
生物多样性 ›› 2019, Vol. 27 ›› Issue (2): 159-167. DOI: 10.17520/biods.2018317 cstr: 32101.14.biods.2018317
王晓月1,2,3,朱鑫鑫3,杨娟3,刘云静3,汤晓辛1,2,3,*( )
)
收稿日期:2018-11-27
接受日期:2018-03-05
出版日期:2019-02-20
发布日期:2019-04-08
通讯作者:
汤晓辛
基金资助:
Wang Xiaoyue1,2,3,Zhu Xinxin3,Yang Juan3,Liu Yunjing3,Tang Xiaoxin1,2,3,*()
Received:2018-11-27
Accepted:2018-03-05
Online:2019-02-20
Published:2019-04-08
Contact:
Tang Xiaoxin
摘要:
植物个体内花型的变异影响繁殖成功, 雌性繁殖性状的变异可能影响雌性的繁殖成功, 也可能作为花粉的受体影响雄性的繁殖成功。然而, 植物个体内不同花柱长度的花产生的花粉是否影响植物的繁殖成功却少有研究。梅(Armeniaca mume)是原产我国的重要木本花卉和经济果树, 我们的野外观察发现在同一植株内, 不同花的花柱长度有变异, 存在长柱型、短柱型和雄花型(雌蕊败育)三种花型, 是比较雌性繁殖性状的变异对两性繁殖成功的影响的理想材料。本文主要测量了不同花型的花部特征, 统计花期, 并开展体外花粉萌发以及人工控制授粉实验。结果表明: 长柱型的花冠直径、雌蕊长、单花花粉数、花粉体积显著大于短柱型和雄花型。长柱型的单花期以及雌花期显著长于短柱型。长柱型、短柱型以及雄花型花粉在活体柱头上的萌发率没有显著性差异, 雄花型的花粉管长度显著高于长柱型和短柱型。长柱型为母本的花粉萌发率以及花粉管的长度要显著高于短柱型。长柱型、短柱型、雄花型花粉授粉与自然对照处理的坐果率没有显著性差异, 而长柱型为母本的坐果率要显著高于短柱型为母本的坐果率。这些结果表明野生梅花的长柱型为母本有利于花粉的萌发和花粉管的伸长, 有高的坐果率; 但其作为花粉供体的雄性功能与其他花型没有差异。
王晓月,朱鑫鑫,杨娟,刘云静,汤晓辛 (2019) 梅花个体内花柱长度的变异及其对繁殖成功的影响. 生物多样性, 27, 159-167. DOI: 10.17520/biods.2018317.
Wang Xiaoyue,Zhu Xinxin,Yang Juan,Liu Yunjing,Tang Xiaoxin (2019) Variation in style length and the effect on reproductive success in Chinese plums (Armeniaca mume). Biodiversity Science, 27, 159-167. DOI: 10.17520/biods.2018317.

图1 不同花柱长度花型的梅花。(A)长柱型、短柱型的花苞(花苞期即可区分); (B)长柱型; (C)短柱型; (D)雄花型。L: 长柱型; S: 短柱型; M: 雄花型。
Fig. 1 Flowers of Armeniaca mume showing different style lengths. (A) Buds of long-style type and short-style type (different type can be distinguished during the bud phase); (B) Long-style type; (C) Short-style type; (D) Male-flower type. L, Long-style type; S, Short-style type; M, Male-flower type.
