
|
|
||
|
第三极荨麻属麻叶荨麻分支的物种界限
生物多样性
2025, 33 (8):
25138-.
DOI: 10.17520/biods.2025138
基于多学科证据的整合分类学是当前解决物种界限问题的有效方法。荨麻属(Urtica)麻叶荨麻分支(cannabina Clade)广泛分布于生物多样性丰富且生态环境独特的第三极地区。然而, 由于该属植物复杂的形态特征和演化历史, 其近缘种之间的界限仍不清楚。本研究以分布于该地区的麻叶荨麻分支(含亚种)的4个物种(麻叶荨麻Urtica cannabina、三角叶荨麻U. triangularis、异株荨麻U. dioica、高原荨麻U. hyperborea)为研究对象, 选取滇藏荨麻(U. mairei)和膜叶荨麻(U. membranifolia)及近缘属为外类群, 基于麻叶荨麻分支50个个体(每个种4-10个)及18个外类群样本的基因组重测序数据, 提取质体基因组和“Angiosperms353核基因数据集”解析系统发育关系和群体结构, 并结合地理分布以及形态性状聚类分析, 开展了整合分类学研究。结果表明: (1)高原荨麻是单系类群, 且在形态特征和地理分布上与其他物种分化明显, 尽管与异株荨麻在喜马拉雅和横断山地区存在同域分布, 但二者在海拔梯度上存在分异; (2)异株荨麻的原亚种与尾尖异株荨麻(Urtica dioica subsp. afghanica)和甘肃异株荨麻(U. dioica subsp. gansuensis)两个亚种在地理分布、形态特征及遗传组成方面均存在明显不同, 暗示可能存在隐存种, 其分类地位有待进一步验证; (3)三角叶荨麻、麻叶荨麻和异株荨麻等近缘物种的分布区重叠, 性状和遗传聚类出现交叉, 同时检测到核质冲突信号, 表明种间可能存在基因渐渗。高山环境中物种的形态趋同可能是物种间性状难以区分的原因。此外, 研究证实质体基因组结合“Angiosperms353核基因数据集”在近缘物种界定中有较好的应用潜力。本研究利用多重证据解析了该地区荨麻属植物的物种界限, 加深了对其种间关系与分布格局的理解, 并为该区域内其他类群的整合分类学研究提供了一个可借鉴的范式。  View image in article
图1
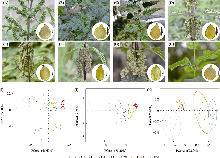
第三极6种荨麻属植物的植株形态及生境。其中(A)-(G)为麻叶荨麻分支的物种, (H)-(I)为荨麻属内的外类群。(A)麻叶荨麻; (B)羽裂荨麻; (C)三角叶荨麻原亚种; (D)尾尖异株荨麻; (E)甘肃异株荨麻; (F)异株荨麻原亚种; (G)高原荨麻; (H)滇藏荨麻; (I)膜叶荨麻标本照(来自中国科学院昆明植物研究所标本馆)。
正文中引用本图/表的段落
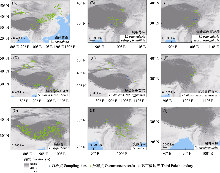
荨麻属(Urtica)隶属于荨麻科, 是全球广布属, 约30种, 其中我国分布有14种, 包括3个特有种(Friis, 1993; Chen et al, 2003)。该属物种形态相似(图1), 表型可塑性较强(Wu et al, 2015), 尤其是同域分布的近缘物种及亚种之间, 种间界限仍不清楚(Taylor, 2009; Henning et al, 2014; Grosse-Veldmann & Weigend, 2015; Grosse-Veldmann et al, 2016)。该属经历了多次不同生物地理区间的扩散事件(Wu et al, 2018), 可能打破了物种的地理与亲缘关系的界限, 使得仅依赖形态特征和地理分布数据很难清晰界定种间关系(Grosse-Veldmann et al, 2016)。近年来, 荨麻属及荨麻科的分子系统学研究不断推进。