中国石松类和蕨类植物多样性研究进展
王婷, 舒江平, 顾钰峰, 李艳清, 杨拓, 徐洲锋, 向建英, 张宪春, 严岳鸿
生物多样性
2022, 30 ( 7):
22381-.
DOI: 10.17520/biods.2022381
石松类和蕨类植物是维管植物的第二大类群, 其起源可追溯到4亿年前。在被子植物出现之前, 石松类和蕨类植物在古地球生态系统中占主导地位, 其重要性一直延续到现在。自20世纪40年代开始, 中国石松类和蕨类植物研究就令世界瞩目, 尤其是2017年第19届国际植物学大会在中国深圳召开后的5年时间里, 中国石松类和蕨类植物研究更是面向世界、走向国际, 研究更为广泛的科学问题, 在物种多样性、保护、系统演化和生态适应性等方面取得了一系列重要研究进展。2017-2022年, 多个中国研究团队利用多组学数据构建了世界石松类和蕨类植物科级水平的生命之树并提出了关键性状孢子囊环带演化的新模式; 解决了石松类和蕨类植物中目级、科级、属级和种级众多关键的系统分类学等问题, 发表了106个新分类群; 开展了大量的植物区系调查和研究, 出版了6部中国石松类和蕨类植物多样性专著和1部世界性专著; 对65种国家重点保护的石松类和蕨类植物进行了迁地保护, 同时实现了桫椤科、水蕨属(Ceratopteris)、观音座莲属(Angiopteris)和鹿角蕨(Platycerium wallichii)等重点保护类群的孢子繁殖; 在系统发育框架下, 研究了石松类和蕨类植物的生态修复功能和生态适应性演化。通过对2017-2022年研究成果的总结和思考, 本文对未来石松类和蕨类植物的发展提出以下建议: (1)提高中国寡型科属以及世界性大科大属的关注力度; (2)加强西藏、四川等薄弱地区石松类和蕨类植物的调查研究, 并结合新技术, 如DNA条形码等以提高区系调查中物种鉴别的效率和准确性; (3)运用多学科交叉的研究方法厘清各科、属、种间系统关系的同时, 还应加强系统和生态适应性演化之间的协同研究; (4)关注石松类和蕨类植物系统位置作为陆生维管植物演化起点的共性科学问题; (5)加强石松类和蕨类植物系统分类学与生态学、植物化学、保护生物学等学科间交叉合作研究。
省级行政区划
Provincial administrative area | 2017-2022年 | 2000-2022年 | 新种
New
species | 新种下分类群 New infraspecific taxon | 新记录
New
record | 采集密度
Sampling density | 新种
New
species | 新种下分类群 New infraspecific taxon | 新记录
New
record | 采集密度
Sampling density | | 安徽 | Anhui | - | - | 2 | 264 | 2 | - | 3 | 267 | | 澳门 | Aomen | - | - | - | - | - | - | - | - | | 北京 | Beijing | - | - | - | 31 | - | 1 | - | 52 | | 重庆 | Chongqing | 1 | - | - | 117 | 6 | 1 | - | 142 | | 福建 | Fujian | 1 | - | - | 109 | 3 | 1 | 1 | 174 | | 甘肃 | Gansu | - | - | - | 101 | - | - | 1 | 138 | | 广东 | Guangdong | 9 | - | - | 1,943 | 16 | 1 | - | 3,764 | | 广西 | Guangxi | 16 | - | - | 124 | 29 | - | 3 | 646 | | 贵州 | Guizhou | 20 | - | - | 786 | 59 | 7 | 2 | 912 | | 海南 | Hainan | 7 | - | - | 108 | 16 | - | 2 | 535 | | 河北 | Hebei | - | - | - | 14 | - | - | - | 21 | | 黑龙江 | Heilongjiang | 1 | - | - | - | 1 | - | 1 | - | | 河南 | Henan | - | - | - | 26 | - | - | - | 28 | | 香港 | Hong Kong | 1 | - | - | - | 2 | 2 | - | - | | 湖北 | Hubei | - | - | - | 2,407 | 4 | 3 | - | 3,218 | | 湖南 | Hunan | 4 | - | - | 102 | 8 | - | 1 | 719 | | 江苏 | Jiangsu | - | - | - | 69 | - | - | - | 104 | | 江西 | Jiangxi | 1 | - | - | 564 | 3 | 1 | 1 | 1,329 | | 吉林 | Jilin | - | - | - | 2 | - | - | - | 8 | | 辽宁 | Liaoning | - | - | - | - | - | - | - | - | | 内蒙古 | Nei Mongol | - | - | - | - | 1 | - | 1 | - | | 宁夏 | Ningxia | - | - | - | 14 | - | - | - | - | | 青海 | Qinghai | - | - | - | 9 | - | - | - | 11 | | 陕西 | Shaanxi | - | - | - | 92 | 3 | - | 1 | 95 | | 山东 | Shandong | 3 | - | - | 56 | 5 | - | - | 77 | | 上海 | Shanghai | - | - | - | 1 | - | - | | 8 | | 山西 | Shanxi | - | - | - | 37 | 1 | - | 1 | - | | 四川 | Sichuan | 5 | - | - | 175 | 16 | 2 | 3 | 837 | | 台湾 | Taiwan | 13 | 1 | 6 | 2 | 25 | 5 | 10 | 7 | | 天津 | Tianjin | - | - | - | 1 | - | - | - | - | | 新疆 | Xinjiang | - | - | - | 15 | - | - | 2 | 44 | | 西藏 | Xizang | 2 | - | - | 211 | 31 | 5 | 3 | 999 | | 云南 | Yunnan | 33 | - | 1 | 2,424 | 71 | 17 | 11 | 6,826 | | 浙江 | Zhejiang | 1 | - | - | 685 | 5 | - | - | 724 |
View table in article
表3
中国各省级行政区划新分类群发现的数量和采集空间分布情况
正文中引用本图/表的段落
以2017-2022年和2000-2022年为时间界线, 对中国石松类和蕨类植物新物种、新种下分类群和中国新记录所属省份进行统计。结果显示(表3), 我国在这2个时间段发表的石松类和蕨类植物新物种、新种下分类群和国家新记录主要集中分布在某些热点或优先保护区域, 如云南、贵州、广西等地。其中, 云南新类群分布最多, 2000-2022年和2017-2022年共计发表了71和33个新物种、17和0个种下分类群、11和1个中国新记录, 占中国近22年来新发表物种数量的1/4, 是国内外学者广泛关注的区域; 其次为贵州, 2000-2022年和2017-2022年共计发表了59和20个新物种、7和0个新种下分类群、2和0个中国新记录, 占中国近22年来新发表物种数量的1/6。然而, 同样位于生物多样性热点区域的西藏和四川, 近年来在被子植物新发现上存在较高的密度(杜诚等, 2021), 但在石松类和蕨类植物类群上并没有体现, 推测与这些区域石松类和蕨类植物专类调查不充分、研究人员较少具有一定的关系。
Biotracks软件是2016年9月对外上线的一款自然观察类的公众科学应用, 截至目前(2022年7月26日)已有35,953位自然观察者, 累积2,314,943个坐标点, 已被各类科学调查和自然观察项目广泛使用(徐洲锋等, 2021), 在中国采集记录中具有一定参考性。为了解我国石松类和蕨类植物标本的采集状况, 预测可能的采集空缺, 我们调取和整理了Biotracks (http://www.biotracks.cn/) 2017-2022年和Global Biodiversity Information Facility (GBIF; https://www.gbif.org/ ) 2000-2022年关于中国石松类和蕨类植物的采集标本点, 并对各省份进行采集分布空间统计和提取(表3)。2个时间段的采集分布空间结果均显示调查密度总体上与新分类群、新记录发现的数量成正比(表3)。2017-2022年期间, Biotracks分布空间数据显示云南、广东、贵州、大别山区、黄山-怀玉山区、大巴山区、岷山-横断山北段区等地具有较高的采集分布空间, 生物多样性薄弱的西北、华北等地采集密度普遍较低; 而西藏、四川等存在部分采集空缺区域。
植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014)。2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3)。从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道。针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结。2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等。尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3)。
英国生态学家诺曼•麦尔在1988年提出了生物多样性热点地区(biodiversity hotspots)的概念, 他认识到这些热点生态系统在很小的地域面积内包含了极其丰富的物种多样性。2000年, 国际环保组织根据特有物种数量和受威胁程度两个评定标准确定了全球34个物种最丰富且受到威胁最大的生物多样性热点地区(Myers et al, 2000), 其中涉及中国的区域有3个, 即中国西南山区(Mountains of South-West China)、华南-中南半岛地区(Indo-Burma)和喜马拉雅地区(Himalayan)。2015年, 为贯彻落实国务院批准发布的《中国生物多样性保护战略与行动计划(2011-2030年)》, 国家环境保护部组织开展了生物多样性保护优先区域边界的核定工作, 确定了35个中国生物多样性保护优先区域范围。结合2017-2022年发表的中国石松类和蕨类植物新物种、新种下分类群和国家新记录所在的省份和调查分布空间情况(表3), 可清晰发现西藏、四川为近年来石松类和蕨类植物调查薄弱的生物多样性热点区域。提高野外调查密度、加强学科队伍建设在一定程度上能促进生物多样性热点地区石松类和蕨类植物物种多样性的发现(表3)。中国幅员辽阔, 气候地貌复杂多样, 物种丰富, 开展中国蕨类植物多样性热点区域本底调查和系统分类等工作仍任重道远。
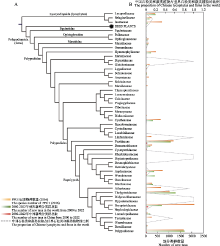
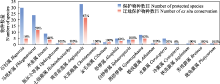
石松类和蕨类植物大系统.自1940年中国蕨类植物分类学创始人秦仁昌先生划时代地将传统水龙骨科重新划分以来, 世界石松类和蕨类植物分类系统的研究进入了快速发展阶段, 但很多关键类群的系统位置仍然存在较大争议(Smith et al, 2006; Christenhusz et al, 2011; PPG I, 2016).2017-2022年来, 中国学者通过系统发育基因组学研究对世界石松类和蕨类植物系统演化框架的重建做出了重要贡献.2018年, 中国两个蕨类研究团队几乎同时基于转录组测序的系统发育基因组学研究重建了世界石松类和蕨类科级的系统发育框架, 解决了数个过往长期存在争议的研究类群的问题, 如木贼目的系统位置、里白科与双扇蕨科的亲缘关系、碗蕨科与凤尾蕨科、水龙骨目的亲缘关系等(Qi et al, 2018; Shen et al, 2018).同时, 基于构建的世界石松类和蕨类植物的系统演化树, 重建了石松类和蕨类植物关键性状孢子囊环带的演化模式(Shen et al, 2018), 比较分析了石松类和蕨类植物转录因子家族的演化模式(Qi et al, 2018).然而, 基于质体基因组数据构建的石松类和蕨类植物系统发育关系与基于转录组数据的结果仍然存在较大差异, 特别是木贼目和膜蕨目等的系统位置, 这些差异可能与质体基因组数据集中存在系统发育冲突信号有关(Kuo et al, 2018).随着研究的深入, 石松类和蕨类植物科级水平的系统框架已基本确定, 然而由于前期研究中缺少多个国外科属的材料, 世界石松类和蕨类植物的完整系统框架仍需要进一步研究. ... Phytochemicals from fern species:Potential for medicine applications 1 2017 ... 石松类和蕨类植物是国家重点保护野生植物名录中包含物种数量最多的类群之一, 而且绝大多数石松类和蕨类植物都具有重要的药用和观赏价值(Cao et al, 2017; Xu ZC et al, 2018; Abraham & Thomas, 2022).近年来, 中国学者开展了少数重点保护的珍稀濒危蕨类植物, 如笔筒树(Sphaeropteris lepifera)、荷叶铁线蕨等相关保护生物学研究(Wei XY et al, 2021; 孙维悦等, 2022), 但绝大多数濒危物种都没有开展相应保护生物学研究, 对于珍稀濒危的石松类和蕨类植物的种群数量和生存现状都不甚清楚.