| 花部特征 Flower character | 长柱型 Long-style type | 短柱型 Short-style type | 雄花型 Male-flower type | Wald χ2 | df | P |
|---|---|---|---|---|---|---|
| 花萼长 Spur length (mm) | 5.30 ± 0.09a | 5.38 ± 0.08a | 4.99 ± 0.10b | 8.970 | 2 | 0.011 |
| 花萼宽 Spur width (mm) | 3.62 ± 0.05a | 3.78 ± 0.07a | 3.76 ± 0.12a | 2.755 | 2 | 0.252 |
| 花冠直径 Corolla diameter (mm) | 24.42 ± 0.44a | 23.43 ± 0.23b | 21.64 ± 0.41c | 27.656 | 2 | < 0.001 |
| 花瓣长 Petal length (mm) | 10.44 ± 0.18a | 10.03 ± 0.16a | 9.17 ± 0.18b | 22.160 | 2 | < 0.001 |
| 花瓣宽 Petal width (mm) | 9.70 ± 0.20a | 9.31 ± 0.16a | 8.05 ± 0.25b | 32.449 | 2 | < 0.001 |
| 雄蕊数 Stamen Number | 53.71 ± 0.71b | 53.56 ± 0.79b | 56.93 ± 1.10a | 8.460 | 2 | 0.015 |
| 雄蕊长 Stamen length (mm) | 9.25 ± 0.21b | 10.39 ± 0.18a | 8.82 ± 0.22b | 33.193 | 2 | < 0.001 |
| 花药长 Anther length (mm) | 1.01 ± 0.02a | 1.02 ± 0.01a | 1.01 ± 0.02a | 0.500 | 2 | 0.779 |
| 雌蕊长 Pistil length (mm) | 14.46 ± 0.20a | 12.34 ± 0.21b | 3.42 ± 0.16c | 8.460 | 2 | 0.015 |
| 花柱长 Style length (mm) | 12.20 ± 0.19a | 10.18 ± 0.18b | / | 58.394 | 1 | < 0.001 |
| 花粉数 Pollen Number | 37,522.22 ± 1,305.85a | 30,386.67 ± 1,412.62b | 32,000.00 ± 2,003.29b | 15.133 | 2 | 0.001 |
| 花粉体积 Pollen volume (mm3) | 1,213.07 ± 28.87a | 1,102.47 ± 31.08b | 1,067.13 ± 49.45b | 10.226 | 2 | 0.006 |
| 胚珠数 Ovule Number | 2.00 ± 0.00a | 2.00 ± 0.00a | 0.90 ± 0.35b | 46.620 | 2 | < 0.001 |
| 单花期 Flower longevity (days) | 10.43 ± 0.24a | 9.29 ± 0.56b | 10.70 ± 0.30a | 6.620 | 2 | 0.037 |
| 雄花期 Male phase (days) | 5.46 ± 0.35a | 5.23 ± 0.52a | / | 0.140 | 1 | 0.079 |
| 雌花期 Female phase (days) | 4.96 ± 0.30a | 4.06 ± 0.30b | / | 4.670 | 1 | 0.031 |
表1 长柱型、短柱型、雄花型梅花花部特征(平均值 ± 标准误)的比较(广义线性模型)。表中数字加粗表示长柱型的指标显著大于短柱型和雄花型相应的指标。花部特征样本量为30, 花期样本量为20。不同小写字母表明不同花型对应的花部特征有显著性差异。
