SLOSS争论研究进展: 分析方法、理论机制及保护实践
章洋, 王彦平
生物多样性
2025, 33 ( 7):
25081-.
DOI: 10.17520/biods.2025081
生物多样性保护领域中的SLOSS (single large or several small)争论概述了在总面积相等或资源有限的情况下, 应该优先保护一个大的保护区(SL)还是几个小的保护区(SS)来最大化保护物种多样性的问题。自20世纪70年代以来, 该问题一直备受生态学家和保护生物学家的关注, 其研究对于保护区设计以及生物多样性保护具有重要意义。随着研究方法的发展和交叉学科的兴起, SLOSS争论在分析方法和理论机制方面都取得了重要的阶段性进展。本文通过文献检索和归纳总结对SLOSS争论的研究现状进行了综述。首先, 我们回顾了SLOSS争论的起源及其对生物多样性保护的重要性。其次, 本文概括评估了SLOSS争论在实证研究和理论方面的分析方法, 包括零模型法、经典的物种累积曲线法以及由该方法所计算出来的饱和指数。随后, 本文还回顾了以往研究者提出的改进的饱和指数, 并介绍了我们所开发的优化后的饱和指数。此外, 本文探讨了几种影响SLOSS争论的理论机制, 如由灭绝率和定殖率的变化所主导的灭绝-定殖动态理论机制及其相应的检验方法。最后, 本文对该领域进行了总结和展望并为未来的发展方向提出了一些针对性建议, 包括采用多种保护策略的重要性以及在保护区规划中经济和生态因素的结合的必要性。本文整合了SLOSS争论中常用的分析方法的适用场景与局限性, 比较了不同方法在划分保护优先级时的优势和争议点, 可为研究者在方法选择及保护规划权衡方面提供参考。
理论假说
Theoretical hypothesis | 预测
Pridictions | 潜在机制
Potential mechanisms | I. 灭绝-定殖动态 Extinction-colonization dynamics
假设1: 灭绝率的变化主导灭绝-定殖动态理论(Fahrig et al, 2022)。物种在斑块中的灭绝率随着斑块的减小而增大。Assumption 1: Changes in extinction rates dominate the theory of extinction-colonization dynamics (Fahrig et al, 2022). Species extinction rates in each patch increase with decreasing patch size. | SL > SS | (1)种群统计学的随机性对于大斑块来说相对较弱(Jagers & Harding, 2009; Tjørve, 2010); (2)不同物种的最小斑块大小需求不同(Patterson & Atmar, 1986; McCarthy et al, 2006); (3)小斑块的总边缘更长, 使得小斑块中的物种迁移率高于大斑块, 导致SS在不利基质中的扩散死亡率高于SL (Flather & Bevers, 2002; Martin & Fahrig, 2016); (4)负的边缘效应强化了前两种机制, 增加了小斑块中物种的灭绝风险(Williams et al, 2005; Moilanen & Wintle, 2007)。
(1) The randomness of population statistics is relatively weak for large patchs (Jagers & Harding, 2009; Tjørve, 2010). (2) Different species have different minimum patch size requirements (Patterson & Atmar, 1986; McCarthy et al, 2006). (3) The total edge length of small patches is longer, resulting in higher species migration rates within small patches than in large patches, leading to higher dispersal mortality rates of SS in unfavourable substrates than in SL (Flather & Bevers, 2002; Martin & Fahrig, 2016). (4) The negative edge effect reinforces the first two mechanisms, increasing the risk of extinction for species in small patches (Williams et al, 2005; Moilanen & Wintle, 2007). | | 假设2: 定殖率的变化主导灭绝-定殖动态理论(Fahrig et al, 2022)。物种在SL中的定殖率小于SS。Assumption 2: Changes in colonization rates dominate the theory of extinction-colonization dynamics (Fahrig et al, 2022). Species colonization rate in SL is lower than SS. | SS > SL | (1)由于SS的边缘与面积的比值更大, 促使SS遇到新物种的概率也更大(Bowman et al, 2002; Fahrig et al, 2011; Puckett & Eggleston, 2016); (2) SS可获得的物种库也有望大于SL, 因为SS在特定范围内的栖息地总量高于SL (Preston, 1962; Fahrig, 2013)。