这一方面是由于石松类和蕨类植物缺少类似于兰科植物的全国性大调查而且类群的社会关注度较小, 另一方面是石松类和蕨类植物的研究人员, 特别是青年科研人员较少, 研究力量严重不足.今后应加大专业科研人员的培养, 组织开展石松类和蕨类植物的全国性大普查, 从而进一步完善石松类和蕨类植物名录和濒危等级评估, 为今后《国家重点保护野生植物名录》的修订奠定基础.同时, 积极开展石松类和蕨类植物的孢子繁殖研究, 实现药用、观赏和珍稀濒危植物的大规模繁殖, 从而对重要经济价值的类群实现产业化应用, 而对重点保护野生植物开展迁地保护和野外回归, 进一步提升石松类和蕨类植物的社会关注度. ... 浙江省景宁县望东垟、大仰湖自然保护区蕨类植物区系分析 1 2020 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 浙江省景宁县望东垟、大仰湖自然保护区蕨类植物区系分析 1 2020 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 浙江建德蕨类植物区系研究 1 2019 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 浙江建德蕨类植物区系研究 1 2019 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... Plastid phylogenomics and plastomic diversity of the extant lycophytes 1 2022 ... 重要类群的系统演化.以往研究仅基于少数叶绿体基因片段和形态学数据, 很难解决石松类和蕨类植物中疑难类群的系统演化问题, 如铁角蕨科、水龙骨科、卷柏科、碗蕨科等.近年来, 随着测序技术和计算科学的不断发展, 利用质体基因组或转录组测序数据来研究中国石松类和蕨类植物的系统分类与演化呈现爆发式增长的情况(杜新宇等, 2019), 近5年国内学者在石松类和蕨类植物的重要类群中也取得了大量进展.在目级水平上, Shu等(2022b)利用转录组数据重建了早期薄囊蕨类的系统演化关系, 确立了马通蕨目的分类地位,①(①夏漪 (2014) 三种观赏蕨类植物配子体发育及卵发生的研究. 硕士学位论文, 上海师范大学, 上海.)该目包括双扇蕨科和马通蕨科, 同时揭示了古多倍化和古杂交事件对于马通蕨目植物在中生代时期辐射演化中起到了重要作用.Du XY等(2021)基于水龙骨目27科100属160种的质体基因组序列, 利用系统发育基因组学方法和多种分析策略解析了水龙骨目深层的系统关系, 同时揭示水龙骨目和被子植物同期多样化的模式, 填补了对陆地植被革命如何塑造现今生态系统的部分认知的空白; Chen等(2022)基于81个质体基因组序列解析了世界石松类的系统发育关系和质体基因组结构变异. ... 药用植物千层塔的基原物种及研究进展 1 2021 ... 新版名录的一个重要特征是在科级和属级水平对一些重要的类群进行了所有种的保护.在科属级别进行全部保护的石松类和蕨类植物有7个类群, 包括石杉属、马尾杉属、水韭属(Isoetes)、观音座莲属、金毛狗属(Cibotium)、水蕨属(Ceratopteris)和桫椤科(除小黑桫椤和粗齿桫椤).一方面, 因为这些类群中的绝大多数物种为经济濒危种(严岳鸿等, 2006), 具有重要的药用、食用和观赏价值, 很容易受到人为采挖破坏.例如, 石杉属和马尾杉属植物广布于热带和亚热带地区, 在我国主产西南和华南, 东北、西北及华东也有分布.由于含有治疗阿尔兹海默症的有效成分——石杉碱甲(Ma et al, 2006, 2007), 而被作为重要的传统药材广泛利用(李国树和徐成东, 2009; 赵刚等, 2018).而且, 由于石杉碱甲的化学合成步骤繁杂、条件苛刻、成本高、收率低等问题, 未能通过人工化学合成而实现量产(Ferreira et al, 2016), 目前仍然主要从植物中提取, 因此对野生石杉属和马尾杉属植物消耗巨大(马小军等, 2009; 赵刚等, 2018; 谢峻等, 2020).然而, 石杉属和马尾杉属植物的植株生长缓慢、孢子萌发时间长、种群更新能力弱(鞠錾, 2009①( 鞠錾 (2009) 四种石杉科植物中产生石杉碱甲的内生真菌的分离、鉴定和其石杉碱甲含量的测定. 硕士学位论文, 复旦大学, 上海.); 齐耀东和王德立, 2017; 肖友利等, 2020), 其中石杉属植物完成一个完整的生活史约需15-20年之久(齐耀东和王德立, 2017), 一旦植物遭到破坏就很难再恢复(陈思思等, 2021).另一方面, 这些类群中大多数物种为演化濒危种, 即处于演化系统基部类群的孑遗物种(严岳鸿等, 2006).它们在地理分布区上表现出孑遗性、间断性、边缘性或孤立性; 环境变化或人为活动的长期干扰可能导致其分布区正逐渐缩小, 如桫椤科、莲座蕨属等.另外, 这些类群还存在较强的物种相似性, 即与同属的其他物种容易混淆(鲁兆莉等, 2021), 在整个属级水平进行保护才有可能实现对濒危野生蕨类植物的有效保护, 具有更好的可操作性. ... 药用植物千层塔的基原物种及研究进展 1 2021 ... 新版名录的一个重要特征是在科级和属级水平对一些重要的类群进行了所有种的保护.在科属级别进行全部保护的石松类和蕨类植物有7个类群, 包括石杉属、马尾杉属、水韭属(Isoetes)、观音座莲属、金毛狗属(Cibotium)、水蕨属(Ceratopteris)和桫椤科(除小黑桫椤和粗齿桫椤).一方面, 因为这些类群中的绝大多数物种为经济濒危种(严岳鸿等, 2006), 具有重要的药用、食用和观赏价值, 很容易受到人为采挖破坏.例如, 石杉属和马尾杉属植物广布于热带和亚热带地区, 在我国主产西南和华南, 东北、西北及华东也有分布.由于含有治疗阿尔兹海默症的有效成分——石杉碱甲(Ma et al, 2006, 2007), 而被作为重要的传统药材广泛利用(李国树和徐成东, 2009; 赵刚等, 2018).而且, 由于石杉碱甲的化学合成步骤繁杂、条件苛刻、成本高、收率低等问题, 未能通过人工化学合成而实现量产(Ferreira et al, 2016), 目前仍然主要从植物中提取, 因此对野生石杉属和马尾杉属植物消耗巨大(马小军等, 2009; 赵刚等, 2018; 谢峻等, 2020).然而, 石杉属和马尾杉属植物的植株生长缓慢、孢子萌发时间长、种群更新能力弱(鞠錾, 2009①( 鞠錾 (2009) 四种石杉科植物中产生石杉碱甲的内生真菌的分离、鉴定和其石杉碱甲含量的测定. 硕士学位论文, 复旦大学, 上海.); 齐耀东和王德立, 2017; 肖友利等, 2020), 其中石杉属植物完成一个完整的生活史约需15-20年之久(齐耀东和王德立, 2017), 一旦植物遭到破坏就很难再恢复(陈思思等, 2021).另一方面, 这些类群中大多数物种为演化濒危种, 即处于演化系统基部类群的孑遗物种(严岳鸿等, 2006).它们在地理分布区上表现出孑遗性、间断性、边缘性或孤立性; 环境变化或人为活动的长期干扰可能导致其分布区正逐渐缩小, 如桫椤科、莲座蕨属等.另外, 这些类群还存在较强的物种相似性, 即与同属的其他物种容易混淆(鲁兆莉等, 2021), 在整个属级水平进行保护才有可能实现对濒危野生蕨类植物的有效保护, 具有更好的可操作性. ... 水蕨配子体和孢子体发育的研究 1 2008 ... Spore reproduction success of lycophytes and ferns under state priority conservation
石松类和蕨类植物大系统.自1940年中国蕨类植物分类学创始人秦仁昌先生划时代地将传统水龙骨科重新划分以来, 世界石松类和蕨类植物分类系统的研究进入了快速发展阶段, 但很多关键类群的系统位置仍然存在较大争议(Smith et al, 2006; Christenhusz et al, 2011; PPG I, 2016).2017-2022年来, 中国学者通过系统发育基因组学研究对世界石松类和蕨类植物系统演化框架的重建做出了重要贡献.2018年, 中国两个蕨类研究团队几乎同时基于转录组测序的系统发育基因组学研究重建了世界石松类和蕨类科级的系统发育框架, 解决了数个过往长期存在争议的研究类群的问题, 如木贼目的系统位置、里白科与双扇蕨科的亲缘关系、碗蕨科与凤尾蕨科、水龙骨目的亲缘关系等(Qi et al, 2018; Shen et al, 2018).同时, 基于构建的世界石松类和蕨类植物的系统演化树, 重建了石松类和蕨类植物关键性状孢子囊环带的演化模式(Shen et al, 2018), 比较分析了石松类和蕨类植物转录因子家族的演化模式(Qi et al, 2018).然而, 基于质体基因组数据构建的石松类和蕨类植物系统发育关系与基于转录组数据的结果仍然存在较大差异, 特别是木贼目和膜蕨目等的系统位置, 这些差异可能与质体基因组数据集中存在系统发育冲突信号有关(Kuo et al, 2018).随着研究的深入, 石松类和蕨类植物科级水平的系统框架已基本确定, 然而由于前期研究中缺少多个国外科属的材料, 世界石松类和蕨类植物的完整系统框架仍需要进一步研究. ... 南雄小流坑--青嶂山保护区维管植物区系研究 1 2017 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 南雄小流坑--青嶂山保护区维管植物区系研究 1 2017 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... A linear sequence of extant families and genera of lycophytes and ferns 1 2011 ... 石松类和蕨类植物大系统.自1940年中国蕨类植物分类学创始人秦仁昌先生划时代地将传统水龙骨科重新划分以来, 世界石松类和蕨类植物分类系统的研究进入了快速发展阶段, 但很多关键类群的系统位置仍然存在较大争议(Smith et al, 2006; Christenhusz et al, 2011; PPG I, 2016).2017-2022年来, 中国学者通过系统发育基因组学研究对世界石松类和蕨类植物系统演化框架的重建做出了重要贡献.2018年, 中国两个蕨类研究团队几乎同时基于转录组测序的系统发育基因组学研究重建了世界石松类和蕨类科级的系统发育框架, 解决了数个过往长期存在争议的研究类群的问题, 如木贼目的系统位置、里白科与双扇蕨科的亲缘关系、碗蕨科与凤尾蕨科、水龙骨目的亲缘关系等(Qi et al, 2018; Shen et al, 2018).同时, 基于构建的世界石松类和蕨类植物的系统演化树, 重建了石松类和蕨类植物关键性状孢子囊环带的演化模式(Shen et al, 2018), 比较分析了石松类和蕨类植物转录因子家族的演化模式(Qi et al, 2018).然而, 基于质体基因组数据构建的石松类和蕨类植物系统发育关系与基于转录组数据的结果仍然存在较大差异, 特别是木贼目和膜蕨目等的系统位置, 这些差异可能与质体基因组数据集中存在系统发育冲突信号有关(Kuo et al, 2018).随着研究的深入, 石松类和蕨类植物科级水平的系统框架已基本确定, 然而由于前期研究中缺少多个国外科属的材料, 世界石松类和蕨类植物的完整系统框架仍需要进一步研究. ... 贵州高原湿地草海水生植物的区系特征与地理分布 1 2022 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 贵州高原湿地草海水生植物的区系特征与地理分布 1 2022 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 水蕨配子体发育的研究 1 2005 ... Spore reproduction success of lycophytes and ferns under state priority conservation