Table 1 Comparisons of floral traits (mean ± SE) among long-style, short-style and male-flower types within individuals of Armeniaca mume using generalized linear model (GLM). Floral traits of long-style type significantly larger than those of short-style and male-flower types are in bold. The sample size of floral characters and floral phase was 30 and 20, respectively. Different letters indicate significant differences in flower traits among three types in the same row.
| 花部特征 Flower character | 长柱型 Long-style type | 短柱型 Short-style type | 雄花型 Male-flower type | Wald χ2 | df | P |
|---|---|---|---|---|---|---|
| 花萼长 Spur length (mm) | 5.30 ± 0.09a | 5.38 ± 0.08a | 4.99 ± 0.10b | 8.970 | 2 | 0.011 |
| 花萼宽 Spur width (mm) | 3.62 ± 0.05a | 3.78 ± 0.07a | 3.76 ± 0.12a | 2.755 | 2 | 0.252 |
| 花冠直径 Corolla diameter (mm) | 24.42 ± 0.44a | 23.43 ± 0.23b | 21.64 ± 0.41c | 27.656 | 2 | < 0.001 |
| 花瓣长 Petal length (mm) | 10.44 ± 0.18a | 10.03 ± 0.16a | 9.17 ± 0.18b | 22.160 | 2 | < 0.001 |
| 花瓣宽 Petal width (mm) | 9.70 ± 0.20a | 9.31 ± 0.16a | 8.05 ± 0.25b | 32.449 | 2 | < 0.001 |
| 雄蕊数 Stamen Number | 53.71 ± 0.71b | 53.56 ± 0.79b | 56.93 ± 1.10a | 8.460 | 2 | 0.015 |
| 雄蕊长 Stamen length (mm) | 9.25 ± 0.21b | 10.39 ± 0.18a | 8.82 ± 0.22b | 33.193 | 2 | < 0.001 |
| 花药长 Anther length (mm) | 1.01 ± 0.02a | 1.02 ± 0.01a | 1.01 ± 0.02a | 0.500 | 2 | 0.779 |
| 雌蕊长 Pistil length (mm) | 14.46 ± 0.20a | 12.34 ± 0.21b | 3.42 ± 0.16c | 8.460 | 2 | 0.015 |
| 花柱长 Style length (mm) | 12.20 ± 0.19a | 10.18 ± 0.18b | / | 58.394 | 1 | < 0.001 |
| 花粉数 Pollen Number | 37,522.22 ± 1,305.85a | 30,386.67 ± 1,412.62b | 32,000.00 ± 2,003.29b | 15.133 | 2 | 0.001 |
| 花粉体积 Pollen volume (mm3) | 1,213.07 ± 28.87a | 1,102.47 ± 31.08b | 1,067.13 ± 49.45b | 10.226 | 2 | 0.006 |
| 胚珠数 Ovule Number | 2.00 ± 0.00a | 2.00 ± 0.00a | 0.90 ± 0.35b | 46.620 | 2 | < 0.001 |
| 单花期 Flower longevity (days) | 10.43 ± 0.24a | 9.29 ± 0.56b | 10.70 ± 0.30a | 6.620 | 2 | 0.037 |