基于常规DNA片段和质体基因组的分析表明荨麻属是一个单系类群(包含西海麻属Hesperocnide), 其姊妹群为征镒麻(Zhengyia shennongensis) (Wu et al, 2013, 2018; Huang et al, 2019; Ogoma et al, 2022)。此外, 基于荨麻属系统发育分析显示, 该属包含多个支持率较高的演化分支, 在安第斯山、新西兰和欧亚大陆北部存在3个物种辐射中心, 但一些物种的划分仍存在问题(Grosse-Veldmann et al, 2016)。在近缘种层面, 有研究关注了异株荨麻(Urtica dioica)、咬人荨麻(U. thunbergiana)的物种界限和分类问题(Henning et al, 2014; Becker et al, 2017a), 但麻叶荨麻分支(cannabina Clade)尚无研究。该分支包括高原荨麻(U. hyperborea)、三角叶荨麻(U. triangularis)、异株荨麻(U. dioica)和麻叶荨麻(U. cannabina), 构成一个单系群(Grosse-Veldmann et al, 2016; Ogoma et al, 2022), 主要分布在第三极及周边地区(图2)。尽管该分支具有清晰的系统发育位置, 但其内部包含多个亚种, 且彼此间同域或邻域分布, 可能进一步模糊了物种的边界。此外, 现有研究主要集中于麻叶荨麻分支内较为常见的物种, 而对整个麻叶荨麻分支的种间界限和分类问题的系统研究仍然不足。例如, 在《中国植物志》中, 异株荨麻包含4个亚种(异株荨麻原亚种U. dioica subsp. dioica、尾尖异株荨麻U. dioica subsp. afghanica、新疆异株荨麻U. dioica subsp. xingjiangensis和甘肃异株荨麻U. dioica subsp. gansuensis), 西藏荨麻(U. tibetica)被列为独立的物种(王文采和陈家瑞, 1995); 而在Flora of China中, 异株荨麻仅包含3个亚种(异株荨麻原亚种、尾尖异株荨麻和甘肃异株荨麻), 其中新疆异株荨麻被视为尾尖异株荨麻的异名, 而西藏荨麻则被视为异株荨麻原亚种的异名(Chen et al, 2003)。因此人们对这些物种的认识仍停留在形态描述与记录阶段, 亟需进一步结合分子及其他证据厘清麻叶荨麻分支的物种界限(Henning et al, 2014; Becker et al, 2017b; Huang et al, 2019)。
第三极(the Third Pole)是指青藏高原、横断山脉、喜马拉雅山脉、兴都库什山脉和喀喇昆仑山脉等组成的区域(Qiu, 2008; Liu J et al, 2022)。区域内包括了中国西南山地、印缅地区、喜马拉雅和中亚山地4个全球生物多样性热点地区(Myers et al, 2000; Liu J et al, 2022)。尽管该地区有较长的植物分类学研究历史, 但由于区域内复杂的气候、地质和地理环境, 加之地缘政治冲突, 目前尚不知确切的植物物种多样性(Ahmad et al, 2025; Yang et al, 2025), 初步估计约有18,000多种维管植物(Liu J et al, 2022), 亟需对不同类群开展深入研究。我国荨麻属植物约有10种在第三极及其周边区域广泛分布, 这包括麻叶荨麻分支以及滇藏荨麻(U. mairei)和膜叶荨麻(U. membranifolia)等(图1), 它们主要分布于西藏、青海以及新疆南部、甘肃南部、四川西部、云南西北部等地(图2)。这些荨麻属植物是重要的药食同源植物, 具有丰富的民俗文化和经济价值(帝玛尔·丹增彭措, 1986; Chen et al, 2003; Viotti et al, 2022; Wang et al, 2023)。