石松类和蕨类植物大系统.自1940年中国蕨类植物分类学创始人秦仁昌先生划时代地将传统水龙骨科重新划分以来, 世界石松类和蕨类植物分类系统的研究进入了快速发展阶段, 但很多关键类群的系统位置仍然存在较大争议(Smith et al, 2006; Christenhusz et al, 2011; PPG I, 2016).2017-2022年来, 中国学者通过系统发育基因组学研究对世界石松类和蕨类植物系统演化框架的重建做出了重要贡献.2018年, 中国两个蕨类研究团队几乎同时基于转录组测序的系统发育基因组学研究重建了世界石松类和蕨类科级的系统发育框架, 解决了数个过往长期存在争议的研究类群的问题, 如木贼目的系统位置、里白科与双扇蕨科的亲缘关系、碗蕨科与凤尾蕨科、水龙骨目的亲缘关系等(Qi et al, 2018; Shen et al, 2018).同时, 基于构建的世界石松类和蕨类植物的系统演化树, 重建了石松类和蕨类植物关键性状孢子囊环带的演化模式(Shen et al, 2018), 比较分析了石松类和蕨类植物转录因子家族的演化模式(Qi et al, 2018).然而, 基于质体基因组数据构建的石松类和蕨类植物系统发育关系与基于转录组数据的结果仍然存在较大差异, 特别是木贼目和膜蕨目等的系统位置, 这些差异可能与质体基因组数据集中存在系统发育冲突信号有关(Kuo et al, 2018).随着研究的深入, 石松类和蕨类植物科级水平的系统框架已基本确定, 然而由于前期研究中缺少多个国外科属的材料, 世界石松类和蕨类植物的完整系统框架仍需要进一步研究. ... Taxonomic, phylogenetic, and functional diversity of ferns at three differently disturbed sites in Longnan County, China 1 2020 ... 石松类和蕨类植物广布于世界各地, 多以草本形式生长在林下、林缘或林窗中, 是森林演替早期阶段的先锋植物.蕨类植物对环境的适应和生态学意义一直受到研究人员的关注.但近年来, 我国对蕨类植物的生态学研究进展较少, 主要集中在蕨类植物的生态修复和生态适应上.在生态修复方面, Yang等(2022)研究发现, 先锋蕨类植物芒萁属能在降雨过后通过有性繁殖和克隆繁殖两种方式快速覆盖受损裸地, 形成密集蕨类灌丛, 在忍耐地表高温、贫瘠土壤、强烈光照的同时, 还有利于改善微环境, 促进土壤养分积累和抵御外来种入侵; Yan等(2019)通过三代全长转录组、二代差异转录组和液泡膜蛋白质组的方法揭示了蜈蚣草(Pteris vittata)的砷超富集机理及其调控分子网络, 对利用植物修复治理砷污染土壤具有重要意义.在生态适应上, Jin等(2021)通过测量345种蕨类植物对环境光和水可利用性敏感的功能性状发现, 鳞毛蕨类和水龙骨类的部分物种可以耐受强光和干旱的生境, 这些类群可能是蕨类植物适应干旱和明亮生境的先驱; Liang等(2022)以亚洲温带广布的二倍体同型孢子蕨类——中华蹄盖蕨复合群(Athyrium sinense complex)为研究对象, 通过群体遗传分析和分布区模拟等揭示了蕨类植物孢子强扩散能力下生态适应塑造的遗传分化格局.Dai等(2020)通过比较不同干扰水平下蕨类植物的分类、系统发育和功能多样性, 发现蕨类植物的多样性在很大程度上受到人类干扰水平的影响, 可作为环境胁迫的一个良好指示物种. ... 井冈山大学绶草群落植物区系与物种多样性研究 1 2020 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 井冈山大学绶草群落植物区系与物种多样性研究 1 2020 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 黔东典型湿地公园高等植物多样性及区系分析 1 2020 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 黔东典型湿地公园高等植物多样性及区系分析 1 2020 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 热带水库消落区植物群落物种组成与结构特征--以海南松涛水库为例 1 2018 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 热带水库消落区植物群落物种组成与结构特征--以海南松涛水库为例 1 2018 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 海南岛蕨类植物的分类、区系地理与保育 1 2004 ... 物种的濒危等级是制定国家重点保护野生植物名录的重要参考依据.世界自然保护联盟(International Union for Conservation of Nature, IUCN)濒危物种红色名录等级和标准是目前国内外学者普遍采用的一种相对客观的评价标准.近年来, 中国学者根据IUCN等级和标准初步对中国石松类和蕨类植物的濒危等级进行了评估(董仕勇, 2004; 严岳鸿等, 2014; 董仕勇等, 2017), 但由于资料缺乏, 很多物种难以准确评估.2013年, 严岳鸿等对中国2,456种蕨类植物进行IUCN等级评估, 结果显示中国的蕨类植物中, 属于灭绝(EX)的有埃及蘋(Marsilea aegyptiaca)、光叶蕨(Cystoathyrium chinense) 2种, 属于极危(CR)的有荷叶铁线蕨(Adiantum nelumboides)等33种, 属于濒危(EN)的有对开蕨(Asplenium komarovii)等51种, 属于易危(VU)的有雨蕨(Gymnogrammitis dareiformis)等109种, 属于近危(NT)的有阔片乌蕨(Sphenomeris biflora)等158种, 属于无危(LC)的有狗脊(Woodwardia japonica)等845种, 不宜评估(NE)的有细叶满江红(Azolla filiculoides)等3种, 数据缺乏(DD)的有1,255种(严岳鸿等, 2014).2017年, 董仕勇等对中国2,244种(包括亚种和变种)蕨类植物进行IUCN红色名录等级评估, 结果显示极危物种有43种(其中6种可能已经灭绝)、濒危物种有68种、易危物种有71种、近危物种有66种、无危物种1,124种、数据缺乏的物种有872种(董仕勇等, 2017). ... 海南岛蕨类植物的分类、区系地理与保育 1 2004 ... 物种的濒危等级是制定国家重点保护野生植物名录的重要参考依据.世界自然保护联盟(International Union for Conservation of Nature, IUCN)濒危物种红色名录等级和标准是目前国内外学者普遍采用的一种相对客观的评价标准.近年来, 中国学者根据IUCN等级和标准初步对中国石松类和蕨类植物的濒危等级进行了评估(董仕勇, 2004; 严岳鸿等, 2014; 董仕勇等, 2017), 但由于资料缺乏, 很多物种难以准确评估.2013年, 严岳鸿等对中国2,456种蕨类植物进行IUCN等级评估, 结果显示中国的蕨类植物中, 属于灭绝(EX)的有埃及蘋(Marsilea aegyptiaca)、光叶蕨(Cystoathyrium chinense) 2种, 属于极危(CR)的有荷叶铁线蕨(Adiantum nelumboides)等33种, 属于濒危(EN)的有对开蕨(Asplenium komarovii)等51种, 属于易危(VU)的有雨蕨(Gymnogrammitis dareiformis)等109种, 属于近危(NT)的有阔片乌蕨(Sphenomeris biflora)等158种, 属于无危(LC)的有狗脊(Woodwardia japonica)等845种, 不宜评估(NE)的有细叶满江红(Azolla filiculoides)等3种, 数据缺乏(DD)的有1,255种(严岳鸿等, 2014).2017年, 董仕勇等对中国2,244种(包括亚种和变种)蕨类植物进行IUCN红色名录等级评估, 结果显示极危物种有43种(其中6种可能已经灭绝)、濒危物种有68种、易危物种有71种、近危物种有66种、无危物种1,124种、数据缺乏的物种有872种(董仕勇等, 2017). ... 2 2022 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ...
石松类和蕨类植物大系统.自1940年中国蕨类植物分类学创始人秦仁昌先生划时代地将传统水龙骨科重新划分以来, 世界石松类和蕨类植物分类系统的研究进入了快速发展阶段, 但很多关键类群的系统位置仍然存在较大争议(Smith et al, 2006; Christenhusz et al, 2011; PPG I, 2016).2017-2022年来, 中国学者通过系统发育基因组学研究对世界石松类和蕨类植物系统演化框架的重建做出了重要贡献.2018年, 中国两个蕨类研究团队几乎同时基于转录组测序的系统发育基因组学研究重建了世界石松类和蕨类科级的系统发育框架, 解决了数个过往长期存在争议的研究类群的问题, 如木贼目的系统位置、里白科与双扇蕨科的亲缘关系、碗蕨科与凤尾蕨科、水龙骨目的亲缘关系等(Qi et al, 2018; Shen et al, 2018).同时, 基于构建的世界石松类和蕨类植物的系统演化树, 重建了石松类和蕨类植物关键性状孢子囊环带的演化模式(Shen et al, 2018), 比较分析了石松类和蕨类植物转录因子家族的演化模式(Qi et al, 2018).然而, 基于质体基因组数据构建的石松类和蕨类植物系统发育关系与基于转录组数据的结果仍然存在较大差异, 特别是木贼目和膜蕨目等的系统位置, 这些差异可能与质体基因组数据集中存在系统发育冲突信号有关(Kuo et al, 2018).随着研究的深入, 石松类和蕨类植物科级水平的系统框架已基本确定, 然而由于前期研究中缺少多个国外科属的材料, 世界石松类和蕨类植物的完整系统框架仍需要进一步研究. ... 1 2015 ... 中国是北半球植物种类最丰富的国家, 目前已记载超过32,500种维管植物(Hong & Blackmore, 2015), 同时也是近年来发现和命名维管植物新物种最多的3个国家之一(RBG Kew, 2016).从中国石松类和蕨类植物新类群的发现数量、比例以及参与新分类群命名学者的人数来看(表2, 附录5-6), 中国物种数量较少的寡型科属类群的物种多样性常常被忽视, 例如木贼科、乌毛蕨科、瓶尔小草科、合囊蕨科、桫椤科、藤蕨科、条蕨科、骨碎补科、紫萁科; 这些类群实际新增数目均低于期望新增数目.在关注程度上, 除合囊蕨科外, 其余类群的实际关注人次均小于期望关注人数, 反映了过往研究学者们对于寡型科属植物关注力度较低的情况.近年来, 部分研究学者在常见寡型科属植物中发现大量新物种, 例如近期关于芒萁属隐性物种的文章(Wei et al, 2022), 作者对亚洲芒萁属植物进行广泛采样和系统研究, 恢复了芒萁属4个变种(D. inaequalis、D. alternans、D. subspeciosa和D. latiloba)的物种地位, 同时描述和命名了2个新物种(D. austrosinensis、D. baliensis).通过世界新增比例预测, 中国寡型科属可能存在较多的物种多样性, 需在日后受到更多研究学者的关注. ... 贵阳阿哈湖国家湿地公园石松类和蕨类植物初步研究 1 2020 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 贵阳阿哈湖国家湿地公园石松类和蕨类植物初步研究 1 2020 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 江西共青城南湖湿地自然保护区维管植物区系分析 1 2020 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 江西共青城南湖湿地自然保护区维管植物区系分析 1 2020 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 茂兰国家级自然保护区石松类和蕨类植物区系特征 1 2021 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 茂兰国家级自然保护区石松类和蕨类植物区系特征 1 2021 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 莲都峰源自然保护区植物区系和地理成分 1 2020 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 莲都峰源自然保护区植物区系和地理成分 1 2020 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... Recurrent genome duplication events likely contributed to both the ancient and recent rise of ferns 4 2020 ... 传统的蕨类植物(pteridophytes)由两个不同的进化谱系, 即石松类(lycophytes)和蕨类植物(ferns)组成(Pryer et al, 2001; Smith et al, 2006; PPG I, 2016), 同时也是高等植物中配子体和孢子体都可以独立生活的植物类群(Haufler et al, 2016).根据化石记录, 石松类和蕨类植物最早可追溯到4亿年前, 是地球上起源最为古老的维管植物(Qi et al, 2018; Shen et al, 2018; 严岳鸿等, 2019; Huang et al, 2020), 同时也是维管植物中继被子植物之后的第二大类群.虽然被子植物在生态系统中占据了主导地位, 但蕨类植物也演化产生了适应多样化环境的形态和物种多样性, 在各生态系统中仍发挥着重要作用(Schneider et al, 2004; Ranker & Haufler, 2008; Huang et al, 2020; Du et al, 2021).中国幅员辽阔, 拥有复杂多样的地貌和气候条件, 是世界上石松类和蕨类植物最为丰富的地区之一.根据《中国生物物种名录》2022版(The Biodiversity Committee of Chinese Academy of Sciences, 2022)最新统计, 我国共有石松类3科12属165种4个种下分类群, 蕨类植物38科177属2,215种228个种下分类群, 为植物学家们提供了丰富的研究材料.自20世纪40年代秦仁昌先生发表水龙骨科自然分类系统, 中国石松类和蕨类植物研究就开始令世界瞩目.尤其是2017年第19届国际植物学大会在中国深圳召开后的5年时间里, 中国石松类和蕨类植物研究更是面向世界、走向国际, 研究更为广泛的科学问题, 在物种多样性、植物保护、系统演化和生态适应性等方面取得了一系列重要进展(Qi et al, 2018; Shen et al, 2018; Huang et al, 2020; Du et al, 2021; Wei R et al, 2021; Wang et al, 2022).本文对2017-2022年中国石松类和蕨类植物的物种多样性、保护、系统演化和生态适应性等研究进展进行综述, 以期为未来中国石松类和蕨类植物的研究提供参考. ...