| 雄花期 Male phase (days) | 5.46 ± 0.35a | 5.23 ± 0.52a | / | 0.140 | 1 | 0.079 |
| 雌花期 Female phase (days) | 4.96 ± 0.30a | 4.06 ± 0.30b | / | 4.670 | 1 | 0.031 |
| 变量 Source of variation | Wald χ2 | df | P |
|---|---|---|---|
| 萌发率 Germination rates | |||
| 授粉处理 Pollination treatments | 1.010 | 2 | 0.604 |
| 花粉受体 Pollen recipient type | 4.110 | 1 | 0.043 |
| 交互作用 Interaction | 3.860 | 2 | 0.145 |
| 花粉管长度 Pollen tube length | |||
| 授粉处理 Pollination treatments | 147.178 | 2 | < 0.001 |
| 花粉受体 Pollen recipient type | 26.615 | 1 | < 0.001 |
| 交互作用 Interaction | 19.526 | 2 | < 0.001 |
表2 授粉处理(长柱型、短柱型和雄花型的花粉)时花粉受体(长柱型和短柱型分别为母本)及其交互作用对梅花柱头花粉的萌发率、花粉管长度的影响(广义线性模型)
Table 2 Effect of pollination treatments (long-style, short-style and male-flower types) and pollen recipient type (long-style vs short-style), and their interaction on pollen germination rates and pollen tube length in Armeniaca mume (generalized linear model)
| 变量 Source of variation | Wald χ2 | df | P |
|---|---|---|---|
| 萌发率 Germination rates | |||
| 授粉处理 Pollination treatments | 1.010 | 2 | 0.604 |
| 花粉受体 Pollen recipient type | 4.110 | 1 | 0.043 |
| 交互作用 Interaction | 3.860 | 2 | 0.145 |
| 花粉管长度 Pollen tube length | |||
| 授粉处理 Pollination treatments | 147.178 | 2 | < 0.001 |
| 花粉受体 Pollen recipient type | 26.615 | 1 | < 0.001 |
| 交互作用 Interaction | 19.526 | 2 | < 0.001 |
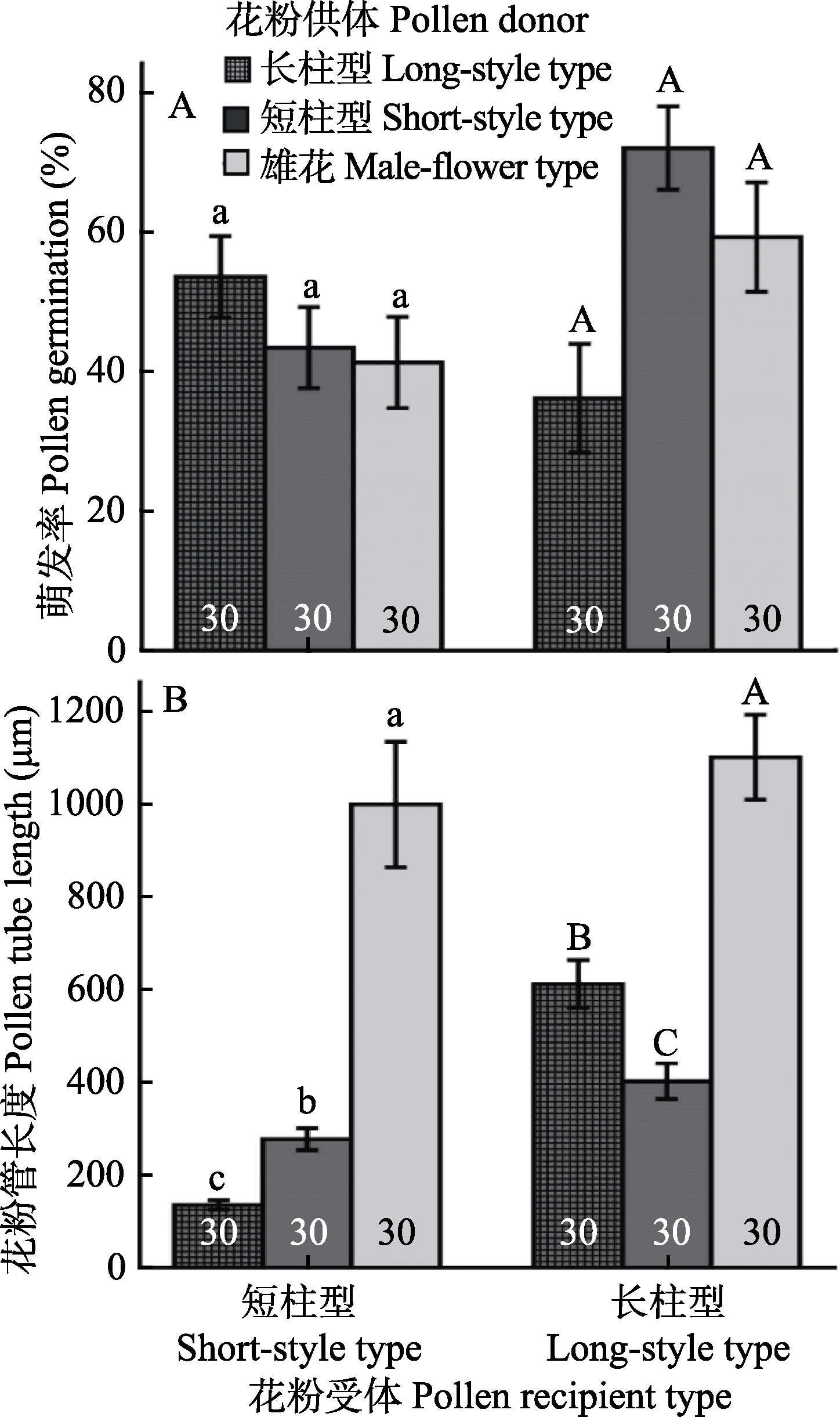
图2 不同处理间梅花的花粉萌发率(A)和花粉管长度(B)的比较(平均值±标准误)。长柱型花粉、短柱型花粉、雄花型花粉分别为花粉供体, 短柱型和长柱型梅花分别作为花粉受体。相同字母表示处理间没有显著差异。条形图内部的数字表示处理的样本量。
Fig. 2 Comparisons of in vivo pollen germination rates (A) and pollen tube length (mean ± SE) (B) of pollen donors from long-style, short-style and male-flower types, with the long- style and short-style types in Armeniaca mume as pollen recipients. The same letters indicate no significant difference within the flower type. Numbers of sample size are shown in the bars.
