
|
|
||
|
第三极荨麻属麻叶荨麻分支的物种界限
生物多样性
2025, 33 (8):
25138-.
DOI: 10.17520/biods.2025138
基于多学科证据的整合分类学是当前解决物种界限问题的有效方法。荨麻属(Urtica)麻叶荨麻分支(cannabina Clade)广泛分布于生物多样性丰富且生态环境独特的第三极地区。然而, 由于该属植物复杂的形态特征和演化历史, 其近缘种之间的界限仍不清楚。本研究以分布于该地区的麻叶荨麻分支(含亚种)的4个物种(麻叶荨麻Urtica cannabina、三角叶荨麻U. triangularis、异株荨麻U. dioica、高原荨麻U. hyperborea)为研究对象, 选取滇藏荨麻(U. mairei)和膜叶荨麻(U. membranifolia)及近缘属为外类群, 基于麻叶荨麻分支50个个体(每个种4-10个)及18个外类群样本的基因组重测序数据, 提取质体基因组和“Angiosperms353核基因数据集”解析系统发育关系和群体结构, 并结合地理分布以及形态性状聚类分析, 开展了整合分类学研究。结果表明: (1)高原荨麻是单系类群, 且在形态特征和地理分布上与其他物种分化明显, 尽管与异株荨麻在喜马拉雅和横断山地区存在同域分布, 但二者在海拔梯度上存在分异; (2)异株荨麻的原亚种与尾尖异株荨麻(Urtica dioica subsp. afghanica)和甘肃异株荨麻(U. dioica subsp. gansuensis)两个亚种在地理分布、形态特征及遗传组成方面均存在明显不同, 暗示可能存在隐存种, 其分类地位有待进一步验证; (3)三角叶荨麻、麻叶荨麻和异株荨麻等近缘物种的分布区重叠, 性状和遗传聚类出现交叉, 同时检测到核质冲突信号, 表明种间可能存在基因渐渗。高山环境中物种的形态趋同可能是物种间性状难以区分的原因。此外, 研究证实质体基因组结合“Angiosperms353核基因数据集”在近缘物种界定中有较好的应用潜力。本研究利用多重证据解析了该地区荨麻属植物的物种界限, 加深了对其种间关系与分布格局的理解, 并为该区域内其他类群的整合分类学研究提供了一个可借鉴的范式。 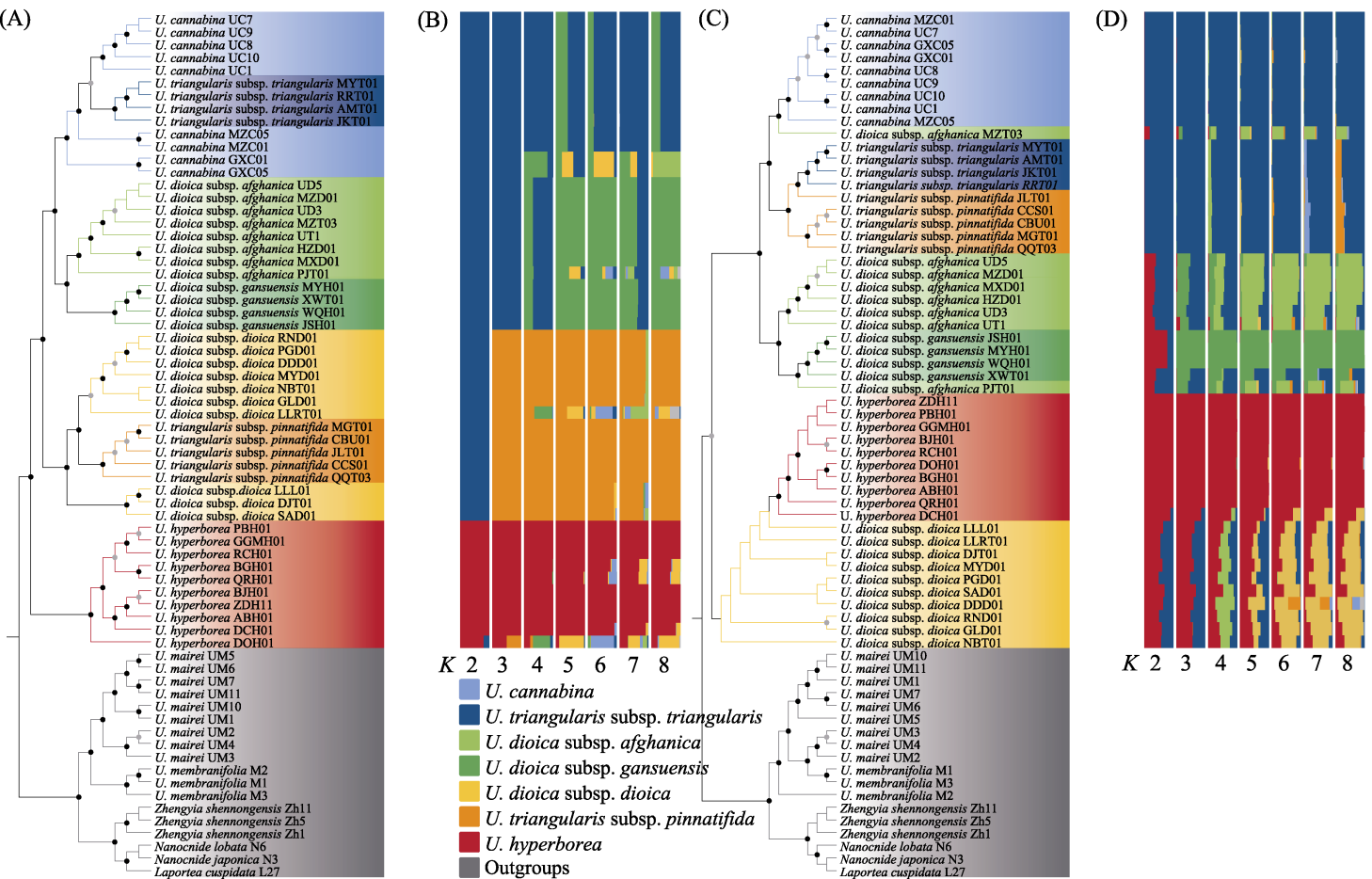 View image in article
图4
基于质体基因组和Angiosperms353核基因数据集的麻叶荨麻分支的系统发育关系和遗传结构。系统发育树节点处黑色圆点表示自展值大于90%, 灰色圆点表示自展值为70%-90%; 不同的背景颜色对应不同的物种。(A)基于最大似然法推断的质体的系统发育树; (B)基于质体基因组数据的4种荨麻属植物的Structure分析结果(最佳K = 5); (C)基于最大似然法推断的Angiosperms353核基因数据集的系统发育树; (D)基于Angiosperms353核基因数据集的4种荨麻属植物的Structure分析结果(最佳K = 3)。U. cannabina: 麻叶荨麻; U. triangularis subsp. triangularis: 三角叶荨麻原亚种; U. dioica subsp. afghanica: 尾尖异株荨麻; U. dioica subsp. gansuensis: 甘肃异株荨麻; U. dioica subsp. dioica: 异株荨麻原亚种; U. triangularis subsp. pinnatifida: 羽裂荨麻; U. hyperborea: 高原荨麻; Outgroups: 外类群。