(1) Since the ratio of SS edges to area is greater, the probability of SS encountering new species is also greater (Bowman et al, 2002; Fahrig et al, 2011; Puckett & Eggleston, 2016). (2) The species pool available to SS is also expected to be larger than that of SL, because SS has a greater total habitat area within a specific range than SL (Preston, 1962; Fahrig, 2013). | II. 风险抵抗作用 Risk resistance effect
SS相较于SL抵御风险的能力更强 SS is more resilient to risk compared to SL | SS > SL | (1)竞争者或捕食者等敌对物种在斑块之间较少移动, 导致弱小物种能够躲避到SS (Levins, 1969; Simberloff & Abele, 1976; Morrison & Barbosa, 1987; Deane et al, 2020); (2)干扰不会通过基质传播, 促使SS将风险扩散到整个景观中(Levins, 1969; Kallimanis et al, 2005; Fahrig, et al, 2022)。
(1) Competitors or predators and other hostile species move less between patches, allowing weaker species to escape to SS (Levins, 1969; Simberloff & Abele, 1976; Morrison & Barbosa, 1987; Deane et al, 2020). (2) Interference does not spread through the matrix, causing SS to spread the risk throughout the landscape (Levins, 1969; Kallimanis et al, 2005; Fahrig et al, 2022). | Ⅲ. β多样性和物种分布的聚集性 β diversity and clumped distributions of species
SS中的β多样性比SL更高(Fahrig et al, 2022) β diversity is higher in SS than in SL (Fahrig et al, 2022) | SS > SL | (1)物种在空间上主要呈聚集分布, 这是由于同种吸引、恋巢性和生境异质性。当生境被清理而形成斑块时, SS在空间上将与更多的物种相交(Diamond, 1975; Higgs & Usher, 1980; Tjørve, 2010; Lasky & Keitt, 2013; May et al, 2019); (2) SS之间的跨生境异质性和β多样性均高于SL (Ewers et al, 2013, Fahrig et al, 2022)。
(1) Species tend to cluster spatially, mainly due to conspecific affinity, nest loyalty, and habitat heterogeneity. When habitats are cleared and patches are formed, SS will intersect with more species spatially (Diamond, 1975; Higgs & Usher, 1980; Tjørve, 2010; Lasky & Keitt, 2013; May et al, 2019). (2) The cross-habitat heterogeneity and β diversity between SS are higher than those between SL (Ewers et al, 2013, Fahrig, et al, 2022). |
View table in article
表4
