SLOSS争论研究进展: 分析方法、理论机制及保护实践
章洋, 王彦平
生物多样性
2025, 33 ( 7):
25081-.
DOI: 10.17520/biods.2025081
生物多样性保护领域中的SLOSS (single large or several small)争论概述了在总面积相等或资源有限的情况下, 应该优先保护一个大的保护区(SL)还是几个小的保护区(SS)来最大化保护物种多样性的问题。自20世纪70年代以来, 该问题一直备受生态学家和保护生物学家的关注, 其研究对于保护区设计以及生物多样性保护具有重要意义。随着研究方法的发展和交叉学科的兴起, SLOSS争论在分析方法和理论机制方面都取得了重要的阶段性进展。本文通过文献检索和归纳总结对SLOSS争论的研究现状进行了综述。首先, 我们回顾了SLOSS争论的起源及其对生物多样性保护的重要性。其次, 本文概括评估了SLOSS争论在实证研究和理论方面的分析方法, 包括零模型法、经典的物种累积曲线法以及由该方法所计算出来的饱和指数。随后, 本文还回顾了以往研究者提出的改进的饱和指数, 并介绍了我们所开发的优化后的饱和指数。此外, 本文探讨了几种影响SLOSS争论的理论机制, 如由灭绝率和定殖率的变化所主导的灭绝-定殖动态理论机制及其相应的检验方法。最后, 本文对该领域进行了总结和展望并为未来的发展方向提出了一些针对性建议, 包括采用多种保护策略的重要性以及在保护区规划中经济和生态因素的结合的必要性。本文整合了SLOSS争论中常用的分析方法的适用场景与局限性, 比较了不同方法在划分保护优先级时的优势和争议点, 可为研究者在方法选择及保护规划权衡方面提供参考。
| 理论模型划分 Category of theoretical models | 特征 Characteristics | 目标预测 Target predictions | 研究案例 Case studies | | 经济模型 Economic models | 同时考虑生态目标和经济成本 Considering both ecological objectives and economic costs | 综合生态和经济效益设计最优保护区 Integration of ecological and economic benefits in the design of optimal protected areas | Groeneveld, 2005; Helmstedt et al, 2014 | | 空间方差结构的超种群模型 Super-population model of the spatial variance structure | 可分析误差范围、物种的空间分布和经济成本 Ranges of error, spatial distribution of species and economic costs can be analyzed | 最大成本限制下的最优保护区配置 Optimal protected area allocation under maximum cost constraints | Picard et al, 2010 | | 集合种群模型 Metapopulation model | 特别关注物种的扩散和迁移 Particularly concerned about the dispersal and movement of species | 物种灭绝风险或种群持久性 Species extinction risk or population persistence | Ovaskainen, 2002; McCarthy et al, 2005; Zhou & Wang, 2006 | | 随机灭绝模型 Stochastic extinction model | 允许随机性过程如随机灭绝事件的发生 Allow for stochastic processes such as random extinction events | 灭绝风险 Extinction risk | Wright & Hubbell, 1983; Burkey, 1989 | | 微观实验模型 Microscopic experimental model | 模式生物为原生生物且实验条件可控 Model organisms are protists and experimental conditions are controlled | 物种灭绝数量 Number of extinct species | Hammill & Clements, 2020; Wolfe et al, 2022 | | 空间隐式模型 Spatially implicit model | 可识别联系种群和景观的一般模式 General patterns of linked populations and landscapes can be recognized | 种群生存能力 Population viability | Pichancourt et al, 2006 | | 计算机模拟模型 Computer simulation model | 