中国石松类和蕨类植物多样性研究进展
王婷, 舒江平, 顾钰峰, 李艳清, 杨拓, 徐洲锋, 向建英, 张宪春, 严岳鸿
生物多样性
2022, 30 ( 7):
22381-.
DOI: 10.17520/biods.2022381
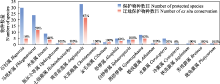
石松类和蕨类植物是维管植物的第二大类群, 其起源可追溯到4亿年前。在被子植物出现之前, 石松类和蕨类植物在古地球生态系统中占主导地位, 其重要性一直延续到现在。自20世纪40年代开始, 中国石松类和蕨类植物研究就令世界瞩目, 尤其是2017年第19届国际植物学大会在中国深圳召开后的5年时间里, 中国石松类和蕨类植物研究更是面向世界、走向国际, 研究更为广泛的科学问题, 在物种多样性、保护、系统演化和生态适应性等方面取得了一系列重要研究进展。2017-2022年, 多个中国研究团队利用多组学数据构建了世界石松类和蕨类植物科级水平的生命之树并提出了关键性状孢子囊环带演化的新模式; 解决了石松类和蕨类植物中目级、科级、属级和种级众多关键的系统分类学等问题, 发表了106个新分类群; 开展了大量的植物区系调查和研究, 出版了6部中国石松类和蕨类植物多样性专著和1部世界性专著; 对65种国家重点保护的石松类和蕨类植物进行了迁地保护, 同时实现了桫椤科、水蕨属(Ceratopteris)、观音座莲属(Angiopteris)和鹿角蕨(Platycerium wallichii)等重点保护类群的孢子繁殖; 在系统发育框架下, 研究了石松类和蕨类植物的生态修复功能和生态适应性演化。通过对2017-2022年研究成果的总结和思考, 本文对未来石松类和蕨类植物的发展提出以下建议: (1)提高中国寡型科属以及世界性大科大属的关注力度; (2)加强西藏、四川等薄弱地区石松类和蕨类植物的调查研究, 并结合新技术, 如DNA条形码等以提高区系调查中物种鉴别的效率和准确性; (3)运用多学科交叉的研究方法厘清各科、属、种间系统关系的同时, 还应加强系统和生态适应性演化之间的协同研究; (4)关注石松类和蕨类植物系统位置作为陆生维管植物演化起点的共性科学问题; (5)加强石松类和蕨类植物系统分类学与生态学、植物化学、保护生物学等学科间交叉合作研究。
| 科 Family | 种 Species | 文献 References | | 石松科 Lycopodiaceae | 蛇足石杉 Huperzia serrata | 焦瑜等, 2014 | | 粗糙马尾杉 Phlegmariurus squarrosus | 唐军等, 2015 | | 水韭科 Isoёtaceae | 中华水韭 Isoëtes sinensis | 路靖等, 2013; 邢建娇等, 2013 | | 云贵水韭 Isoëtes yunguiensis | 焦瑜等, 2014; 骆强等, 2016 | | 瓶尔小草科 Ophioglossaceae | 七指蕨 Helminthostachys zeylanica | 宋莉英等, 2015 | | 合囊蕨科 Marattiaceae | 福建观音座莲 Angiopteris fokiensis | 曾汉元和丁炳扬, 2003; 何俊等, 2021 | | 秦氏观音座莲 Angiopteris chingii | 焦瑜等, 2014 | | 相马氏观音座莲 Angiopteris somae | 焦瑜等, 2014 | | 圆基观音座莲 Angiopteris subrotundata | 焦瑜等, 2014 | | 河口观音座莲 Angiopteris hokouensis | 焦瑜等, 2014 | | 披针观音座莲 Angiopteris caudatiformis | 焦瑜等, 2014 | | 王氏观音座莲 Angiopteris wangii | 焦瑜等, 2014 | | 天星蕨 Christensenia assamica | 焦瑜等, 2014 | | 金毛狗科 Cibotiaceae | 金毛狗 Cibotium barometz | 张祖荣等, 2010; 邢建娇等, 2012; 王益和, 2012 | | 桫椤科 Cyatheaceae | 桫椤 