石松类和蕨类植物大系统.自1940年中国蕨类植物分类学创始人秦仁昌先生划时代地将传统水龙骨科重新划分以来, 世界石松类和蕨类植物分类系统的研究进入了快速发展阶段, 但很多关键类群的系统位置仍然存在较大争议(Smith et al, 2006; Christenhusz et al, 2011; PPG I, 2016).2017-2022年来, 中国学者通过系统发育基因组学研究对世界石松类和蕨类植物系统演化框架的重建做出了重要贡献.2018年, 中国两个蕨类研究团队几乎同时基于转录组测序的系统发育基因组学研究重建了世界石松类和蕨类科级的系统发育框架, 解决了数个过往长期存在争议的研究类群的问题, 如木贼目的系统位置、里白科与双扇蕨科的亲缘关系、碗蕨科与凤尾蕨科、水龙骨目的亲缘关系等(Qi et al, 2018; Shen et al, 2018).同时, 基于构建的世界石松类和蕨类植物的系统演化树, 重建了石松类和蕨类植物关键性状孢子囊环带的演化模式(Shen et al, 2018), 比较分析了石松类和蕨类植物转录因子家族的演化模式(Qi et al, 2018).然而, 基于质体基因组数据构建的石松类和蕨类植物系统发育关系与基于转录组数据的结果仍然存在较大差异, 特别是木贼目和膜蕨目等的系统位置, 这些差异可能与质体基因组数据集中存在系统发育冲突信号有关(Kuo et al, 2018).随着研究的深入, 石松类和蕨类植物科级水平的系统框架已基本确定, 然而由于前期研究中缺少多个国外科属的材料, 世界石松类和蕨类植物的完整系统框架仍需要进一步研究. ... 1 2021 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 1 2021 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... Functional traits: Adaption of ferns in forest 2 2021 ... 石松类和蕨类植物广布于世界各地, 多以草本形式生长在林下、林缘或林窗中, 是森林演替早期阶段的先锋植物.蕨类植物对环境的适应和生态学意义一直受到研究人员的关注.但近年来, 我国对蕨类植物的生态学研究进展较少, 主要集中在蕨类植物的生态修复和生态适应上.在生态修复方面, Yang等(2022)研究发现, 先锋蕨类植物芒萁属能在降雨过后通过有性繁殖和克隆繁殖两种方式快速覆盖受损裸地, 形成密集蕨类灌丛, 在忍耐地表高温、贫瘠土壤、强烈光照的同时, 还有利于改善微环境, 促进土壤养分积累和抵御外来种入侵; Yan等(2019)通过三代全长转录组、二代差异转录组和液泡膜蛋白质组的方法揭示了蜈蚣草(Pteris vittata)的砷超富集机理及其调控分子网络, 对利用植物修复治理砷污染土壤具有重要意义.在生态适应上, Jin等(2021)通过测量345种蕨类植物对环境光和水可利用性敏感的功能性状发现, 鳞毛蕨类和水龙骨类的部分物种可以耐受强光和干旱的生境, 这些类群可能是蕨类植物适应干旱和明亮生境的先驱; Liang等(2022)以亚洲温带广布的二倍体同型孢子蕨类——中华蹄盖蕨复合群(Athyrium sinense complex)为研究对象, 通过群体遗传分析和分布区模拟等揭示了蕨类植物孢子强扩散能力下生态适应塑造的遗传分化格局.Dai等(2020)通过比较不同干扰水平下蕨类植物的分类、系统发育和功能多样性, 发现蕨类植物的多样性在很大程度上受到人类干扰水平的影响, 可作为环境胁迫的一个良好指示物种. ...
石松类和蕨类植物大系统.自1940年中国蕨类植物分类学创始人秦仁昌先生划时代地将传统水龙骨科重新划分以来, 世界石松类和蕨类植物分类系统的研究进入了快速发展阶段, 但很多关键类群的系统位置仍然存在较大争议(Smith et al, 2006; Christenhusz et al, 2011; PPG I, 2016).2017-2022年来, 中国学者通过系统发育基因组学研究对世界石松类和蕨类植物系统演化框架的重建做出了重要贡献.2018年, 中国两个蕨类研究团队几乎同时基于转录组测序的系统发育基因组学研究重建了世界石松类和蕨类科级的系统发育框架, 解决了数个过往长期存在争议的研究类群的问题, 如木贼目的系统位置、里白科与双扇蕨科的亲缘关系、碗蕨科与凤尾蕨科、水龙骨目的亲缘关系等(Qi et al, 2018; Shen et al, 2018).同时, 基于构建的世界石松类和蕨类植物的系统演化树, 重建了石松类和蕨类植物关键性状孢子囊环带的演化模式(Shen et al, 2018), 比较分析了石松类和蕨类植物转录因子家族的演化模式(Qi et al, 2018).然而, 基于质体基因组数据构建的石松类和蕨类植物系统发育关系与基于转录组数据的结果仍然存在较大差异, 特别是木贼目和膜蕨目等的系统位置, 这些差异可能与质体基因组数据集中存在系统发育冲突信号有关(Kuo et al, 2018).随着研究的深入, 石松类和蕨类植物科级水平的系统框架已基本确定, 然而由于前期研究中缺少多个国外科属的材料, 世界石松类和蕨类植物的完整系统框架仍需要进一步研究. ... 海南文昌维管植物区系特征分析 1 2018 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 海南文昌维管植物区系特征分析 1 2018 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 药用蕨类植物的研究进展 1 2009 ... 新版名录的一个重要特征是在科级和属级水平对一些重要的类群进行了所有种的保护.在科属级别进行全部保护的石松类和蕨类植物有7个类群, 包括石杉属、马尾杉属、水韭属(Isoetes)、观音座莲属、金毛狗属(Cibotium)、水蕨属(Ceratopteris)和桫椤科(除小黑桫椤和粗齿桫椤).一方面, 因为这些类群中的绝大多数物种为经济濒危种(严岳鸿等, 2006), 具有重要的药用、食用和观赏价值, 很容易受到人为采挖破坏.例如, 石杉属和马尾杉属植物广布于热带和亚热带地区, 在我国主产西南和华南, 东北、西北及华东也有分布.由于含有治疗阿尔兹海默症的有效成分——石杉碱甲(Ma et al, 2006, 2007), 而被作为重要的传统药材广泛利用(李国树和徐成东, 2009; 赵刚等, 2018).而且, 由于石杉碱甲的化学合成步骤繁杂、条件苛刻、成本高、收率低等问题, 未能通过人工化学合成而实现量产(Ferreira et al, 2016), 目前仍然主要从植物中提取, 因此对野生石杉属和马尾杉属植物消耗巨大(马小军等, 2009; 赵刚等, 2018; 谢峻等, 2020).然而, 石杉属和马尾杉属植物的植株生长缓慢、孢子萌发时间长、种群更新能力弱(鞠錾, 2009①( 鞠錾 (2009) 四种石杉科植物中产生石杉碱甲的内生真菌的分离、鉴定和其石杉碱甲含量的测定. 硕士学位论文, 复旦大学, 上海.); 齐耀东和王德立, 2017; 肖友利等, 2020), 其中石杉属植物完成一个完整的生活史约需15-20年之久(齐耀东和王德立, 2017), 一旦植物遭到破坏就很难再恢复(陈思思等, 2021).另一方面, 这些类群中大多数物种为演化濒危种, 即处于演化系统基部类群的孑遗物种(严岳鸿等, 2006).它们在地理分布区上表现出孑遗性、间断性、边缘性或孤立性; 环境变化或人为活动的长期干扰可能导致其分布区正逐渐缩小, 如桫椤科、莲座蕨属等.另外, 这些类群还存在较强的物种相似性, 即与同属的其他物种容易混淆(鲁兆莉等, 2021), 在整个属级水平进行保护才有可能实现对濒危野生蕨类植物的有效保护, 具有更好的可操作性. ... 药用蕨类植物的研究进展 1 2009 ... 新版名录的一个重要特征是在科级和属级水平对一些重要的类群进行了所有种的保护.在科属级别进行全部保护的石松类和蕨类植物有7个类群, 包括石杉属、马尾杉属、水韭属(Isoetes)、观音座莲属、金毛狗属(Cibotium)、水蕨属(Ceratopteris)和桫椤科(除小黑桫椤和粗齿桫椤).一方面, 因为这些类群中的绝大多数物种为经济濒危种(严岳鸿等, 2006), 具有重要的药用、食用和观赏价值, 很容易受到人为采挖破坏.例如, 石杉属和马尾杉属植物广布于热带和亚热带地区, 在我国主产西南和华南, 东北、西北及华东也有分布.由于含有治疗阿尔兹海默症的有效成分——石杉碱甲(Ma et al, 2006, 2007), 而被作为重要的传统药材广泛利用(李国树和徐成东, 2009; 赵刚等, 2018).而且, 由于石杉碱甲的化学合成步骤繁杂、条件苛刻、成本高、收率低等问题, 未能通过人工化学合成而实现量产(Ferreira et al, 2016), 目前仍然主要从植物中提取, 因此对野生石杉属和马尾杉属植物消耗巨大(马小军等, 2009; 赵刚等, 2018; 谢峻等, 2020).然而, 石杉属和马尾杉属植物的植株生长缓慢、孢子萌发时间长、种群更新能力弱(鞠錾, 2009①( 鞠錾 (2009) 四种石杉科植物中产生石杉碱甲的内生真菌的分离、鉴定和其石杉碱甲含量的测定. 硕士学位论文, 复旦大学, 上海.); 齐耀东和王德立, 2017; 肖友利等, 2020), 其中石杉属植物完成一个完整的生活史约需15-20年之久(齐耀东和王德立, 2017), 一旦植物遭到破坏就很难再恢复(陈思思等, 2021).另一方面, 这些类群中大多数物种为演化濒危种, 即处于演化系统基部类群的孑遗物种(严岳鸿等, 2006).它们在地理分布区上表现出孑遗性、间断性、边缘性或孤立性; 环境变化或人为活动的长期干扰可能导致其分布区正逐渐缩小, 如桫椤科、莲座蕨属等.另外, 这些类群还存在较强的物种相似性, 即与同属的其他物种容易混淆(鲁兆莉等, 2021), 在整个属级水平进行保护才有可能实现对濒危野生蕨类植物的有效保护, 具有更好的可操作性. ... 南涧大龙潭州级自然保护区植物资源与区系研究 1 2022 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 南涧大龙潭州级自然保护区植物资源与区系研究 1 2022 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 珠海淇澳岛石松类和蕨类植物区系特点及其资源利用分析 1 2022 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 珠海淇澳岛石松类和蕨类植物区系特点及其资源利用分析 1 2022 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... Genomes of leafy and leafless Platanthera orchids illuminate the evolution of mycoheterotrophy 1 2022 ... 石松类植物是维管植物最早分化的类群, 而蕨类植物与种子植物互为姐妹类群(Pryer et al, 2001).作为陆生维管植物演化的两个关键类群, 石松类和蕨类植物对于完整理解陆生维管植物的起源和演化历史上重要的共性科学问题具有关键作用, 如根、叶、芽等器官的起源和发育机制、气孔调控的转换机制、光合作用的演化模式、维管组织的起源、演化和发育机制、植物与昆虫和微生物等的协同演化等等, 这些关键共性科学问题的研究应重点关注.植物与昆虫、植物与微生物的协同演化一直以来都是植物科学家高度关注的热点科学问题(Jiang et al, 2017; Li et al, 2022).在石松类和蕨类植物中, 植物与昆虫的协同演化研究也取得一些重要进展(Shukla et al, 2016; Huang et al, 2021), 在石松类和蕨类植物的系统框架下研究植物与昆虫, 特别是与微生物的协同演化几乎无人问津. ... 荷叶铁线蕨孢子的离体繁殖 1 2008 ... Spore reproduction success of lycophytes and ferns under state priority conservation