允许调整多个参数以模拟不同的生态场景 Allow adjusting multiple parameters to simulate different ecological scenarios | 种群生存能力 Population viability | Liu et al, 2017; Pelletier, 2000 | | 物种分布模拟模型 Distribution simulated model | 允许改变物种分布的不同组成部分 Allow changes to the different components of species distribution | 物种存活和灭绝概率 Species survival and extinction probabilities | May et al, 2019 | | 空间分配模型 Spatial allocation model | 可应用于不同类型的斑块和物种 Can be applied to different types of patches and species | 种群可持续性 Population sustainability | van Langevelde et al, 2002 | | 空间均衡模型 Spatial equilibrium model | 关注生态、经济和环境政策因素 Ecological, economic, and environmental policy factors are concerned | 灭绝风险 Extinction risk | Eppink & Withagen, 2005 | | 基于个体的模型 Individual-based model | 强调个体行为和运动对种群动态的影响 Emphasize the effects of individual behavior and movement on population dynamics | 种群规模 Population size | Fahse et al, 1998; Nakajima &
Boggs, 2015 | | 空间显式模型 Spatially explicit model | 关注物种在空间上的扩散且可与其他模型结合使用 Focus on spatial dispersal of species and can be used in conjunction with other models | 种群持久性 Population persistence | Robert, 2009; Evans et al, 2013; Niebuhr et al, 2015 | | 多斑块模型 Multi-patch model | 可调整物种迁移率的变化 Can adjust the changes in species migration rate | 种群规模 Population size | Elbetch et al, 2022 | | 渔业模型 Fishery model | 可应用于海洋保护区且通常涉及捕捞努力的影响 Can be applied to marine protected areas and typically involves the impact of fishing efforts | 种群规模 Population size | Kritzer, 2004; Moussaoui & Auger, 2015 | | 生态系统模型 Ecosystem model | 允许在广泛的空间尺度和各种类型的情景下运行 Allow operation in a wide range of spatial scales and various types of scenarios | 综合生态和经济效益设计最优保护区 Integration of ecological and economic benefits in the design of optimal protected areas | Salomon et al, 2002 | | 景观模拟模型 Landscape simulation model | 允许评估景观指标的变化且可模拟多种场景 Allow for the evaluation of changes in landscape indicators and can simulate multiple scenarios | 灭绝风险和生物多样性 Extinction risk and biodiversity | Watts & Hughes, 2024 |
View table in article
表3
现有理论模型及目标评估
正文中引用本图/表的段落