| 变量 Source of variation | Wald χ2 | df | P |
|---|---|---|---|
| 坐果率 Fruit set | |||
| 授粉处理 Pollination treatments | 0.704 | 3 | 0.872 |
| 花粉受体 Pollen recipient type | 3.914 | 1 | 0.048 |
| 交互作用 Interaction | 5.920 | 3 | 0.116 |
表3 授粉处理(自然对照、长柱型、短柱型和雄花型的花粉)时花粉受体(长柱型和短柱型分别为母本)及其交互作用对梅花坐果率的影响(广义线性模型)
Table 3 Effect of pollination treatments (control, long-style type, short-style type, male-flower type) and pollen recipient type (long-style vs short-style), and their interaction on fruit set of Armeniaca mume (generalized linear model)
| 变量 Source of variation | Wald χ2 | df | P |
|---|---|---|---|
| 坐果率 Fruit set | |||
| 授粉处理 Pollination treatments | 0.704 | 3 | 0.872 |
| 花粉受体 Pollen recipient type | 3.914 | 1 | 0.048 |
| 交互作用 Interaction | 5.920 | 3 | 0.116 |
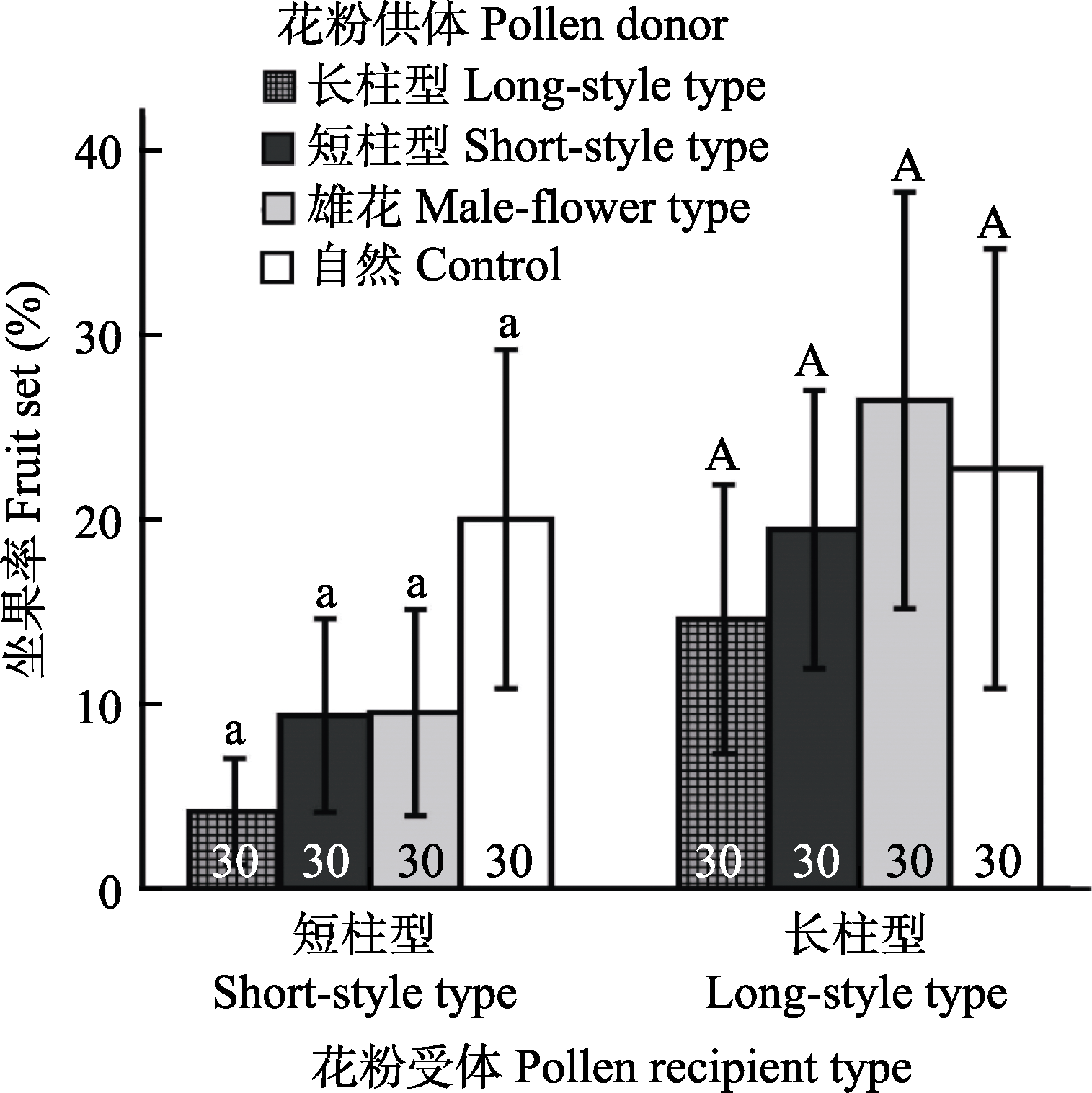
图3 不同处理梅花坐果率比较。花粉供体为长柱型、短柱型、雄花型花粉和自然对照, 短柱型和长柱型梅花分别作为花粉受体。相同字母表示处理间没有显著的差异。条形图内部的数字表示处理的样本量。
Fig. 3 Comparison of fruit set between pollen donors from long-style, short-style and male-flower types, with the long-style and short-style types in Armeniaca mume as pollen recipients. Bars with the same letters are not significantly different. Numbers of sample size are shown in the bars.