石松类和蕨类植物大系统.自1940年中国蕨类植物分类学创始人秦仁昌先生划时代地将传统水龙骨科重新划分以来, 世界石松类和蕨类植物分类系统的研究进入了快速发展阶段, 但很多关键类群的系统位置仍然存在较大争议(Smith et al, 2006; Christenhusz et al, 2011; PPG I, 2016).2017-2022年来, 中国学者通过系统发育基因组学研究对世界石松类和蕨类植物系统演化框架的重建做出了重要贡献.2018年, 中国两个蕨类研究团队几乎同时基于转录组测序的系统发育基因组学研究重建了世界石松类和蕨类科级的系统发育框架, 解决了数个过往长期存在争议的研究类群的问题, 如木贼目的系统位置、里白科与双扇蕨科的亲缘关系、碗蕨科与凤尾蕨科、水龙骨目的亲缘关系等(Qi et al, 2018; Shen et al, 2018).同时, 基于构建的世界石松类和蕨类植物的系统演化树, 重建了石松类和蕨类植物关键性状孢子囊环带的演化模式(Shen et al, 2018), 比较分析了石松类和蕨类植物转录因子家族的演化模式(Qi et al, 2018).然而, 基于质体基因组数据构建的石松类和蕨类植物系统发育关系与基于转录组数据的结果仍然存在较大差异, 特别是木贼目和膜蕨目等的系统位置, 这些差异可能与质体基因组数据集中存在系统发育冲突信号有关(Kuo et al, 2018).随着研究的深入, 石松类和蕨类植物科级水平的系统框架已基本确定, 然而由于前期研究中缺少多个国外科属的材料, 世界石松类和蕨类植物的完整系统框架仍需要进一步研究. ... 浙江省长兴县湿地维管植物多样性及区系 1 2020 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 浙江省长兴县湿地维管植物多样性及区系 1 2020 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 中华水韭雌配子体及胚胎发育的初步研究 1 2013 ... Spore reproduction success of lycophytes and ferns under state priority conservation
石松类和蕨类植物大系统.自1940年中国蕨类植物分类学创始人秦仁昌先生划时代地将传统水龙骨科重新划分以来, 世界石松类和蕨类植物分类系统的研究进入了快速发展阶段, 但很多关键类群的系统位置仍然存在较大争议(Smith et al, 2006; Christenhusz et al, 2011; PPG I, 2016).2017-2022年来, 中国学者通过系统发育基因组学研究对世界石松类和蕨类植物系统演化框架的重建做出了重要贡献.2018年, 中国两个蕨类研究团队几乎同时基于转录组测序的系统发育基因组学研究重建了世界石松类和蕨类科级的系统发育框架, 解决了数个过往长期存在争议的研究类群的问题, 如木贼目的系统位置、里白科与双扇蕨科的亲缘关系、碗蕨科与凤尾蕨科、水龙骨目的亲缘关系等(Qi et al, 2018; Shen et al, 2018).同时, 基于构建的世界石松类和蕨类植物的系统演化树, 重建了石松类和蕨类植物关键性状孢子囊环带的演化模式(Shen et al, 2018), 比较分析了石松类和蕨类植物转录因子家族的演化模式(Qi et al, 2018).然而, 基于质体基因组数据构建的石松类和蕨类植物系统发育关系与基于转录组数据的结果仍然存在较大差异, 特别是木贼目和膜蕨目等的系统位置, 这些差异可能与质体基因组数据集中存在系统发育冲突信号有关(Kuo et al, 2018).随着研究的深入, 石松类和蕨类植物科级水平的系统框架已基本确定, 然而由于前期研究中缺少多个国外科属的材料, 世界石松类和蕨类植物的完整系统框架仍需要进一步研究. ... 珍稀药源植物蛇足石杉(千层塔)研究进展 1 2009 ... 新版名录的一个重要特征是在科级和属级水平对一些重要的类群进行了所有种的保护.在科属级别进行全部保护的石松类和蕨类植物有7个类群, 包括石杉属、马尾杉属、水韭属(Isoetes)、观音座莲属、金毛狗属(Cibotium)、水蕨属(Ceratopteris)和桫椤科(除小黑桫椤和粗齿桫椤).一方面, 因为这些类群中的绝大多数物种为经济濒危种(严岳鸿等, 2006), 具有重要的药用、食用和观赏价值, 很容易受到人为采挖破坏.例如, 石杉属和马尾杉属植物广布于热带和亚热带地区, 在我国主产西南和华南, 东北、西北及华东也有分布.由于含有治疗阿尔兹海默症的有效成分——石杉碱甲(Ma et al, 2006, 2007), 而被作为重要的传统药材广泛利用(李国树和徐成东, 2009; 赵刚等, 2018).而且, 由于石杉碱甲的化学合成步骤繁杂、条件苛刻、成本高、收率低等问题, 未能通过人工化学合成而实现量产(Ferreira et al, 2016), 目前仍然主要从植物中提取, 因此对野生石杉属和马尾杉属植物消耗巨大(马小军等, 2009; 赵刚等, 2018; 谢峻等, 2020).然而, 石杉属和马尾杉属植物的植株生长缓慢、孢子萌发时间长、种群更新能力弱(鞠錾, 2009①( 鞠錾 (2009) 四种石杉科植物中产生石杉碱甲的内生真菌的分离、鉴定和其石杉碱甲含量的测定. 硕士学位论文, 复旦大学, 上海.); 齐耀东和王德立, 2017; 肖友利等, 2020), 其中石杉属植物完成一个完整的生活史约需15-20年之久(齐耀东和王德立, 2017), 一旦植物遭到破坏就很难再恢复(陈思思等, 2021).另一方面, 这些类群中大多数物种为演化濒危种, 即处于演化系统基部类群的孑遗物种(严岳鸿等, 2006).它们在地理分布区上表现出孑遗性、间断性、边缘性或孤立性; 环境变化或人为活动的长期干扰可能导致其分布区正逐渐缩小, 如桫椤科、莲座蕨属等.另外, 这些类群还存在较强的物种相似性, 即与同属的其他物种容易混淆(鲁兆莉等, 2021), 在整个属级水平进行保护才有可能实现对濒危野生蕨类植物的有效保护, 具有更好的可操作性. ... 珍稀药源植物蛇足石杉(千层塔)研究进展 1 2009 ... 新版名录的一个重要特征是在科级和属级水平对一些重要的类群进行了所有种的保护.在科属级别进行全部保护的石松类和蕨类植物有7个类群, 包括石杉属、马尾杉属、水韭属(Isoetes)、观音座莲属、金毛狗属(Cibotium)、水蕨属(Ceratopteris)和桫椤科(除小黑桫椤和粗齿桫椤).一方面, 因为这些类群中的绝大多数物种为经济濒危种(严岳鸿等, 2006), 具有重要的药用、食用和观赏价值, 很容易受到人为采挖破坏.例如, 石杉属和马尾杉属植物广布于热带和亚热带地区, 在我国主产西南和华南, 东北、西北及华东也有分布.由于含有治疗阿尔兹海默症的有效成分——石杉碱甲(Ma et al, 2006, 2007), 而被作为重要的传统药材广泛利用(李国树和徐成东, 2009; 赵刚等, 2018).而且, 由于石杉碱甲的化学合成步骤繁杂、条件苛刻、成本高、收率低等问题, 未能通过人工化学合成而实现量产(Ferreira et al, 2016), 目前仍然主要从植物中提取, 因此对野生石杉属和马尾杉属植物消耗巨大(马小军等, 2009; 赵刚等, 2018; 谢峻等, 2020).然而, 石杉属和马尾杉属植物的植株生长缓慢、孢子萌发时间长、种群更新能力弱(鞠錾, 2009①( 鞠錾 (2009) 四种石杉科植物中产生石杉碱甲的内生真菌的分离、鉴定和其石杉碱甲含量的测定. 硕士学位论文, 复旦大学, 上海.); 齐耀东和王德立, 2017; 肖友利等, 2020), 其中石杉属植物完成一个完整的生活史约需15-20年之久(齐耀东和王德立, 2017), 一旦植物遭到破坏就很难再恢复(陈思思等, 2021).另一方面, 这些类群中大多数物种为演化濒危种, 即处于演化系统基部类群的孑遗物种(严岳鸿等, 2006).它们在地理分布区上表现出孑遗性、间断性、边缘性或孤立性; 环境变化或人为活动的长期干扰可能导致其分布区正逐渐缩小, 如桫椤科、莲座蕨属等.另外, 这些类群还存在较强的物种相似性, 即与同属的其他物种容易混淆(鲁兆莉等, 2021), 在整个属级水平进行保护才有可能实现对濒危野生蕨类植物的有效保护, 具有更好的可操作性. ... A survey of potential huperzine A natural resources in China: The Huperziaceae 1 2006 ... 新版名录的一个重要特征是在科级和属级水平对一些重要的类群进行了所有种的保护.在科属级别进行全部保护的石松类和蕨类植物有7个类群, 包括石杉属、马尾杉属、水韭属(Isoetes)、观音座莲属、金毛狗属(Cibotium)、水蕨属(Ceratopteris)和桫椤科(除小黑桫椤和粗齿桫椤).一方面, 因为这些类群中的绝大多数物种为经济濒危种(严岳鸿等, 2006), 具有重要的药用、食用和观赏价值, 很容易受到人为采挖破坏.例如, 石杉属和马尾杉属植物广布于热带和亚热带地区, 在我国主产西南和华南, 东北、西北及华东也有分布.由于含有治疗阿尔兹海默症的有效成分——石杉碱甲(Ma et al, 2006, 2007), 而被作为重要的传统药材广泛利用(李国树和徐成东, 2009; 赵刚等, 2018).而且, 由于石杉碱甲的化学合成步骤繁杂、条件苛刻、成本高、收率低等问题, 未能通过人工化学合成而实现量产(Ferreira et al, 2016), 目前仍然主要从植物中提取, 因此对野生石杉属和马尾杉属植物消耗巨大(马小军等, 2009; 赵刚等, 2018; 谢峻等, 2020).然而, 石杉属和马尾杉属植物的植株生长缓慢、孢子萌发时间长、种群更新能力弱(鞠錾, 2009①( 鞠錾 (2009) 四种石杉科植物中产生石杉碱甲的内生真菌的分离、鉴定和其石杉碱甲含量的测定. 硕士学位论文, 复旦大学, 上海.); 齐耀东和王德立, 2017; 肖友利等, 2020), 其中石杉属植物完成一个完整的生活史约需15-20年之久(齐耀东和王德立, 2017), 一旦植物遭到破坏就很难再恢复(陈思思等, 2021).另一方面, 这些类群中大多数物种为演化濒危种, 即处于演化系统基部类群的孑遗物种(严岳鸿等, 2006).它们在地理分布区上表现出孑遗性、间断性、边缘性或孤立性; 环境变化或人为活动的长期干扰可能导致其分布区正逐渐缩小, 如桫椤科、莲座蕨属等.另外, 这些类群还存在较强的物种相似性, 即与同属的其他物种容易混淆(鲁兆莉等, 2021), 在整个属级水平进行保护才有可能实现对濒危野生蕨类植物的有效保护, 具有更好的可操作性. ... Huperzine A from Huperzia species-An ethnopharmacolgical review 1 2007 ... 新版名录的一个重要特征是在科级和属级水平对一些重要的类群进行了所有种的保护.在科属级别进行全部保护的石松类和蕨类植物有7个类群, 包括石杉属、马尾杉属、水韭属(Isoetes)、观音座莲属、金毛狗属(Cibotium)、水蕨属(Ceratopteris)和桫椤科(除小黑桫椤和粗齿桫椤).一方面, 因为这些类群中的绝大多数物种为经济濒危种(严岳鸿等, 2006), 具有重要的药用、食用和观赏价值, 很容易受到人为采挖破坏.例如, 石杉属和马尾杉属植物广布于热带和亚热带地区, 在我国主产西南和华南, 东北、西北及华东也有分布.由于含有治疗阿尔兹海默症的有效成分——石杉碱甲(Ma et al, 2006, 2007), 而被作为重要的传统药材广泛利用(李国树和徐成东, 2009; 赵刚等, 2018).而且, 由于石杉碱甲的化学合成步骤繁杂、条件苛刻、成本高、收率低等问题, 未能通过人工化学合成而实现量产(Ferreira et al, 2016), 目前仍然主要从植物中提取, 因此对野生石杉属和马尾杉属植物消耗巨大(马小军等, 2009; 赵刚等, 2018; 谢峻等, 2020).