Alsophila spinulosa | 王金娟等, 2007; 王辉等, 2013 | | 中华桫椤 Alsophila costularis | 王金娟等, 2007; 刘保东等, 2020 | | 粗齿桫椤 Alsophila denticulata | 曾汉元和丁炳扬, 2003 | | 笔筒树 Sphaeropteris lepifera | 马洪娜等, 2010 | | 白桫椤 Sphaeropteris brunoniana | 蒋胜军等, 2002; 王金娟等, 2007; 陈贵菊等, 2008 | | 海南白桫椤 Sphaeropteris hainanensis | 蒋胜军等, 2002 | | 滇南桫椤 Alsophila austroyunnanensis | 焦瑜等, 2014 | | 黑桫椤 Alsophila podophylla | 张祖荣等, 2010; 焦瑜等, 2014 | | 大叶黑桫椤 Alsophila gigantea | 焦瑜等, 2014; 郎月婷等, 2021 | | 阴生桫椤 Alsophila latebrosa | 焦瑜等, 2014; 郎月婷等, 2021 | | 凤尾蕨科 Pteridaceae | 荷叶铁线蕨 Adiantum reniforme | 黄芳, 2008; 李洪林等, 2008 | | 粗梗水蕨 Ceratopteris pteridoides | 孙锐等, 2008; 王文明等, 2018 | | 水蕨 Ceratopteris thalictroides | 戴锡玲等, 2005; 陈蔚辉等, 2008; 陈雨虹等, 2010; 朱晓凤等, 2016 | | 乌毛蕨科 Blechnaceae | 对开蕨 Asplenium komarovii | 刘保东等, 1993; 焦瑜等, 2014; 曾汉元和丁炳扬, 2003; 王蕾等, 2009; 韩汶雨等, 2020 | | 苏铁蕨 Brainea insignis | 王玥等, 2007; 郭建瑞等, 2008; 夏漪, 2014① | | 鹿角蕨科 Platyceriaceae | 鹿角蕨 Platycerium wallichii | 王卫清等, 2011; 郭捡等, 2012, 2013; 吴建设等, 2020; 叶秀仙等, 2020 |
View table in article
表5
孢子繁殖成功的国家重点保护石松类和蕨类植物
正文中引用本图/表的段落
我国对蕨类配子体的研究及繁育最早可以追溯到20世纪80年代的哈尔滨师范大学包文美教授, 其先后对问荆(Equisetum arvense)、卷柏(Selaginella tamariscina)及水龙骨科、岩蕨科、铁角蕨科、紫萁科、(原)中国蕨科、(原)铁线蕨科等蕨类植物进行了繁育研究。随后, 哈尔滨师范大学的刘保东教授和上海师范大学的王全喜教授继续对多种中国产的蕨类植物进行了孢子繁育和系统的配子体发育研究(严岳鸿和石雷, 2014)。近年来随着国家对生态文明建设的逐渐重视, 深圳市中国科学院仙湖植物园、中国科学院上海辰山植物园、中国科学院华南植物园、中国科学院西双版纳植物园、南京林业大学、西南林业大学等研究单位和高校都相继开展了石松类和蕨类植物的繁育工作, 其中孢子繁育研究主要集中在薄囊蕨类, 特别是外形美观、常用来做观赏植物开发的蕨类植物最多, 如凤尾蕨科、水龙骨科和铁角蕨科等, 其次是易于存活适合做园林搭配的蕨类植物, 如蹄盖蕨科、鳞毛蕨科、乌毛蕨科等。国家重点保护石松类和蕨类植物的孢子繁育近年来也取得了一定的研究成果(表5), 特别是桫椤科、水蕨属、鹿角蕨等植物, 这些类群植物的孢子繁育成活率已明显大大提高。另外, 福建观音座莲(Angiopteris fokiensis)的孢子萌发培养体系也在去年得到了新突破(何俊等, 2021), 填补了观音座莲属孢子萌发培养体系长期研究的空白, 为该属其他物种的保护、开发和利用提供了重要的理论基础和技术支持。与此同时, 还有一些演化较为原始或孑遗的石松类和蕨类植物, 如带状瓶尔小草、七指蕨、天星蕨、石杉属、马尾杉属等植物的孢子繁育瓶颈仍没有得到解决, 是后续需要重点关注的类群。
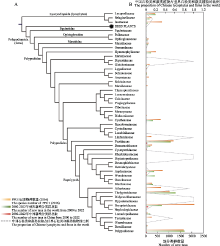