例如, 在第三极广泛分布的高原荨麻和异株荨麻原亚种, 在藏语里称为“sabo”, 相传是藏地著名密教修行者米拉日巴大师在山洞修行时的主要食物, 为纪念米拉日巴大师, 当地牧民至今仍保留着每年春天食用高原荨麻嫩叶的传统。高原荨麻还可作为冬季重要的食饲来源, 或研磨成粉掺到青稞面或青稞饼中供牲畜或人类食用。此外, 三角叶荨麻、滇藏荨麻等物种, 同样被当地人们作为重要的药、食、饲来源(中国科学院西北高原生物研究所, 1991; 苏日娜等, 2018)。这些植物在第三极生态系统中扮演着重要角色, 既与人们生计相关, 又与精神信仰和医药价值密切关联。然而, 高原荨麻、异株荨麻、三角叶荨麻等近缘种在形态上十分相似(图1), 地理分布有所重叠(图2), 因此其在形态、地理和遗传上有何不同需要进一步澄清。
通过搜索全球生物多样性信息库(Global Biodiversity Information Facility, GBIF) (
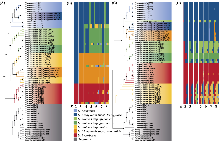
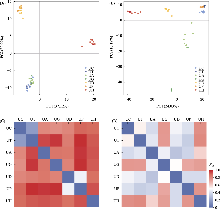
在麻叶荨麻分支中, 高原荨麻在核基因与质体系统发育树中均为单系, 与采集自第三极的异株荨麻原亚种(异名为西藏荨麻)近缘(Grosse-Veldmann et al, 2016; Huang et al, 2019; Ogoma et al, 2022), 且其性状和地理分布也与其他物种明显不同, 支持其为一个较好的单系物种。异株荨麻的物种界限较为复杂, 本研究包含异株荨麻的3个亚种: 异株荨麻原亚种、甘肃异株荨麻及尾尖异株荨麻, 但在系统树中三者并未聚成单系(图4A-D), 形态上亦难以区分(图1, 图3), 可能代表不同的隐存种。其中甘肃异株荨麻及尾尖异株荨麻在质体基因组和核基因组都是单系(图4), 且存在显著的遗传分化(图5, 附录10, 附录11), 因此可能代表了2个姐妹隐存种。此外, 尾尖异株荨麻的个体PJT01在核质数据中系统位置不一致, 提示存在基因渐渗的可能(图4)。此前研究显示, 第三极地区与欧洲地区的异株荨麻聚在完全不同的分支(Grosse-Veldmann et al, 2016), 加之异株荨麻多样的形态特征(Chen et al 2003; Grosse- Veldmann & Weigend, 2015; Wu et al 2015), 进一步支持异株荨麻复合群可能包含多个完全不同的物种。
形态特征缺乏明显差异在本研究中尤为突出, 麻叶荨麻分支中的多个物种, 包括高原荨麻和异株荨麻原亚种, 在适应性状上表现出高度相似性, 这很可能是高山环境的选择压力导致不同物种在形态特征上趋同演化的结果(Wu SD et al, 2022)。本研究观察到的荨麻属植物刺毛密集、叶片狭长和花被片增大等特征(图1, 附录2), 与其他高山植物如苞叶雪莲(Saussurea obvallata)和喜山葶苈(Draba oreades)的适应性状很相似, 这些物种普遍呈植株低矮丛生或垫状生长、叶窄披针或微型化, 叶面覆厚蜡质层与密集绒刺以抵御强风与高紫外线(Zhang et al, 2023; Zhang et al, 2024)。这种形态趋同演化现象的背后可能蕴含复杂的调控机制, 相关的代谢物差异研究表明, 同域分布的高原荨麻与异株荨麻原亚种在适应不同海拔环境时, 表现出不同的适应机制(Deng et al, 2025)。此外, Zhou等(2024)在高山栎组(Quercus sect. Heterobalanus)的研究中也发现, 植物会通过调节碳、氮、磷元素的含量和利用效率以及独特的叶结构来适应高山环境。但上述趋同演化的分子基础, 还需要未来深入的功能基因组学及其与表型和环境的耦合研究。
本文的其它图/表
|
{kind=link}