随着相关研究数量的不断增加, 亟需对该领域重新回顾以深入了解现有研究进展和知识空白。因此, 我们以“SLOSS” “SLOSS debate” “single large or several small”为关键词,在Web of Science和Google Scholar等数据库中进行了文献检索, 并筛选出了截至2024年12月发表的所有涉及到SLOSS争论的研究。然后, 我们对每篇文献逐一查看, 通过检查出版物的标题、摘要和主要内容, 删除了不相关的文献、重复内容、评论和综述论文。随后, 我们对剩余文献进行仔细查阅, 只有提供了SLOSS争论的常规方法或理论模型并给出了最后结果的论文才会被保留进入后续分析。最终, 我们总共获得了210篇关于SLOSS争论的实证(附录1)和理论(附录2)研究论文。其中, 使用了具体的数据集的论文有151篇, 这些数据集所涵盖的地理区域主要集中在北美洲、南美洲和大洋洲, 而其他各洲的研究相对较少(附录3)。从国家分布来看, 美国的研究数量最多, 澳大利亚和巴西分别占据第二和第三名, 墨西哥、英国、加拿大等也发表了一些研究(附录3)。从研究时间来看, 关于SLOSS争论的研究在2006年达到顶峰, 此后逐渐下降, 随后在2015年到2022年期间又大致呈现上升趋势(附录3)。这可能与生境破碎化本身对物种多样性的影响以及新的SLOSS指数被提出有关(Gavish et al, 2012; Fahrig, 2017)。
Mac Nally和Lake (1999)注意到, SS和SL曲线下面积的计算均是从最小累积面积所对应的点开始, 并且SL曲线的下限对应于最大斑块的面积。其次, 两条曲线的上限是相同的, 也即所有斑块的总累积面积相等。因此, SL曲线下的面积主要依赖于最大斑块的面积。为了减轻这种依赖性, 他们建议从最大斑块的面积到所有斑块的总累积面积来计算区间内的积分。这些积分的差值代表图3中Ψ符号所示的面积。之后, 他们用Ψ除以两个积分的面积范围ΔA得到ξ统计量, 即ξ = Ψ/ΔA (图3)。ξ统计量表示SS曲线和SL曲线在ΔA区间内的累积丰富度加权的差异(Mac Nally & Lake, 1999)。若SS曲线位于SL曲线之上, 则ξ统计量为正, 表明几个小斑块的平均累积物种丰富度超过单个大的斑块, 若ξ统计量为负, 则为相反的情况(Mac Nally & Lake, 1999)。
在研究早期, Wright和Hubbell (1983)就已经将理论模型法应用于SLOSS分析。他们探讨了单个大型和几个小型保护区对物种灭绝概率的影响, 发现一个大的保护区对于减小物种的灭绝风险更有利。此后, 日趋增多的理论模型被相继提出(表3), 同时也成为了理论分析SLOSS问题的重要对策。目前, 对于理论模型的构建主要围绕3个方面: 最小化物种的灭绝风险、延长种群持久性以及扩大单一物种的种群规模(Groeneveld, 2005)。虽然不同模型的前提假设各不相同, 但这些模型都是在总面积相等的情况下进行条件模拟, 然后针对以上几个方面的其中一个或两个进行探讨。在本文中, 我们重点介绍了5种有代表性的理论模型: 经济模型、集合种群模型、空间显式模型、渔业模型、微观实验模型。
近年来, 预测SLOSS模式的理论机制已经成为一个重点研究议题。例如, Fahrig (2022)总结了以往的SLOSS预测和实证研究后提出了一个“立方体假说” (SLOSS cube hypothesis), 该立方体的3个坐标轴分别代表物种在斑块之间的移动频率、风险扩散作用以及量化跨生境异质性水平的物种聚集指数。这意味着SLOSS预测在通常情况下可以由以上3个条件的组合来表征。在SLOSS立方体内, SS > SL、SL > SS和SS = SL所占据的体积各不相同。其中, SS > SL在立方体中占据的体积最大, 其次是SS = SL, 最后为SL > SS, 即关于SLOSS研究的实证工作的结果主要支持几个小保护区且有相当一部分的研究没有发现SS和SL策略之间的明显差异, 而单个大保护区更好的结果则相对罕见(Fahrig, 2022)。
(4)在多样性维度上, 以往有关SLOSS争论的研究几乎只涉及物种多样性层面, 还很少有研究涉及到功能和谱系多样性(La Sorte et al, 2023)。现有证据表明功能多样性不能很好地代表谱系多样性(Calba et al, 2014), 而谱系多样性也不能可靠地反映功能多样性(Mazel et al, 2018), 这表明这两个指标可能会有不同的结果。因此, 我们建议以后应同时从物种、功能和谱系3个维度进行SLOSS研究, 以便更深入地理解SLOSS争论。
大多数关于SLOSS争论的理论模型往往只考虑如何使生态效益最大化, 常忽略了经济因素在SLOSS争论中的重要性(Groeneveld, 2005), 包括经济成本、财政支出等, 而经济模型则综合考虑了生态效益和经济效益, 从而能够在资源有限的情况下对其进行合理分配以达到预期目标. ... Difficult decisions: Strategies for conservation prioritization when taxonomic, phylogenetic and functional diversity are not spatially congruent 1 2018 ... 生物多样性的维持与保护是当今世界面临的最重要的挑战之一(Cadotte & Tucker, 2018).