石松类和蕨类植物大系统.自1940年中国蕨类植物分类学创始人秦仁昌先生划时代地将传统水龙骨科重新划分以来, 世界石松类和蕨类植物分类系统的研究进入了快速发展阶段, 但很多关键类群的系统位置仍然存在较大争议(Smith et al, 2006; Christenhusz et al, 2011; PPG I, 2016).2017-2022年来, 中国学者通过系统发育基因组学研究对世界石松类和蕨类植物系统演化框架的重建做出了重要贡献.2018年, 中国两个蕨类研究团队几乎同时基于转录组测序的系统发育基因组学研究重建了世界石松类和蕨类科级的系统发育框架, 解决了数个过往长期存在争议的研究类群的问题, 如木贼目的系统位置、里白科与双扇蕨科的亲缘关系、碗蕨科与凤尾蕨科、水龙骨目的亲缘关系等(Qi et al, 2018; Shen et al, 2018).同时, 基于构建的世界石松类和蕨类植物的系统演化树, 重建了石松类和蕨类植物关键性状孢子囊环带的演化模式(Shen et al, 2018), 比较分析了石松类和蕨类植物转录因子家族的演化模式(Qi et al, 2018).然而, 基于质体基因组数据构建的石松类和蕨类植物系统发育关系与基于转录组数据的结果仍然存在较大差异, 特别是木贼目和膜蕨目等的系统位置, 这些差异可能与质体基因组数据集中存在系统发育冲突信号有关(Kuo et al, 2018).随着研究的深入, 石松类和蕨类植物科级水平的系统框架已基本确定, 然而由于前期研究中缺少多个国外科属的材料, 世界石松类和蕨类植物的完整系统框架仍需要进一步研究. ... Sex and the single gametophyte: Revising the homosporous vascular plant life cycle in light of contemporary research 1 2016 ... 传统的蕨类植物(pteridophytes)由两个不同的进化谱系, 即石松类(lycophytes)和蕨类植物(ferns)组成(Pryer et al, 2001; Smith et al, 2006; PPG I, 2016), 同时也是高等植物中配子体和孢子体都可以独立生活的植物类群(Haufler et al, 2016).根据化石记录, 石松类和蕨类植物最早可追溯到4亿年前, 是地球上起源最为古老的维管植物(Qi et al, 2018; Shen et al, 2018; 严岳鸿等, 2019; Huang et al, 2020), 同时也是维管植物中继被子植物之后的第二大类群.虽然被子植物在生态系统中占据了主导地位, 但蕨类植物也演化产生了适应多样化环境的形态和物种多样性, 在各生态系统中仍发挥着重要作用(Schneider et al, 2004; Ranker & Haufler, 2008; Huang et al, 2020; Du et al, 2021).中国幅员辽阔, 拥有复杂多样的地貌和气候条件, 是世界上石松类和蕨类植物最为丰富的地区之一.根据《中国生物物种名录》2022版(The Biodiversity Committee of Chinese Academy of Sciences, 2022)最新统计, 我国共有石松类3科12属165种4个种下分类群, 蕨类植物38科177属2,215种228个种下分类群, 为植物学家们提供了丰富的研究材料.自20世纪40年代秦仁昌先生发表水龙骨科自然分类系统, 中国石松类和蕨类植物研究就开始令世界瞩目.尤其是2017年第19届国际植物学大会在中国深圳召开后的5年时间里, 中国石松类和蕨类植物研究更是面向世界、走向国际, 研究更为广泛的科学问题, 在物种多样性、植物保护、系统演化和生态适应性等方面取得了一系列重要进展(Qi et al, 2018; Shen et al, 2018; Huang et al, 2020; Du et al, 2021; Wei R et al, 2021; Wang et al, 2022).