然而, 石杉属和马尾杉属植物的植株生长缓慢、孢子萌发时间长、种群更新能力弱(鞠錾, 2009①( 鞠錾 (2009) 四种石杉科植物中产生石杉碱甲的内生真菌的分离、鉴定和其石杉碱甲含量的测定. 硕士学位论文, 复旦大学, 上海.); 齐耀东和王德立, 2017; 肖友利等, 2020), 其中石杉属植物完成一个完整的生活史约需15-20年之久(齐耀东和王德立, 2017), 一旦植物遭到破坏就很难再恢复(陈思思等, 2021).另一方面, 这些类群中大多数物种为演化濒危种, 即处于演化系统基部类群的孑遗物种(严岳鸿等, 2006).它们在地理分布区上表现出孑遗性、间断性、边缘性或孤立性; 环境变化或人为活动的长期干扰可能导致其分布区正逐渐缩小, 如桫椤科、莲座蕨属等.另外, 这些类群还存在较强的物种相似性, 即与同属的其他物种容易混淆(鲁兆莉等, 2021), 在整个属级水平进行保护才有可能实现对濒危野生蕨类植物的有效保护, 具有更好的可操作性. ... Ancestral stomatal control results in a canalization of fern and lycophyte adaptation to drought 1 2013 ... 此外, 石松类和蕨类植物对于理解维管植物气孔和光合作用的适应性演化模式非常关键.与种子植物不同, 石松类和蕨类植物的气孔缺少对脱落酸的主动响应, 对于干旱胁迫或气体交换都是处于被动防御状态, 这表明在与石松类和蕨类植物分化后, 植物水分平衡发生了由被动代谢向主动控制的根本转变(Brodribb & McAdam, 2011; McAdam & Brodribb, 2013).然而, 目前对于气孔起源和演化、气孔开闭方式的转变机制等相关科学问题在石松类和蕨类植物中仍然知之甚少.光合作用是地球上最重要的化学反应, 其起源、演化和分子机制一直以来都是科学家关注的重点科学问题.在维管植物中除了C3光合作用以外, 还演化出了两种重要的碳浓缩机制, 即C4光合作用和CAM光合作用.在种子植物中, C3和C4两种光合作用已经开展了大量的基因组学和生理生态学研究(Yang et al, 2017; Wai et al, 2019; Wang et al, 2019; Schlüter & Weber, 2020; Schiller & Brautigam, 2021), 而在石松类和蕨类植物中, 目前仅发现存在CAM光合作用, 其起源和适应性演化机制研究相当缺乏.在石松类和蕨类植物中CAM光合作用有其特别之处, 如水韭属植物是起源最古老的水生CAM植物(Wickell et al, 2021)、鹿角蕨属植物中孢子叶和营养叶分别执行C3和CAM光合作用(Rut et al, 2008)等.理解石松类和蕨类植物的光合作用演化模式对于陆生维管植物光合作用的起源历史和演化机制非常关键, 但这方面几乎没有学者开展深入的研究. ... Molecular and environmental regulation of root development 1 2019 ... 根的出现是维管植物演化过程中适应陆地环境的关键一步, 目前普遍认为根的起源事件发生了两次, 分别产生了石松类植物为代表的二歧根(bifurcating root)和以真叶植物(蕨类植物和种子植物)为代表的多类型根, 如不定根(adventitious root)、内皮层衍生的侧根(endodermis-derived lateral root)、初生根(primary root)中柱鞘衍生的顶端侧根(pericycle-derived acropetal lateral root)和不定侧根(adventitious lateral root)等(Motte et al, 2019).虽然根的起源和发育已经取得了一定进展(于阳阳等, 2015), 发现了生长素和WOX基因家族在不定根生长和发育中的关键作用(Yu et al, 2020), 但利用石松类和蕨类植物研究整个维管植物框架下根的起源和演化机制仍然较少.同样, 相比于国外研究, 中国学者在石松类和蕨类植物中对维管植物的芽和叶的演化和发育也缺少系统研究(Vasco et al, 2013; Plackett et al, 2015). ... Biodiversity hotspots for conservation priorities 1 2000 ... 英国生态学家诺曼•麦尔在1988年提出了生物多样性热点地区(biodiversity hotspots)的概念, 他认识到这些热点生态系统在很小的地域面积内包含了极其丰富的物种多样性.2000年, 国际环保组织根据特有物种数量和受威胁程度两个评定标准确定了全球34个物种最丰富且受到威胁最大的生物多样性热点地区(Myers et al, 2000), 其中涉及中国的区域有3个, 即中国西南山区(Mountains of South-West China)、华南-中南半岛地区(Indo-Burma)和喜马拉雅地区(Himalayan).2015年, 为贯彻落实国务院批准发布的《中国生物多样性保护战略与行动计划(2011-2030年)》, 国家环境保护部组织开展了生物多样性保护优先区域边界的核定工作, 确定了35个中国生物多样性保护优先区域范围.结合2017-2022年发表的中国石松类和蕨类植物新物种、新种下分类群和国家新记录所在的省份和调查分布空间情况(表3), 可清晰发现西藏、四川为近年来石松类和蕨类植物调查薄弱的生物多样性热点区域.提高野外调查密度、加强学科队伍建设在一定程度上能促进生物多样性热点地区石松类和蕨类植物物种多样性的发现(表3).中国幅员辽阔, 气候地貌复杂多样, 物种丰富, 开展中国蕨类植物多样性热点区域本底调查和系统分类等工作仍任重道远. ... 生长素及其运输抑制剂对粗梗水蕨孢子萌发的影响 0 2017 生长素及其运输抑制剂对粗梗水蕨孢子萌发的影响 0 2017 Ferns: The missing link in shoot evolution and development 1 2015 ... 根的出现是维管植物演化过程中适应陆地环境的关键一步, 目前普遍认为根的起源事件发生了两次, 分别产生了石松类植物为代表的二歧根(bifurcating root)和以真叶植物(蕨类植物和种子植物)为代表的多类型根, 如不定根(adventitious root)、内皮层衍生的侧根(endodermis-derived lateral root)、初生根(primary root)中柱鞘衍生的顶端侧根(pericycle-derived acropetal lateral root)和不定侧根(adventitious lateral root)等(Motte et al, 2019).虽然根的起源和发育已经取得了一定进展(于阳阳等, 2015), 发现了生长素和WOX基因家族在不定根生长和发育中的关键作用(Yu et al, 2020), 但利用石松类和蕨类植物研究整个维管植物框架下根的起源和演化机制仍然较少.同样, 相比于国外研究, 中国学者在石松类和蕨类植物中对维管植物的芽和叶的演化和发育也缺少系统研究(Vasco et al, 2013; Plackett et al, 2015). ... A community-derived classification for extant lycophytes and ferns 11 2016 ... 传统的蕨类植物(pteridophytes)由两个不同的进化谱系, 即石松类(lycophytes)和蕨类植物(ferns)组成(Pryer et al, 2001; Smith et al, 2006; PPG I, 2016), 同时也是高等植物中配子体和孢子体都可以独立生活的植物类群(Haufler et al, 2016).根据化石记录, 石松类和蕨类植物最早可追溯到4亿年前, 是地球上起源最为古老的维管植物(Qi et al, 2018; Shen et al, 2018; 严岳鸿等, 2019; Huang et al, 2020), 同时也是维管植物中继被子植物之后的第二大类群.虽然被子植物在生态系统中占据了主导地位, 但蕨类植物也演化产生了适应多样化环境的形态和物种多样性, 在各生态系统中仍发挥着重要作用(Schneider et al, 2004; Ranker & Haufler, 2008; Huang et al, 2020; Du et al, 2021).中国幅员辽阔, 拥有复杂多样的地貌和气候条件, 是世界上石松类和蕨类植物最为丰富的地区之一.根据《中国生物物种名录》2022版(The Biodiversity Committee of Chinese Academy of Sciences, 2022)最新统计, 我国共有石松类3科12属165种4个种下分类群, 蕨类植物38科177属2,215种228个种下分类群, 为植物学家们提供了丰富的研究材料.自20世纪40年代秦仁昌先生发表水龙骨科自然分类系统, 中国石松类和蕨类植物研究就开始令世界瞩目.尤其是2017年第19届国际植物学大会在中国深圳召开后的5年时间里, 中国石松类和蕨类植物研究更是面向世界、走向国际, 研究更为广泛的科学问题, 在物种多样性、植物保护、系统演化和生态适应性等方面取得了一系列重要进展(Qi et al, 2018; Shen et al, 2018; Huang et al, 2020; Du et al, 2021; Wei R et al, 2021; Wang et al, 2022).本文对2017-2022年中国石松类和蕨类植物的物种多样性、保护、系统演化和生态适应性等研究进展进行综述, 以期为未来中国石松类和蕨类植物的研究提供参考. ...
以2017-2022年和2000-2022年为时间界线, 对中国石松类和蕨类植物新物种、新种下分类群和中国新记录所属省份进行统计.结果显示(表3), 我国在这2个时间段发表的石松类和蕨类植物新物种、新种下分类群和国家新记录主要集中分布在某些热点或优先保护区域, 如云南、贵州、广西等地.其中, 云南新类群分布最多, 2000-2022年和2017-2022年共计发表了71和33个新物种、17和0个种下分类群、11和1个中国新记录, 占中国近22年来新发表物种数量的1/4, 是国内外学者广泛关注的区域; 其次为贵州, 2000-2022年和2017-2022年共计发表了59和20个新物种、7和0个新种下分类群、2和0个中国新记录, 占中国近22年来新发表物种数量的1/6.然而, 同样位于生物多样性热点区域的西藏和四川, 近年来在被子植物新发现上存在较高的密度(杜诚等, 2021), 但在石松类和蕨类植物类群上并没有体现, 推测与这些区域石松类和蕨类植物专类调查不充分、研究人员较少具有一定的关系. ...
以2017-2022年和2000-2022年为时间界线, 对中国石松类和蕨类植物新物种、新种下分类群和中国新记录所属省份进行统计.结果显示(表3), 我国在这2个时间段发表的石松类和蕨类植物新物种、新种下分类群和国家新记录主要集中分布在某些热点或优先保护区域, 如云南、贵州、广西等地.其中, 云南新类群分布最多, 2000-2022年和2017-2022年共计发表了71和33个新物种、17和0个种下分类群、11和1个中国新记录, 占中国近22年来新发表物种数量的1/4, 是国内外学者广泛关注的区域; 其次为贵州, 2000-2022年和2017-2022年共计发表了59和20个新物种、7和0个新种下分类群、2和0个中国新记录, 占中国近22年来新发表物种数量的1/6.然而, 同样位于生物多样性热点区域的西藏和四川, 近年来在被子植物新发现上存在较高的密度(杜诚等, 2021), 但在石松类和蕨类植物类群上并没有体现, 推测与这些区域石松类和蕨类植物专类调查不充分、研究人员较少具有一定的关系. ...