在保护规划的理论框架中, 备受关注的SLOSS争论描述了在总面积固定不变或资源有限的情况下, 应该优先选择单个大的保护区(single large, SL)还是几个小的保护区(several small, SS)来最大化保护物种多样性的问题.SLOSS争论起源于Diamond (1975)关于保护区设计的论文, 该文将岛屿生物地理学理论应用到保护区设计和规划中, 并基于面积和隔离度对物种多样性的影响, 提出在限定总面积的前提下, 单个大的保护区是比几个小的保护区更好的保护区设计策略, 即“SL > SS”原则.然而, 该法则随后便受到Simberloff和Abele (1976)的反对, 他们指出, 岛屿生物地理学理论并未对SLOSS问题提供明确的答案, 而且他们认为Diamond (1975)的论文没有理论依据, 也没有实际数据来支撑小保护区的灭绝风险较高的论点.此后, 支持“SL > SS”观点的学者便与反对者展开了激烈的对峙, SLOSS争论由此形成, 同时也成为了保护生物学领域中最具争论的理论问题之一(Moussaoui & Auger, 2015). ... Measuring and explaining large-scale distribution of functional and phylogenetic diversity in birds: Separating ecological drivers from methodological choices 1 2014 ... (4)在多样性维度上, 以往有关SLOSS争论的研究几乎只涉及物种多样性层面, 还很少有研究涉及到功能和谱系多样性(La Sorte et al, 2023).现有证据表明功能多样性不能很好地代表谱系多样性(Calba et al, 2014), 而谱系多样性也不能可靠地反映功能多样性(Mazel et al, 2018), 这表明这两个指标可能会有不同的结果.因此, 我们建议以后应同时从物种、功能和谱系3个维度进行SLOSS研究, 以便更深入地理解SLOSS争论. ... On random placement and species-area relations 1 1981 ... 为了确定物种累积的潜在生态机制, 包括最大化从小到大方向增加物种的可能性, 区分物种占用对斑块大小的依赖性等, 有必要控制任何采样因素对种-面积关系的影响, 这可以通过零模型来实现(Deane, 2022).最简单的解释是假设种-面积关系是一种被动取样现象(Connor & McCoy, 1979), 即在一个斑块中共同观察到一个物种的概率取决于样本的大小以及物种在景观中的相对多度(Arrhenius, 1921; Coleman, 1981).但是, 如果我们假设占用率与物种多度呈正相关, 则可以使用固定总丰富度和总占用率(保持每个地点的行及每个物种的列的总和恒定)的算法作为物种存在-缺失矩阵的零模型来进行预测(Deane, 2022). ... 1 1979 ... SLOSS问题的核心在于理解和评估不同保护区配置对物种多样性的影响.自20世纪70年代以来, 生态学家和保护生物学家一直关注于SLOSS争论的不同方面.早期的研究主要集中在各生物类群的的数据分析, 例如通过调查不同生物群系的物种数并运用种-面积关系来探讨保护区大小和数量对物种丰富度的影响(Connor & Abele, 1979).此外, 一些理论学家早就意识到不同区域之间的物种组成是解决SLOSS问题的关键(Simberloff & Abele, 1976; Higgs & Usher, 1980; Tj?rve, 2010), 即如果小保护区中的物种组成完全是大保护区中的物种组成的一个嵌套子集(Patterson & Atmar, 1986; Wang et al, 2023), 那么一个大的保护区会比总面积相等的几个小的保护区保护的物种数更多.主要原因如下: 其一, 如果单个大保护区和几个小保护区的物种组成没有差异(或者说物种组成差异仅由面积决定, 则表明小保护区中的物种完全是大保护区中物种的嵌套子集), 那么SL > SS; 其二, 若几个小保护区与单个连续的大保护区的物种组成存在差异(几个小保护区并不具有嵌套的物种组成), 那么可以从小保护区之间共有物种数的角度来讨论SLOSS问题, 即若保护区之间共有物种数较多, 那么SL > SS, 否则SS > SL (Tj?rve, 2010).然而, Boecklen (1997)利用148组岛屿物种分布数据检验了物种嵌套程度对于塑造SLOSS模式(SS或SL)的作用, 发现嵌套性并不能为解决SLOSS问题提供可靠的证据.进入21世纪以后, 随着生态建模技术的进步, 学者们开始运用理论模型并从多尺度生态过程(如扩散限制、灭绝-定殖理论)的视角出发, 针对如何使生态指标(如物种数量、灭绝率)最优的保护目的来确定保护区的空间配置(Groeneveld, 2005), 包括景观层面的生态学模型、种群动态学模型以及空间模型(Ovaskainen, 2002; Pichancourt et al, 2006; McCarthy et al, 2011), 这些生态学模型的发展推动了SLOSS争论在理论层面的深化.