本文对2017-2022年中国石松类和蕨类植物的物种多样性、保护、系统演化和生态适应性等研究进展进行综述, 以期为未来中国石松类和蕨类植物的研究提供参考. ... A molecular phylogeny of selligueoid ferns (Polypodiaceae): Implications for a natural delimitation despite homoplasy and rapid radiation 1 2018 ... 在科级水平上, 卷柏科、凤尾蕨科、碗蕨科、铁角蕨科、叉蕨科、水龙骨科等世界性大科的系统学研究方面取得了一系列重要研究进展.其中卷柏科和铁角蕨科主要基于形态和分子证据来解决科内属/组间的系统发育关系(Zhou et al, 2016, 2022; Xu KW et al, 2018; Xu et al, 2020; Zhang HR et al, 2020; Zhang MH et al, 2022)和厘清各疑难复合类群(如小卷柏复合群(Selaginella helvetica complex)、红枝卷柏复合群(S. sanguinolenta complex)、北京铁角蕨复合群(Asplenium pekinense complex)、变异铁角蕨复合群(A. varians complex)等)的系统分类地位(Liang et al, 2021; Zhang MH et al, 2021, 2022).同时, 卷柏科特殊质体结构, 如具有标志性同向重复序列(DR)的环形结构、GC含量高、碱基替换速率快等特点也常受到人们的广泛关注(Zhang MH et al, 2021, 2022; Xiang et al, 2022; Zhou et al, 2022).其他备受关注的类群, 如水龙骨科、凤尾蕨科、碗蕨科和叉蕨科等则着重通过扩大抽样来厘清科间或科下的系统关系和演化历史(Wei R et al, 2017; Zhang et al, 2017a, b; He et al, 2018; Zhang & Zhang, 2018a, b; Wei & Zhang, 2020; Zhao et al, 2020; Du et al, 2021; Zhou et al, 2022).如Wei R等(2021)通过在全球范围内对水龙骨科关键类群进行取样, 结合高通量测序技术获取了28属78个代表物种的叶绿体全基因组数据, 构建了迄今为止该科最为稳定的系统框架; 并通过根状茎鳞片形态、叶片分裂式样、叶脉类型、孢子囊群类型、隔丝形态以及孢子形态等形态学证据确定了各分支界线. ... 2 2020 ... 我国对蕨类配子体的研究及繁育最早可以追溯到20世纪80年代的哈尔滨师范大学包文美教授, 其先后对问荆(Equisetum arvense)、卷柏(Selaginella tamariscina)及水龙骨科、岩蕨科、铁角蕨科、紫萁科、(原)中国蕨科、(原)铁线蕨科等蕨类植物进行了繁育研究.随后, 哈尔滨师范大学的刘保东教授和上海师范大学的王全喜教授继续对多种中国产的蕨类植物进行了孢子繁育和系统的配子体发育研究(严岳鸿和石雷, 2014).近年来随着国家对生态文明建设的逐渐重视, 深圳市中国科学院仙湖植物园、中国科学院上海辰山植物园、中国科学院华南植物园、中国科学院西双版纳植物园、南京林业大学、西南林业大学等研究单位和高校都相继开展了石松类和蕨类植物的繁育工作, 其中孢子繁育研究主要集中在薄囊蕨类, 特别是外形美观、常用来做观赏植物开发的蕨类植物最多, 如凤尾蕨科、水龙骨科和铁角蕨科等, 其次是易于存活适合做园林搭配的蕨类植物, 如蹄盖蕨科、鳞毛蕨科、乌毛蕨科等.国家重点保护石松类和蕨类植物的孢子繁育近年来也取得了一定的研究成果(表5), 特别是桫椤科、水蕨属、鹿角蕨等植物, 这些类群植物的孢子繁育成活率已明显大大提高.另外, 福建观音座莲(Angiopteris fokiensis)的孢子萌发培养体系也在去年得到了新突破(何俊等, 2021), 填补了观音座莲属孢子萌发培养体系长期研究的空白, 为该属其他物种的保护、开发和利用提供了重要的理论基础和技术支持.与此同时, 还有一些演化较为原始或孑遗的石松类和蕨类植物, 如带状瓶尔小草、七指蕨、天星蕨、石杉属、马尾杉属等植物的孢子繁育瓶颈仍没有得到解决, 是后续需要重点关注的类群. ...