石松类和蕨类植物大系统.自1940年中国蕨类植物分类学创始人秦仁昌先生划时代地将传统水龙骨科重新划分以来, 世界石松类和蕨类植物分类系统的研究进入了快速发展阶段, 但很多关键类群的系统位置仍然存在较大争议(Smith et al, 2006; Christenhusz et al, 2011; PPG I, 2016).2017-2022年来, 中国学者通过系统发育基因组学研究对世界石松类和蕨类植物系统演化框架的重建做出了重要贡献.2018年, 中国两个蕨类研究团队几乎同时基于转录组测序的系统发育基因组学研究重建了世界石松类和蕨类科级的系统发育框架, 解决了数个过往长期存在争议的研究类群的问题, 如木贼目的系统位置、里白科与双扇蕨科的亲缘关系、碗蕨科与凤尾蕨科、水龙骨目的亲缘关系等(Qi et al, 2018; Shen et al, 2018).同时, 基于构建的世界石松类和蕨类植物的系统演化树, 重建了石松类和蕨类植物关键性状孢子囊环带的演化模式(Shen et al, 2018), 比较分析了石松类和蕨类植物转录因子家族的演化模式(Qi et al, 2018).然而, 基于质体基因组数据构建的石松类和蕨类植物系统发育关系与基于转录组数据的结果仍然存在较大差异, 特别是木贼目和膜蕨目等的系统位置, 这些差异可能与质体基因组数据集中存在系统发育冲突信号有关(Kuo et al, 2018).随着研究的深入, 石松类和蕨类植物科级水平的系统框架已基本确定, 然而由于前期研究中缺少多个国外科属的材料, 世界石松类和蕨类植物的完整系统框架仍需要进一步研究. ... 云南哀牢山地区森林附生维管植物多样性及区系特征 1 2018 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 云南哀牢山地区森林附生维管植物多样性及区系特征 1 2018 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 粗糙马尾杉的驯化栽培 1 2015 ... Spore reproduction success of lycophytes and ferns under state priority conservation
石松类和蕨类植物大系统.自1940年中国蕨类植物分类学创始人秦仁昌先生划时代地将传统水龙骨科重新划分以来, 世界石松类和蕨类植物分类系统的研究进入了快速发展阶段, 但很多关键类群的系统位置仍然存在较大争议(Smith et al, 2006; Christenhusz et al, 2011; PPG I, 2016).2017-2022年来, 中国学者通过系统发育基因组学研究对世界石松类和蕨类植物系统演化框架的重建做出了重要贡献.2018年, 中国两个蕨类研究团队几乎同时基于转录组测序的系统发育基因组学研究重建了世界石松类和蕨类科级的系统发育框架, 解决了数个过往长期存在争议的研究类群的问题, 如木贼目的系统位置、里白科与双扇蕨科的亲缘关系、碗蕨科与凤尾蕨科、水龙骨目的亲缘关系等(Qi et al, 2018; Shen et al, 2018).同时, 基于构建的世界石松类和蕨类植物的系统演化树, 重建了石松类和蕨类植物关键性状孢子囊环带的演化模式(Shen et al, 2018), 比较分析了石松类和蕨类植物转录因子家族的演化模式(Qi et al, 2018).然而, 基于质体基因组数据构建的石松类和蕨类植物系统发育关系与基于转录组数据的结果仍然存在较大差异, 特别是木贼目和膜蕨目等的系统位置, 这些差异可能与质体基因组数据集中存在系统发育冲突信号有关(Kuo et al, 2018).随着研究的深入, 石松类和蕨类植物科级水平的系统框架已基本确定, 然而由于前期研究中缺少多个国外科属的材料, 世界石松类和蕨类植物的完整系统框架仍需要进一步研究. ... 韶关南雄市观音岽自然保护区蕨类植物区系研究 1 2020 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 韶关南雄市观音岽自然保护区蕨类植物区系研究 1 2020 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 海南中部丘陵区热带次生林物种多样性及区系分析 1 2018 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 海南中部丘陵区热带次生林物种多样性及区系分析 1 2018 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 云南纳板河保护区蕨类植物区系地理研究 1 2017 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 云南纳板河保护区蕨类植物区系地理研究 1 2017 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 西沙群岛植物资源多样性调查与评价 1 2019 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 西沙群岛植物资源多样性调查与评价 1 2019 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 全基因组复制事件的绝对定年揭示莲座蕨属植物的迟滞演化 1 2021 ... 此外, 石松类和蕨类植物的适应性演化也开始受到国内学者的关注.其中, 在全基因组加倍(Zhang R et al, 2019; Huang et al, 2020; 汪浩等, 2019; 王婷等, 2021; Xia et al, 2022b)、自然杂交和基因家族扩张收缩(Zhang J et al, 2019; Xia et al, 2022a)等方面取得了一系列重要进展, 揭示了多倍化和自然杂交事件对于石松类和蕨类植物的适应性演化和物种形成具有重要作用, 这些进化事件也导致石松类和蕨类植物的系统发育网络错综复杂, 增加了系统构建的难度(Shu et al, 2022b). ... 全基因组复制事件的绝对定年揭示莲座蕨属植物的迟滞演化 1 2021 ... 此外, 石松类和蕨类植物的适应性演化也开始受到国内学者的关注.其中, 在全基因组加倍(Zhang R et al, 2019; Huang et al, 2020; 汪浩等, 2019; 王婷等, 2021; Xia et al, 2022b)、自然杂交和基因家族扩张收缩(Zhang J et al, 2019; Xia et al, 2022a)等方面取得了一系列重要进展, 揭示了多倍化和自然杂交事件对于石松类和蕨类植物的适应性演化和物种形成具有重要作用, 这些进化事件也导致石松类和蕨类植物的系统发育网络错综复杂, 增加了系统构建的难度(Shu et al, 2022b). ... 1 2018 ... Spore reproduction success of lycophytes and ferns under state priority conservation
石松类和蕨类植物大系统.自1940年中国蕨类植物分类学创始人秦仁昌先生划时代地将传统水龙骨科重新划分以来, 世界石松类和蕨类植物分类系统的研究进入了快速发展阶段, 但很多关键类群的系统位置仍然存在较大争议(Smith et al, 2006; Christenhusz et al, 2011; PPG I, 2016).2017-2022年来, 中国学者通过系统发育基因组学研究对世界石松类和蕨类植物系统演化框架的重建做出了重要贡献.2018年, 中国两个蕨类研究团队几乎同时基于转录组测序的系统发育基因组学研究重建了世界石松类和蕨类科级的系统发育框架, 解决了数个过往长期存在争议的研究类群的问题, 如木贼目的系统位置、里白科与双扇蕨科的亲缘关系、碗蕨科与凤尾蕨科、水龙骨目的亲缘关系等(Qi et al, 2018; Shen et al, 2018).同时, 基于构建的世界石松类和蕨类植物的系统演化树, 重建了石松类和蕨类植物关键性状孢子囊环带的演化模式(Shen et al, 2018), 比较分析了石松类和蕨类植物转录因子家族的演化模式(Qi et al, 2018).然而, 基于质体基因组数据构建的石松类和蕨类植物系统发育关系与基于转录组数据的结果仍然存在较大差异, 特别是木贼目和膜蕨目等的系统位置, 这些差异可能与质体基因组数据集中存在系统发育冲突信号有关(Kuo et al, 2018).随着研究的深入, 石松类和蕨类植物科级水平的系统框架已基本确定, 然而由于前期研究中缺少多个国外科属的材料, 世界石松类和蕨类植物的完整系统框架仍需要进一步研究. ... 浙江景宁畲族自治县石松类和蕨类植物区系研究 1 2019 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 浙江景宁畲族自治县石松类和蕨类植物区系研究 1 2019 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... Plastid phylogenomics resolve deep relationships among eupolypod II Ferns with rapid radiation and rate heterogeneity 3 2017 ... 在科级水平上, 卷柏科、凤尾蕨科、碗蕨科、铁角蕨科、叉蕨科、水龙骨科等世界性大科的系统学研究方面取得了一系列重要研究进展.其中卷柏科和铁角蕨科主要基于形态和分子证据来解决科内属/组间的系统发育关系(Zhou et al, 2016, 2022; Xu KW et al, 2018; Xu et al, 2020; Zhang HR et al, 2020; Zhang MH et al, 2022)和厘清各疑难复合类群(如小卷柏复合群(Selaginella helvetica complex)、红枝卷柏复合群(S. sanguinolenta complex)、北京铁角蕨复合群(Asplenium pekinense complex)、变异铁角蕨复合群(A. varians complex)等)的系统分类地位(Liang et al, 2021; Zhang MH et al, 2021, 2022).同时, 卷柏科特殊质体结构, 如具有标志性同向重复序列(DR)的环形结构、GC含量高、碱基替换速率快等特点也常受到人们的广泛关注(Zhang MH et al, 2021, 2022; Xiang et al, 2022; Zhou et al, 2022).其他备受关注的类群, 如水龙骨科、凤尾蕨科、碗蕨科和叉蕨科等则着重通过扩大抽样来厘清科间或科下的系统关系和演化历史(Wei R et al, 2017; Zhang et al, 2017a, b; He et al, 2018; Zhang & Zhang, 2018a, b; Wei & Zhang, 2020; Zhao et al, 2020; Du et al, 2021; Zhou et al, 2022).如Wei R等(2021)通过在全球范围内对水龙骨科关键类群进行取样, 结合高通量测序技术获取了28属78个代表物种的叶绿体全基因组数据, 构建了迄今为止该科最为稳定的系统框架; 并通过根状茎鳞片形态、叶片分裂式样、叶脉类型、孢子囊群类型、隔丝形态以及孢子形态等形态学证据确定了各分支界线. ...
石松类和蕨类植物大系统.自1940年中国蕨类植物分类学创始人秦仁昌先生划时代地将传统水龙骨科重新划分以来, 世界石松类和蕨类植物分类系统的研究进入了快速发展阶段, 但很多关键类群的系统位置仍然存在较大争议(Smith et al, 2006; Christenhusz et al, 2011; PPG I, 2016).2017-2022年来, 中国学者通过系统发育基因组学研究对世界石松类和蕨类植物系统演化框架的重建做出了重要贡献.2018年, 中国两个蕨类研究团队几乎同时基于转录组测序的系统发育基因组学研究重建了世界石松类和蕨类科级的系统发育框架, 解决了数个过往长期存在争议的研究类群的问题, 如木贼目的系统位置、里白科与双扇蕨科的亲缘关系、碗蕨科与凤尾蕨科、水龙骨目的亲缘关系等(Qi et al, 2018; Shen et al, 2018).同时, 基于构建的世界石松类和蕨类植物的系统演化树, 重建了石松类和蕨类植物关键性状孢子囊环带的演化模式(Shen et al, 2018), 比较分析了石松类和蕨类植物转录因子家族的演化模式(Qi et al, 2018).然而, 基于质体基因组数据构建的石松类和蕨类植物系统发育关系与基于转录组数据的结果仍然存在较大差异, 特别是木贼目和膜蕨目等的系统位置, 这些差异可能与质体基因组数据集中存在系统发育冲突信号有关(Kuo et al, 2018).随着研究的深入, 石松类和蕨类植物科级水平的系统框架已基本确定, 然而由于前期研究中缺少多个国外科属的材料, 世界石松类和蕨类植物的完整系统框架仍需要进一步研究. ... 1 2021 ... 随着我国综合国力的增强和植物学研究的长足发展, 国内研究学者开始走出国门, 联合其他国家的科研院所到国外进行植物考察.在2017-2022年期间, 我国植物学家参与并完成了东非肯尼亚和加那利群岛维管植物的多样性调查和编目, 分别记载2个地区石松类和蕨类植物22科60属140种(周亚东, 2017)和21科30属63种, 并于2021年出版了Flora of Canary Islands (Xin et al, 2021).Shang等(2018)发表了亚洲太平洋地区的蕨类植物新属卫蕨属(Hiya), 并对马达加斯加(Shang et al, 2020)和亚洲太平洋地区(Shang et al, 2021)翼盖蕨属(Didymochlaena)植物进行修订并发表了多个新分类群; Wei等(2022)对亚洲芒萁属(Dicranopteris)植物进行分类修订并发表了新种巴厘岛芒萁(Dicranopteris baliensis); Zhang MH等(2021)、Wu等(2017)、Dong等(2022)对越南的卷柏科、叉蕨科等植物进行研究, 发表分布于越南的新种Selaginella pseudotamariscina、S. guihaia和Tectaria fungii.协助其他国家完成植物调查和编目, 在了解世界生物多样性的同时, 也提高了我国植物研究在世界舞台上的影响力, 并为我国研究院所和高校师生提供了第一手学习资料. ... 江西九连山自然保护区蕨类植物资源调查和分析 1 2021 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 江西九连山自然保护区蕨类植物资源调查和分析 1 2021 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... A global plastid phylogeny of the fern genus Asplenium (Aspleniaceae) 2 2020 ... 在科级水平上, 卷柏科、凤尾蕨科、碗蕨科、铁角蕨科、叉蕨科、水龙骨科等世界性大科的系统学研究方面取得了一系列重要研究进展.其中卷柏科和铁角蕨科主要基于形态和分子证据来解决科内属/组间的系统发育关系(Zhou et al, 2016, 2022; Xu KW et al, 2018; Xu et al, 2020; Zhang HR et al, 2020; Zhang MH et al, 2022)和厘清各疑难复合类群(如小卷柏复合群(Selaginella helvetica complex)、红枝卷柏复合群(S. sanguinolenta complex)、北京铁角蕨复合群(Asplenium pekinense complex)、变异铁角蕨复合群(A. varians complex)等)的系统分类地位(Liang et al, 2021; Zhang MH et al, 2021, 2022).同时, 卷柏科特殊质体结构, 如具有标志性同向重复序列(DR)的环形结构、GC含量高、碱基替换速率快等特点也常受到人们的广泛关注(Zhang MH et al, 2021, 2022; Xiang et al, 2022; Zhou et al, 2022).其他备受关注的类群, 如水龙骨科、凤尾蕨科、碗蕨科和叉蕨科等则着重通过扩大抽样来厘清科间或科下的系统关系和演化历史(Wei R et al, 2017; Zhang et al, 2017a, b; He et al, 2018; Zhang & Zhang, 2018a, b; Wei & Zhang, 2020; Zhao et al, 2020; Du et al, 2021; Zhou et al, 2022).如Wei R等(2021)通过在全球范围内对水龙骨科关键类群进行取样, 结合高通量测序技术获取了28属78个代表物种的叶绿体全基因组数据, 构建了迄今为止该科最为稳定的系统框架; 并通过根状茎鳞片形态、叶片分裂式样、叶脉类型、孢子囊群类型、隔丝形态以及孢子形态等形态学证据确定了各分支界线. ...