尽管有些研究在不同条件下可能会得出相同的结论, 但SLOSS仍然被描述为一场辩论, 且一直处于争议之中(Le Roux et al, 2015; Kendal et al, 2017; MacDonald et al, 2018). ... The statistics and biology of the species-area relationship 1 1979 ... 为了确定物种累积的潜在生态机制, 包括最大化从小到大方向增加物种的可能性, 区分物种占用对斑块大小的依赖性等, 有必要控制任何采样因素对种-面积关系的影响, 这可以通过零模型来实现(Deane, 2022).最简单的解释是假设种-面积关系是一种被动取样现象(Connor & McCoy, 1979), 即在一个斑块中共同观察到一个物种的概率取决于样本的大小以及物种在景观中的相对多度(Arrhenius, 1921; Coleman, 1981).但是, 如果我们假设占用率与物种多度呈正相关, 则可以使用固定总丰富度和总占用率(保持每个地点的行及每个物种的列的总和恒定)的算法作为物种存在-缺失矩阵的零模型来进行预测(Deane, 2022). ... The relationship between nested subsets, habitat subdivision, and species diversity 3 1995 ... Existing saturation and SLOSS indices and their characteristics
本文的其它图/表
-
 表1
与SLOSS争论相关的概念
表1
与SLOSS争论相关的概念
-
 图1
物种累积曲线法(SLOSS比较法)示意图。在给定的栖息地总面积下, (a)从小到大的曲线完全高于从大到小的曲线, 代表SS > SL; (b)从大到小的曲线完全在从小到大曲线的下方, 代表SL > SS; (c)两条曲线交叉(inconclusive), 代表无法确定。
图1
物种累积曲线法(SLOSS比较法)示意图。在给定的栖息地总面积下, (a)从小到大的曲线完全高于从大到小的曲线, 代表SS > SL; (b)从大到小的曲线完全在从小到大曲线的下方, 代表SL > SS; (c)两条曲线交叉(inconclusive), 代表无法确定。
-
表2
现有饱和指数和SLOSS指数及其特征
-
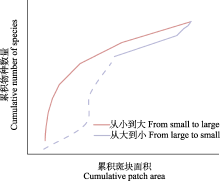 图2
累积种-面积关系曲线示意图。数据点之间的线段按S = cAz进行非线性插值而来, 虚线代表从大到小曲线上的最大斑块面积对应的点与原点的非线性累积趋势。
图2
累积种-面积关系曲线示意图。数据点之间的线段按S = cAz进行非线性插值而来, 虚线代表从大到小曲线上的最大斑块面积对应的点与原点的非线性累积趋势。
-
 图3
用于表征从小到大累积曲线(红色)和从大到小累积曲线(灰色)之间偏差的ξ统计量以及指标ΔA示意图。Ψ代表从最大斑块面积推导出的两条曲线下面积的差值; ΔA表示估算两个面积的区域范围。图(a)表示ξ统计量为正的情况, 图(b)代表ξ统计量为负的情况。
图3
用于表征从小到大累积曲线(红色)和从大到小累积曲线(灰色)之间偏差的ξ统计量以及指标ΔA示意图。Ψ代表从最大斑块面积推导出的两条曲线下面积的差值; ΔA表示估算两个面积的区域范围。图(a)表示ξ统计量为正的情况, 图(b)代表ξ统计量为负的情况。
-
 图4
推导ISU和IDI指数的示意图。(a)表示直接将黄色区域面积减去或除以黄色区域和紫色区域面积之和的情况; (b)表示将所有黄色区域的面积相加再对全部的黄色区域和紫色区域面积的总和作减法或除法的情况。
图4
推导ISU和IDI指数的示意图。(a)表示直接将黄色区域面积减去或除以黄色区域和紫色区域面积之和的情况; (b)表示将所有黄色区域的面积相加再对全部的黄色区域和紫色区域面积的总和作减法或除法的情况。
-
表4
预测SLOSS模式的理论假说和机制(修改自Fahrig等, 2022)。SL: 一个大的保护区; SS: 几个小的保护区。
-
 图5
当物种呈聚集分布时, 少量大斑块相交的物种数量(a)少于几个小斑块(b)和(c)。不同的颜色及相应的形状代表生境丧失前单个连续栖息地中的不同物种。黑色方框代表因生境丧失而形成的斑块。当少数大斑块和几个小斑块的斑块边缘之间的最大距离相等或类似时, 几个小斑块集能更均匀地覆盖在整个景观中, 因此相交的物种数更多(Fahrig et al, 2022)。(a)中2个大斑块与3个物种相交, 然而(b)中5个小斑块与6个物种相交。在斑块之间相距较远的情况下, 这种效应还会放大: (c)中5个小斑块相交了11个物种(该图改自Fahrig等, 2022)。
图5
当物种呈聚集分布时, 少量大斑块相交的物种数量(a)少于几个小斑块(b)和(c)。不同的颜色及相应的形状代表生境丧失前单个连续栖息地中的不同物种。黑色方框代表因生境丧失而形成的斑块。当少数大斑块和几个小斑块的斑块边缘之间的最大距离相等或类似时, 几个小斑块集能更均匀地覆盖在整个景观中, 因此相交的物种数更多(Fahrig et al, 2022)。(a)中2个大斑块与3个物种相交, 然而(b)中5个小斑块与6个物种相交。在斑块之间相距较远的情况下, 这种效应还会放大: (c)中5个小斑块相交了11个物种(该图改自Fahrig等, 2022)。
|