石松类和蕨类植物大系统.自1940年中国蕨类植物分类学创始人秦仁昌先生划时代地将传统水龙骨科重新划分以来, 世界石松类和蕨类植物分类系统的研究进入了快速发展阶段, 但很多关键类群的系统位置仍然存在较大争议(Smith et al, 2006; Christenhusz et al, 2011; PPG I, 2016).2017-2022年来, 中国学者通过系统发育基因组学研究对世界石松类和蕨类植物系统演化框架的重建做出了重要贡献.2018年, 中国两个蕨类研究团队几乎同时基于转录组测序的系统发育基因组学研究重建了世界石松类和蕨类科级的系统发育框架, 解决了数个过往长期存在争议的研究类群的问题, 如木贼目的系统位置、里白科与双扇蕨科的亲缘关系、碗蕨科与凤尾蕨科、水龙骨目的亲缘关系等(Qi et al, 2018; Shen et al, 2018).同时, 基于构建的世界石松类和蕨类植物的系统演化树, 重建了石松类和蕨类植物关键性状孢子囊环带的演化模式(Shen et al, 2018), 比较分析了石松类和蕨类植物转录因子家族的演化模式(Qi et al, 2018).然而, 基于质体基因组数据构建的石松类和蕨类植物系统发育关系与基于转录组数据的结果仍然存在较大差异, 特别是木贼目和膜蕨目等的系统位置, 这些差异可能与质体基因组数据集中存在系统发育冲突信号有关(Kuo et al, 2018).随着研究的深入, 石松类和蕨类植物科级水平的系统框架已基本确定, 然而由于前期研究中缺少多个国外科属的材料, 世界石松类和蕨类植物的完整系统框架仍需要进一步研究. ... 2 2020 ... 我国对蕨类配子体的研究及繁育最早可以追溯到20世纪80年代的哈尔滨师范大学包文美教授, 其先后对问荆(Equisetum arvense)、卷柏(Selaginella tamariscina)及水龙骨科、岩蕨科、铁角蕨科、紫萁科、(原)中国蕨科、(原)铁线蕨科等蕨类植物进行了繁育研究.随后, 哈尔滨师范大学的刘保东教授和上海师范大学的王全喜教授继续对多种中国产的蕨类植物进行了孢子繁育和系统的配子体发育研究(严岳鸿和石雷, 2014).近年来随着国家对生态文明建设的逐渐重视, 深圳市中国科学院仙湖植物园、中国科学院上海辰山植物园、中国科学院华南植物园、中国科学院西双版纳植物园、南京林业大学、西南林业大学等研究单位和高校都相继开展了石松类和蕨类植物的繁育工作, 其中孢子繁育研究主要集中在薄囊蕨类, 特别是外形美观、常用来做观赏植物开发的蕨类植物最多, 如凤尾蕨科、水龙骨科和铁角蕨科等, 其次是易于存活适合做园林搭配的蕨类植物, 如蹄盖蕨科、鳞毛蕨科、乌毛蕨科等.国家重点保护石松类和蕨类植物的孢子繁育近年来也取得了一定的研究成果(表5), 特别是桫椤科、水蕨属、鹿角蕨等植物, 这些类群植物的孢子繁育成活率已明显大大提高.另外, 福建观音座莲(Angiopteris fokiensis)的孢子萌发培养体系也在去年得到了新突破(何俊等, 2021), 填补了观音座莲属孢子萌发培养体系长期研究的空白, 为该属其他物种的保护、开发和利用提供了重要的理论基础和技术支持.与此同时, 还有一些演化较为原始或孑遗的石松类和蕨类植物, 如带状瓶尔小草、七指蕨、天星蕨、石杉属、马尾杉属等植物的孢子繁育瓶颈仍没有得到解决, 是后续需要重点关注的类群. ...
本文的其它图/表
|