| [1] |
Barrett SCH, Hough J ( 2013) Sexual dimorphism in flowering plants. Journal of Experimental Botany, 64, 67-82.
DOI URL |
| [2] |
Barrett SCH, Harder LD ( 2005) The evolution of polymorphic sexual systems in daffodils (Narcissus). New Phytologist, 154, 45-53.
DOI URL PMID |
| [3] |
Bell G ( 1985) On the function of flowers. Proceedings of the Royal Society B: Biological Sciences, 224, 223-265.
DOI URL |
| [4] |
Bernasconi G, Lang DJ, Schmid B ( 2007) Microgametophyte population sizes and plant reproductive output in the insect- pollinated Prunella grandiflora (Lamiaceae). New Phytologist, 173, 393-400.
DOI URL PMID |
| [5] |
Campbell DR ( 1989) Measurements of selection in a hermaphroditic plant: Variation in male and female pollination success. Evolution, 43, 318-334.
DOI URL PMID |
| [6] |
Cresswell JE ( 2000) Manipulation of female architecture in flowers reveals a narrow optimum for pollen deposition. Ecology, 81, 3244-3249.
DOI URL |
| [7] | Chen JY, Bao MZ ( 1992) Botanical classification and horticultural classification of Chinese Mei (Prunus mume) resources. Journal of Zhejiang Forest College, 9, 119-132. (in Chinese with English abstract) |
| [ 陈俊愉, 包满珠 ( 1992) 中国梅(Prunus mume)的植物学分类和园艺学分类. 浙江林学院学报, 9, 119-132.] | |
| [8] |
Diggle PK ( 1995) Architectural effects and the interpretation of patterns of fruit and seed development. Annual Review of Ecology and Systematics, 26, 531-552.
DOI URL |
| [9] |
Diggle PK ( 1997) Ontogenetic contingency and floral morphology: The effects of architecture and resource limitation. International Journal of Plant Sciences, 158, 99-107.
DOI URL |
| [10] |
Erbar C ( 2003) Pollen tube transmitting tissue: Place of competition of male gametophytes. International Journal of Plant Sciences, 164, 265-277.
DOI URL |
| [11] |
Granado-Yela C, Balaguer L, Cayuela L, Méndez M ( 2017) Unusual positional effects on flower sex in an andromonoecious tree: Resource competition, architectural constraints, or inhibition by the apical flower? American Journal of Botany, 104, 608-615.
DOI URL PMID |
| [12] |
Hong Y, Fang HT, Na R ( 2006) Pollen viability and stigma receptivity of Prunus mongolica Maxim. Guihaia, 26, 589-591. (in Chinese with English abstract)
DOI URL |
|
[ 红雨, 方海涛, 那仁 ( 2006) 濒危植物蒙古扁桃花粉活力和柱头可授性研究. 广西植物, 26, 589-591.]
DOI URL |
|
| [13] |
Huang SQ, Guo YH ( 2000) New advances in pollination biology. Chinese Science Bulletin, 45, 225-237. (in Chinese)
DOI |
|
[ 黄双全, 郭友好 ( 2000) 传粉生物学的研究进展. 科学通报, 45, 225-237.]
DOI |
|
| [14] |
Johnson MTJ, Smith SD, Rausher MD ( 2010) Effects of plant sex on range distributions and allocation to reproduction. New Phytologist, 186, 769-779.
DOI URL PMID |
| [15] |
Jürgens A, Witt T, Gottsberger G ( 2012) Pollen grain size variation in Caryophylloideae: A mixed strategy for pollen deposition along styles with long stigmatic areas? Plant Systematics and Evolution, 298, 9-24.