石松类和蕨类植物大系统.自1940年中国蕨类植物分类学创始人秦仁昌先生划时代地将传统水龙骨科重新划分以来, 世界石松类和蕨类植物分类系统的研究进入了快速发展阶段, 但很多关键类群的系统位置仍然存在较大争议(Smith et al, 2006; Christenhusz et al, 2011; PPG I, 2016).2017-2022年来, 中国学者通过系统发育基因组学研究对世界石松类和蕨类植物系统演化框架的重建做出了重要贡献.2018年, 中国两个蕨类研究团队几乎同时基于转录组测序的系统发育基因组学研究重建了世界石松类和蕨类科级的系统发育框架, 解决了数个过往长期存在争议的研究类群的问题, 如木贼目的系统位置、里白科与双扇蕨科的亲缘关系、碗蕨科与凤尾蕨科、水龙骨目的亲缘关系等(Qi et al, 2018; Shen et al, 2018).同时, 基于构建的世界石松类和蕨类植物的系统演化树, 重建了石松类和蕨类植物关键性状孢子囊环带的演化模式(Shen et al, 2018), 比较分析了石松类和蕨类植物转录因子家族的演化模式(Qi et al, 2018).然而, 基于质体基因组数据构建的石松类和蕨类植物系统发育关系与基于转录组数据的结果仍然存在较大差异, 特别是木贼目和膜蕨目等的系统位置, 这些差异可能与质体基因组数据集中存在系统发育冲突信号有关(Kuo et al, 2018).随着研究的深入, 石松类和蕨类植物科级水平的系统框架已基本确定, 然而由于前期研究中缺少多个国外科属的材料, 世界石松类和蕨类植物的完整系统框架仍需要进一步研究. ... 印江车家河湿地公园维管植物资源分析 1 2017 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 印江车家河湿地公园维管植物资源分析 1 2017 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... Plastome- based phylogenomics resolves the placement of the sanguinolenta group in the spikemoss of lycophyte (Selaginellaceae) 1 2020 ... 在科级水平上, 卷柏科、凤尾蕨科、碗蕨科、铁角蕨科、叉蕨科、水龙骨科等世界性大科的系统学研究方面取得了一系列重要研究进展.其中卷柏科和铁角蕨科主要基于形态和分子证据来解决科内属/组间的系统发育关系(Zhou et al, 2016, 2022; Xu KW et al, 2018; Xu et al, 2020; Zhang HR et al, 2020; Zhang MH et al, 2022)和厘清各疑难复合类群(如小卷柏复合群(Selaginella helvetica complex)、红枝卷柏复合群(S. sanguinolenta complex)、北京铁角蕨复合群(Asplenium pekinense complex)、变异铁角蕨复合群(A. varians complex)等)的系统分类地位(Liang et al, 2021; Zhang MH et al, 2021, 2022).同时, 卷柏科特殊质体结构, 如具有标志性同向重复序列(DR)的环形结构、GC含量高、碱基替换速率快等特点也常受到人们的广泛关注(Zhang MH et al, 2021, 2022; Xiang et al, 2022; Zhou et al, 2022).其他备受关注的类群, 如水龙骨科、凤尾蕨科、碗蕨科和叉蕨科等则着重通过扩大抽样来厘清科间或科下的系统关系和演化历史(Wei R et al, 2017; Zhang et al, 2017a, b; He et al, 2018; Zhang & Zhang, 2018a, b; Wei & Zhang, 2020; Zhao et al, 2020; Du et al, 2021; Zhou et al, 2022).如Wei R等(2021)通过在全球范围内对水龙骨科关键类群进行取样, 结合高通量测序技术获取了28属78个代表物种的叶绿体全基因组数据, 构建了迄今为止该科最为稳定的系统框架; 并通过根状茎鳞片形态、叶片分裂式样、叶脉类型、孢子囊群类型、隔丝形态以及孢子形态等形态学证据确定了各分支界线. ... The unique evolutionary trajectory and dynamic conformations of DR and IR/DR coexisting plastomes of the early vascular plant Selaginellaceae (Lycophyte) 0 2019 Transcriptomic evidence of adaptive evolution of the epiphytic fern Asplenium nidus 1 2019 ... 此外, 石松类和蕨类植物的适应性演化也开始受到国内学者的关注.其中, 在全基因组加倍(Zhang R et al, 2019; Huang et al, 2020; 汪浩等, 2019; 王婷等, 2021; Xia et al, 2022b)、自然杂交和基因家族扩张收缩(Zhang J et al, 2019; Xia et al, 2022a)等方面取得了一系列重要进展, 揭示了多倍化和自然杂交事件对于石松类和蕨类植物的适应性演化和物种形成具有重要作用, 这些进化事件也导致石松类和蕨类植物的系统发育网络错综复杂, 增加了系统构建的难度(Shu et al, 2022b). ... 海南五指山国家级自然保护区石松类和蕨类植物区系研究 1 2017 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... 海南五指山国家级自然保护区石松类和蕨类植物区系研究 1 2017 ... 植物区系地理研究对了解某一区域的植物种类组成、地理成分以及区系之间亲缘关系具有重要意义(严岳鸿等, 2014).2017-2022年来, 我国石松类和蕨类植物区系地理研究的报道不胜枚举, 研究地域主要集中在贵州、海南、广东、江西、浙江、云南等省份(陈志红等, 2017; 康婉媚等, 2017; 王倩等,2017; 张潮等, 2017; 张凯等, 2017; 张廷跃和李艳, 2017; 赵清, 2017①( 赵清 (2017) 惠东县栋梁嶂森林公园植物区系及资源分析. 硕士学位论文, 仲恺农业工程学院, 广州.); 朱晓峰和徐梅梅, 2017; 丁扬等, 2018; 姬红利等, 2018; 季必浩等, 2018; 孔华清等, 2018; 李冬琳等, 2018; 汤丹丹等, 2018; 王牌等, 2018; 夏顺颖, 2018; 张伟清等, 2018; 陈日红等, 2019; 林汝强等, 2019; 沈秋慧, 2019; 宋佳昱, 2019②(② 宋佳昱 (2019) 海南琼海白石岭自然植物群落及野生观赏植物资源调查应用研究. 硕士学位论文, 海南大学, 海口.); 王清隆等, 2019; 王宗琪等, 2019; 陈莉娟等, 2020; 邓夏雨等, 2020; 邓贤兰和徐佳红, 2020; 杜晓洁等, 2020; 侯天文等, 2020; 胡根秀等, 2020; 胡伟等, 2020; 刘政等, 2020; 王梦楠等, 2020; 蔡国俊等, 2021; 胡佳玉等, 2021; 徐国良和曾晓辉, 2021; 杨礼旦, 2021; 代亮亮等, 2022; 黎明等, 2022; 李金亮等, 2022; 王德芬等, 2022; 杨丹等, 2022), 与采集密度数据大体一致(表3).从这些研究报道来看, 涉及到石松类和蕨类植物的区系研究主要集中在某一个山区或保护区内小范围地开展, 而对于更大范围甚至是全国性的石松类和蕨类植物区系研究鲜有报道.针对某一地区开展的调查研究或某一时期国内石松类和蕨类植物物种统计结果整理成研究报告或者专著, 也是对物种多样性的思考和总结.2017-2022年间, 一系列石松类和蕨类植物调查研究的专著被出版, 如《中国茂兰石松类与蕨类植物》(张宪春和姚正明, 2017)、《梵净山蕨类植物》(苟光前等, 2017)、《海南蕨类植物》(严岳鸿和周喜乐, 2018)、《中国武陵山区蕨类植物》(严岳鸿和周喜乐, 2021)、《华东石松类与蕨类植物多样性编目》(金冬梅和严岳鸿, 2021)、《广州石松类和蕨类植物》(董仕勇, 2022)等.尽管我国石松类和蕨类植物区系调查研究在近年来取得较好的成绩, 但仍存在许多调查空白或薄弱区域, 例如华北地区和西北西区(表3). ... Evolutionary relationships of the ancient fern lineage the adder’s tongues (Ophioglossaceae) with description of Sahashia gen 1 2020 ... 在属级水平上, Zhao等(2020)基于8个高变异位点的DNA分子序列构建了71种瓦韦属(Lepisorus)及其近缘属的系统发育框架, 结合25个形态数据编码和遗传学证据, 解决了瓦韦属及其近缘类群深层的系统发育关系, 将原先扇蕨属(Neocheiropteris)、盾蕨属(Neolepisorus)、伏石蕨属(Lemmaphyllum)、鳞果星蕨属(Lepidomicrosorium)、毛鳞蕨属(Tricholepidium)等都纳入瓦韦属的范畴; Zhang L等(2017a, b,2018a, b, 2020)采用多个叶绿体片段和核基因数据重建了世界范围内凤尾蕨属(Pteris)、叉蕨属(Tectaria)、鹿角蕨亚科以及瓶尔小草科的系统发育关系并根据形态特征划定属间界线, 促进了蕨类植物的分类学和系统学研究进展; Shang等(2018)利用rbcL、atpA、rps4、rpl16 4个叶绿体基因片段重建了世界碗蕨科的系统发育关系和性状演化, 确定了国外产蕨类植物新属卫蕨属的独立地位. ... A classification of the fern genus Tectaria (Tectariaceae: Polypodiales) based on molecular and morphological evidence 3 2018a ... 在科级水平上, 卷柏科、凤尾蕨科、碗蕨科、铁角蕨科、叉蕨科、水龙骨科等世界性大科的系统学研究方面取得了一系列重要研究进展.其中卷柏科和铁角蕨科主要基于形态和分子证据来解决科内属/组间的系统发育关系(Zhou et al, 2016, 2022; Xu KW et al, 2018; Xu et al, 2020; Zhang HR et al, 2020; Zhang MH et al, 2022)和厘清各疑难复合类群(如小卷柏复合群(Selaginella helvetica complex)、红枝卷柏复合群(S. sanguinolenta complex)、北京铁角蕨复合群(Asplenium pekinense complex)、变异铁角蕨复合群(A. varians complex)等)的系统分类地位(Liang et al, 2021; Zhang MH et al, 2021, 2022).同时, 卷柏科特殊质体结构, 如具有标志性同向重复序列(DR)的环形结构、GC含量高、碱基替换速率快等特点也常受到人们的广泛关注(Zhang MH et al, 2021, 2022; Xiang et al, 2022; Zhou et al, 2022).其他备受关注的类群, 如水龙骨科、凤尾蕨科、碗蕨科和叉蕨科等则着重通过扩大抽样来厘清科间或科下的系统关系和演化历史(Wei R et al, 2017; Zhang et al, 2017a, b; He et al, 2018; Zhang & Zhang, 2018a, b; Wei & Zhang, 2020; Zhao et al, 2020; Du et al, 2021; Zhou et al, 2022).如Wei R等(2021)通过在全球范围内对水龙骨科关键类群进行取样, 结合高通量测序技术获取了28属78个代表物种的叶绿体全基因组数据, 构建了迄今为止该科最为稳定的系统框架; 并通过根状茎鳞片形态、叶片分裂式样、叶脉类型、孢子囊群类型、隔丝形态以及孢子形态等形态学证据确定了各分支界线. ...
本文的其它图/表
|