, 包括抵御入侵种或共同寻找食物资源时的集群以减少死亡率, 比如冬季哺乳动物个体之间的社会联系来定位觅食区域或对抗(Peignier et al, 2019), 雀形目中雌鸟的筑巢位置更靠近边界(Ramsay et al, 1999)等, 这些统称为同种吸引(Vité & Francke, 1976; Schuck-Paim & Alonso, 2001)和恋巢性(Weatherhead & Forbes, 1994).然而, 动物倾向于聚集分布的主要驱动力或许来自它们对部分微生境(局部地形、小气候)的响应(详见Fahrig等(2022)的综述论文).当区域因栖息地破碎化或土地清理等过程形成几个小斑块时, 与少数大斑块相比, 前者将与那些在斑块形成之前就已经存在的物种更广泛地相交(图5), 这增加了SS中的β多样性, 因此可以预测SS > SL.Saravia等(2025)的最新研究在运用多物种空间显式模型探讨SLOSS问题时就明确考了虑物种数量稳定后的栖息地破碎化, 这种情况被认为比所有物种最初以随机模式出现在破碎的栖息地中更现实(Rybicki et al, 2020).正如立方体假说中所提到的那样, 物种在破碎化之前的空间分布格局越异质性, 这种类比就越重要(Saravia et al, 2025). ... 1 1986 ... Concepts related to the SLOSS debate
本文的其它图/表
|




{kind=link}