DOI URL |
| [16] |
Lankinen ?, Skogsmyr I ( 2001) Evolution of pistil length as a choice mechanism for pollen quality. Oikos, 92, 81-90.
DOI URL |
| [17] |
Liu CQ, Huang SQ ( 2012) Does the relative importance of resource competition and architectural effect in floral variation vary with stages of floral ontogeny? Journal of Systematics and Evolution, 50, 119-124.
DOI URL |
| [18] |
Lloyd DG ( 1980) Sexual strategies in plants. I. An hypothesis of serial adjustment of maternal investment during one reproductive session. New Phytologist, 86, 69-79.
DOI URL |
| [19] | Love J, Graham SW, Irwin JA, Ashton PA, Bretagnolle F, Abbott RJ ( 2016) Self-pollination, style length development and seed set in self-compatible Asteraceae: Evidence from Senecio vulgaris L. Transactions of the Botanical Society of Edinburgh, 9, 371-379. |
| [20] | Lu LD, Bruce B ( 2003) Armeniaca. In: Flora of China (eds Wu ZY, Raven PH, Hong DY), Vol. 9, pp. 396-401. Science Press, Beijing & Missouri Botanical Garden Press St. Louis. |
| [21] |
Ma WJ, Zhang FP, Peng YQ, Yang DR ( 2009) Comparison of style length and reproduction success in Ficus of different breeding systems. Chinese Journal of Plant Ecology, 33, 911-918. (in Chinese with English abstract)
DOI URL |
|
[ 马文娟, 张凤萍, 彭艳琼, 杨大荣 ( 2009) 不同繁育系统的榕树雌花的花柱长度和繁殖率比较. 植物生态学报, 33, 911-918.]
DOI URL |
|
| [22] |
Mazer SJ, Dudley LS, Hove AA, Emms SK, Verhoeven AS ( 2010) Physiological performance in Clarkia sister taxa with contrasting mating systems: Do early-flowering autogamous taxa avoid water stress relative to their pollinator-dependent counterparts? International Journal of Plant Sciences, 171, 1029-1047.
DOI URL |
| [23] |
Mazer SJ, Moghaddasi A, Bello AK, Hove AA ( 2016) Winning in style: Longer styles receive more pollen, but style length does not affect pollen attrition in wild Clarkia populations. American Journal of Botany, 103, 408-422.
DOI URL PMID |
| [24] |
Mulcahy DL ( 1979) The rise of the angiosperms: A genecological factor. Science, 206, 20-23.
DOI URL |
| [25] |
Plitmann U ( 1993) Pollen tube attrition as related to breeding systems in Brassicaceae. Plant Systematics and Evolution, 188, 65-72.
DOI URL |
| [26] |
Ramesha BT, Yetish MD, Ravikanth G, Ganeshaiah KN, Ghazoul J, Shaanker RU ( 2011) Stylish lengths: Mate choice in flowers. Journal of Biosciences, 36, 229-234.
DOI URL PMID |
| [27] | Roulston TH, Cane JH, Buchmann SL ( 2000) What governs protein content of pollen: Pollinator preferences, pollen- pistil interactions, or phylogeny? Ecological Monographs, 70, 617-643. |
| [28] |
Stephenson AG, Travers SE, Mena-Ali JI, Winsor JA ( 2003) Pollen performance before and during the autotrophic- heterotrophic transition of pollen tube growth. Philosophical Transactions of the Royal Society of London B: Biological Sciences, 358, 1009-1018.
DOI URL |
| [29] |
Taylor ML, Williams JH ( 2012) Pollen tube development in two species of Trithuria (Hydatellaceae) with contrasting breeding systems. Sexual Plant Reproduction, 25, 83-96.
DOI URL PMID |
| [30] | Ushimaru A, Itagaki T, Ishii HS ( 2003) Variation in floral organ size depends on function. Evolutionary Ecology Research, 5, 615-622. |
| [31] |
Wang XF, Armbruster WS, Huang SQ ( 2012) Extra-gynoecial pollen-tube growth in apocarpous angiosperms is phylogenetically widespread and probably adaptive. New Phytologist, 193, 253-260.