正文中引用本图/表的段落
(4)遗传结构。基于质体的系统发育树表明(图4A), 高原荨麻是单系类群, 其姐妹类群是异株荨麻原亚种和羽裂荨麻; 尾尖异株荨麻和甘肃异株荨麻未与异株荨麻原亚种聚类; 异株荨麻原亚种为多系; 羽裂荨麻未与三角叶荨麻原亚种聚类, 而三角叶荨麻原亚种与麻叶荨麻互为姐妹群, 麻叶荨麻为多系。基于质体基因组数据的Structure分析结果显示(图4B), 高原荨麻及6个近缘种可以分为4个遗传谱系(最佳K值为5, 见附录8A), 高原荨麻为一个谱系, 异株荨麻原亚种和羽裂荨麻为一个谱系, 尾尖异株荨麻和甘肃异株荨麻为一个谱系, 麻叶荨麻和三角叶荨麻原亚种为一个谱系。
基于291个Angiosperms353核基因串联数据集的系统发育树(图4C)同样表明高原荨麻为单系, 与异株荨麻原亚种是姐妹群, 异株荨麻原亚种为并系。同时, 尾尖异株荨麻和甘肃异株荨麻未与异株荨麻原亚种聚类, 而羽裂荨麻则与三角叶荨麻原亚种互为姐妹群, 与麻叶荨麻亲缘关系最近。核基因串联数据集推断的ML树与ASTRAL-III物种树拓扑结构基本一致(附录9), 然而羽裂荨麻的系统发育位置存在核质冲突。此外, 异株荨麻原亚种在核基因中表现为并系, 而麻叶荨麻则为单系。Structure分析结果显示(图4D), 4个物种可以分为3个遗传谱系(最佳K值为3, 见附录8B): 高原荨麻和异株荨麻原亚种为一个谱系, 且高原荨麻的遗传组分高度一致; 麻叶荨麻和三角叶荨麻(包括羽裂荨麻)为另一类。分析还显示, 甘肃异株荨麻和尾尖异株荨麻谱系分化不明显, 尾尖异株荨麻的个体MZT03、PJT01与近缘种之间可能存在遗传组分的混合, 暗示其与近缘种之间可能存在基因流。
在麻叶荨麻分支中, 高原荨麻在核基因与质体系统发育树中均为单系, 与采集自第三极的异株荨麻原亚种(异名为西藏荨麻)近缘(Grosse-Veldmann et al, 2016; Huang et al, 2019; Ogoma et al, 2022), 且其性状和地理分布也与其他物种明显不同, 支持其为一个较好的单系物种。异株荨麻的物种界限较为复杂, 本研究包含异株荨麻的3个亚种: 异株荨麻原亚种、甘肃异株荨麻及尾尖异株荨麻, 但在系统树中三者并未聚成单系(图4A-D), 形态上亦难以区分(图1, 图3), 可能代表不同的隐存种。其中甘肃异株荨麻及尾尖异株荨麻在质体基因组和核基因组都是单系(图4), 且存在显著的遗传分化(图5, 附录10, 附录11), 因此可能代表了2个姐妹隐存种。此外, 尾尖异株荨麻的个体PJT01在核质数据中系统位置不一致, 提示存在基因渐渗的可能(图4)。此前研究显示, 第三极地区与欧洲地区的异株荨麻聚在完全不同的分支(Grosse-Veldmann et al, 2016), 加之异株荨麻多样的形态特征(Chen et al 2003; Grosse- Veldmann & Weigend, 2015; Wu et al 2015), 进一步支持异株荨麻复合群可能包含多个完全不同的物种。
本文的其它图/表
|

{kind=link}