预测SLOSS模式的理论假说和机制(修改自Fahrig等, 2022)。SL: 一个大的保护区; SS: 几个小的保护区。
正文中引用本图/表的段落
在上述公式中, Lj为物种所占据斑块的面积的对数, $\bar{L}$是对数转换后的斑块面积的平均值。当η为负值时, 表明SS占优势; 当η为正值时, 表明SL占优势。然而, η指数的缺陷之一是不能为功能和谱系层面的SLOSS研究提供有用的信息, 另外当将η指数的观测值和蒙特卡洛置换检验产生的随机η值进行比较时可能会面临P值不显著的问题(附录4)。
为了解释几个小保护区为什么总是比单一大保护区拥有更多的物种, 理论已经预测了大量期望SS > SL或SL > SS出现的条件(表4) (Fahrig, 2020)。然而, 这些预测并未与实证数据相结合或与其进行对比。另外, 与SLOSS争论相关的大多数实证研究的结果一致表明SS > SL占据主导地位, 比如Riva和Fahrig (2022)系统地收集了32个数据集, 然后利用SLOSS比较分别对这些数据集中的所有物种以及IUCN红色名录(IUCN, 2021)中显示的数量下降的物种进行分析后发现, 所有32个数据集的SLOSS比较都支持SS > SL, 并且对14个数据集中的数量下降的物种的分析也呈现出了类似的结果: 12个数据集支持SS > SL, 2个数据集支持SS = SL。鉴于此, 解决SLOSS争论的关键步骤是确定哪种理论机制导致了SS > SL模式的频繁出现(Fahrig, 2020)。深入理解SLOSS模式的形成机制对于生物多样性保护具有重要意义。
本文的其它图/表
-
 表1
与SLOSS争论相关的概念
表1
与SLOSS争论相关的概念
-
 图1
物种累积曲线法(SLOSS比较法)示意图。在给定的栖息地总面积下, (a)从小到大的曲线完全高于从大到小的曲线, 代表SS > SL; (b)从大到小的曲线完全在从小到大曲线的下方, 代表SL > SS; (c)两条曲线交叉(inconclusive), 代表无法确定。
图1
物种累积曲线法(SLOSS比较法)示意图。在给定的栖息地总面积下, (a)从小到大的曲线完全高于从大到小的曲线, 代表SS > SL; (b)从大到小的曲线完全在从小到大曲线的下方, 代表SL > SS; (c)两条曲线交叉(inconclusive), 代表无法确定。
-
表2
现有饱和指数和SLOSS指数及其特征
-
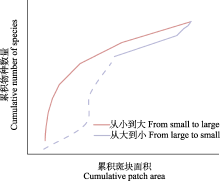 图2
累积种-面积关系曲线示意图。数据点之间的线段按S = cAz进行非线性插值而来, 虚线代表从大到小曲线上的最大斑块面积对应的点与原点的非线性累积趋势。
图2
累积种-面积关系曲线示意图。数据点之间的线段按S = cAz进行非线性插值而来, 虚线代表从大到小曲线上的最大斑块面积对应的点与原点的非线性累积趋势。
-
 图3
用于表征从小到大累积曲线(红色)和从大到小累积曲线(灰色)之间偏差的ξ统计量以及指标ΔA示意图。Ψ代表从最大斑块面积推导出的两条曲线下面积的差值; ΔA表示估算两个面积的区域范围。图(a)表示ξ统计量为正的情况, 图(b)代表ξ统计量为负的情况。
图3
用于表征从小到大累积曲线(红色)和从大到小累积曲线(灰色)之间偏差的ξ统计量以及指标ΔA示意图。Ψ代表从最大斑块面积推导出的两条曲线下面积的差值; ΔA表示估算两个面积的区域范围。图(a)表示ξ统计量为正的情况, 图(b)代表ξ统计量为负的情况。
-
 图4
推导ISU和IDI指数的示意图。(a)表示直接将黄色区域面积减去或除以黄色区域和紫色区域面积之和的情况; (b)表示将所有黄色区域的面积相加再对全部的黄色区域和紫色区域面积的总和作减法或除法的情况。
图4
推导ISU和IDI指数的示意图。(a)表示直接将黄色区域面积减去或除以黄色区域和紫色区域面积之和的情况; (b)表示将所有黄色区域的面积相加再对全部的黄色区域和紫色区域面积的总和作减法或除法的情况。
-
表3
现有理论模型及目标评估
-
 图5
当物种呈聚集分布时, 少量大斑块相交的物种数量(a)少于几个小斑块(b)和(c)。不同的颜色及相应的形状代表生境丧失前单个连续栖息地中的不同物种。黑色方框代表因生境丧失而形成的斑块。当少数大斑块和几个小斑块的斑块边缘之间的最大距离相等或类似时, 几个小斑块集能更均匀地覆盖在整个景观中, 因此相交的物种数更多(Fahrig et al, 2022)。(a)中2个大斑块与3个物种相交, 然而(b)中5个小斑块与6个物种相交。在斑块之间相距较远的情况下, 这种效应还会放大: (c)中5个小斑块相交了11个物种(该图改自Fahrig等, 2022)。
图5
当物种呈聚集分布时, 少量大斑块相交的物种数量(a)少于几个小斑块(b)和(c)。不同的颜色及相应的形状代表生境丧失前单个连续栖息地中的不同物种。黑色方框代表因生境丧失而形成的斑块。当少数大斑块和几个小斑块的斑块边缘之间的最大距离相等或类似时, 几个小斑块集能更均匀地覆盖在整个景观中, 因此相交的物种数更多(Fahrig et al, 2022)。(a)中2个大斑块与3个物种相交, 然而(b)中5个小斑块与6个物种相交。在斑块之间相距较远的情况下, 这种效应还会放大: (c)中5个小斑块相交了11个物种(该图改自Fahrig等, 2022)。
|