DOI URL |
| [32] |
Wang XY, Quan QM, Wang B, Li YX, Huang SQ ( 2018) Pollen competition between morphs in a pollen-color dimorphic herb and the loss of phenotypic polymorphism within populations. Evolution, 72, 785-797.
DOI URL PMID |
| [33] |
Wang YF, Gao HY, Shi HY, Wang JH, Du GZ ( 2008) Adaptive significance of Saussurea parviflora's sexual organs, Qinghai-Tibetan Plateau, China. Journal of Plant Ecology (Chinese Version), 32, 379-384. (in Chinese with English abstract)
DOI URL |
|
[ 王一峰, 高宏岩, 施海燕, 王剑虹, 杜国祯 ( 2008) 小花风毛菊的性器官在青藏高原的海拔变异. 植物生态学报, 32, 379-384.]
DOI URL |
|
| [34] |
Zhang CX, Tateishi N, Tanabe K ( 2010) Pollen density on the stigma affects endogenous gibberellin metabolism, seed and fruit set, and fruit quality in Pyrus pyrifolia. Journal of Experimental Botany, 61, 4291-4302.
DOI URL |
| [1] | 刘俊, 张天祥, 张壹萱, 黄晓凤, 韩卫杰, 吴问国, 黄晓飞, 黄理政, 张阳. 基于DNA宏条形码的江西桃红岭梅花鹿食性与营养策略[J]. 生物多样性, 2026, 34(2): 25214-. |
| [2] | 葛湘湘, 徐玉洁, 汤菊. 朱砂梅花朝向变异影响传粉者访花行为和传粉效率[J]. 生物多样性, 2025, 33(9): 25221-. |
| [3] | 麦晓烔, 康佳, 李梓琛, 王天明. 东北虎豹国家公园梅花鹿活动节律及其对道路的响应[J]. 生物多样性, 2024, 32(11): 24178-. |
| [4] | 热依拉穆·麦麦提吐尔逊, 艾沙江·阿不都沙拉木. 石榴花瓣和雄蕊对其传粉过程与繁殖成功的影响[J]. 生物多样性, 2023, 31(7): 22633-. |
| [5] | 李永光, 任辉, 张英杰, 李瑞宁, 艾昊, 黄先忠. 十字花科植物PEBP基因家族的分子进化[J]. 生物多样性, 2022, 30(6): 21545-. |
| [6] | 胡德美, 姚仁秀, 陈燕, 游贤松, 王顺雨, 汤晓辛, 王晓月. 青篱柴通过促进亲和花粉生长而提高传粉精确性[J]. 生物多样性, 2021, 29(7): 887-896. |
| [7] | 王世彤, 徐耀粘, 杨腾, 魏新增, 江明喜. 微生境对黄梅秤锤树野生种群叶片功能性状的影响[J]. 生物多样性, 2020, 28(3): 277-288. |
| [8] | 李晶, 周天阳, 鲁雪丽, 李新涛, 孙斌, 孟红杰. 珍稀植物连香树在其中国分布区北缘的种子性状及幼苗更新限制[J]. 生物多样性, 2020, 28(10): 1161-1173. |
| [9] | 王世彤, 吴浩, 刘梦婷, 张佳鑫, 刘检明, 孟红杰, 徐耀粘, 乔秀娟, 魏新增, 卢志军, 江明喜. 极小种群野生植物黄梅秤锤树群落结构与动态[J]. 生物多样性, 2018, 26(7): 749-759. |
| [10] | 黄至欢, 陆奇丰, 陈颖卓. 地锦苗在石灰岩土壤和红壤生境中的繁殖成功的比较[J]. 生物多样性, 2017, 25(9): 972-980. |
| [11] | 杜巍, 王帅, 王满囷, 汪小凡. 谁是腊梅的主要传粉者: 昆虫行为与传粉作用[J]. 生物多样性, 2012, 20(3): 400-404. |
| [12] | 周世良, 叶文国. 夏腊梅的遗传多样性及其保护[J]. 生物多样性, 2002, 10(1): 1-6. |
| [13] | 徐宏发, 陆厚基, 盛和林, 顾长明. 华南梅花鹿的分布和现状[J]. 生物多样性, 1998, 06(2): 87-91. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
备案号:京ICP备16067583号-7
Copyright © 2026 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn
![]()