|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
第三极荨麻属麻叶荨麻分支的物种界限
生物多样性
2025, 33 (8):
25138-.
DOI: 10.17520/biods.2025138
基于多学科证据的整合分类学是当前解决物种界限问题的有效方法。荨麻属(Urtica)麻叶荨麻分支(cannabina Clade)广泛分布于生物多样性丰富且生态环境独特的第三极地区。然而, 由于该属植物复杂的形态特征和演化历史, 其近缘种之间的界限仍不清楚。本研究以分布于该地区的麻叶荨麻分支(含亚种)的4个物种(麻叶荨麻Urtica cannabina、三角叶荨麻U. triangularis、异株荨麻U. dioica、高原荨麻U. hyperborea)为研究对象, 选取滇藏荨麻(U. mairei)和膜叶荨麻(U. membranifolia)及近缘属为外类群, 基于麻叶荨麻分支50个个体(每个种4-10个)及18个外类群样本的基因组重测序数据, 提取质体基因组和“Angiosperms353核基因数据集”解析系统发育关系和群体结构, 并结合地理分布以及形态性状聚类分析, 开展了整合分类学研究。结果表明: (1)高原荨麻是单系类群, 且在形态特征和地理分布上与其他物种分化明显, 尽管与异株荨麻在喜马拉雅和横断山地区存在同域分布, 但二者在海拔梯度上存在分异; (2)异株荨麻的原亚种与尾尖异株荨麻(Urtica dioica subsp. afghanica)和甘肃异株荨麻(U. dioica subsp. gansuensis)两个亚种在地理分布、形态特征及遗传组成方面均存在明显不同, 暗示可能存在隐存种, 其分类地位有待进一步验证; (3)三角叶荨麻、麻叶荨麻和异株荨麻等近缘物种的分布区重叠, 性状和遗传聚类出现交叉, 同时检测到核质冲突信号, 表明种间可能存在基因渐渗。高山环境中物种的形态趋同可能是物种间性状难以区分的原因。此外, 研究证实质体基因组结合“Angiosperms353核基因数据集”在近缘物种界定中有较好的应用潜力。本研究利用多重证据解析了该地区荨麻属植物的物种界限, 加深了对其种间关系与分布格局的理解, 并为该区域内其他类群的整合分类学研究提供了一个可借鉴的范式。
表1
第三极6种荨麻属植物物种水平上的基因组特征和遗传多样性。麻叶荨麻分支有4种, 荨麻属内的外类群有2种(加粗显示)。
正文中引用本图/表的段落
基于多学科证据的整合分类学是当前解决物种界限问题的有效方法。荨麻属(Urtica)麻叶荨麻分支(cannabina Clade)广泛分布于生物多样性丰富且生态环境独特的第三极地区。然而, 由于该属植物复杂的形态特征和演化历史, 其近缘种之间的界限仍不清楚。本研究以分布于该地区的麻叶荨麻分支(含亚种)的4个物种(麻叶荨麻Urtica cannabina、三角叶荨麻U. triangularis、异株荨麻U. dioica、高原荨麻U. hyperborea)为研究对象, 选取滇藏荨麻(U. mairei)和膜叶荨麻(U. membranifolia)及近缘属为外类群, 基于麻叶荨麻分支50个个体(每个种4-10个)及18个外类群样本的基因组重测序数据, 提取质体基因组和“Angiosperms353核基因数据集”解析系统发育关系和群体结构, 并结合地理分布以及形态性状聚类分析, 开展了整合分类学研究。结果表明: (1)高原荨麻是单系类群, 且在形态特征和地理分布上与其他物种分化明显, 尽管与异株荨麻在喜马拉雅和横断山地区存在同域分布, 但二者在海拔梯度上存在分异; (2)异株荨麻的原亚种与尾尖异株荨麻(Urtica dioica subsp. afghanica)和甘肃异株荨麻(U. dioica subsp. gansuensis)两个亚种在地理分布、形态特征及遗传组成方面均存在明显不同, 暗示可能存在隐存种, 其分类地位有待进一步验证; (3)三角叶荨麻、麻叶荨麻和异株荨麻等近缘物种的分布区重叠, 性状和遗传聚类出现交叉, 同时检测到核质冲突信号, 表明种间可能存在基因渐渗。高山环境中物种的形态趋同可能是物种间性状难以区分的原因。此外, 研究证实质体基因组结合“Angiosperms353核基因数据集”在近缘物种界定中有较好的应用潜力。本研究利用多重证据解析了该地区荨麻属植物的物种界限, 加深了对其种间关系与分布格局的理解, 并为该区域内其他类群的整合分类学研究提供了一个可借鉴的范式。
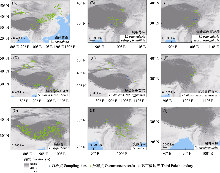
通过搜索全球生物多样性信息库(Global Biodiversity Information Facility, GBIF) (
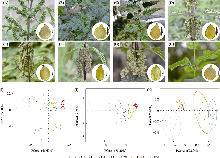
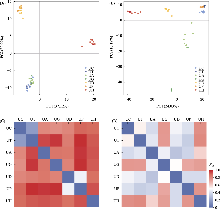
基于Flora of China (Chen et al, 2003)的描述, 选择16个重要形态性状进行测量, 包括10个质量性状(叶形、叶基、叶缘、叶尖、二级脉对数、基出脉伸达部位、托叶状态、刺毛密度、瘦果表面、退化雌蕊)和6个数量性状(叶长、叶宽、叶面积、叶柄长、花序长、花序梗长) (附录2, 附录3, 附录4)。对于数量性状, 选择成熟植株标本上的1-3片叶片(从顶部向下的第5-7对叶片), 使用0-150 mm电子数显游标卡尺(中国桂林, 桂林量具刃具有限责任公司)进行测量。叶面积的计算首先在拍摄叶片图像时添加10 cm的标尺作为参照物; 其次将带有标尺的图像导入ImageJ软件(Abràmoff et al, 2004), 使用“set scale”功能按照实际长度设置比例尺, 将单位转换为实际长度单位cm; 最后使用“set measurements”功能计算叶面积。质量性状参照Harris的标准进行统计(Harris & Harris, 2001)。我们通过体视显微镜观察种子表面特征后拍照记录, 并对其进行编码, 即种子表面光滑编码为1, 其他带疣点或细洼点编码为2。观察植株中部茎上的刺毛, 并对2 cm长的茎段一侧的刺毛计数并拍照记录, 因人工计数易产生误差, 因此将刺毛密度分为“密生” “稍密” “疏生” “几无” 4个等级, 其他质量性状编码规则见附录5。对所得性状, 利用主坐标分析(principal coordinate analysis, PCoA)对变量进行降维, 计算性状距离并可视化性状的分布和聚类情况。
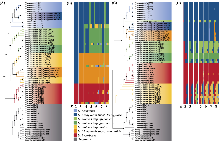
(1)质体基因组结构特征。本研究共组装了62个个体的质体基因组, 包含麻叶荨麻分支中4个物种(包含5个亚种)的50个个体、滇藏荨麻的9个个体以及膜叶荨麻的3个个体。其中三角叶荨麻和麻叶荨麻的质体基因组属于首次报道。从每个物种中选择1个个体进行质体基因组结构特征分析, 结果表明, 荨麻属植物的质体基因组均具有典型四分体结构, 包含2个反向重复区(inverted repeats, IRa和IRb)、1个大单拷贝区(large single copy, LSC)和1个小单拷贝区(small single copy, SSC) (附录6)。总GC含量为36.54%-36.80%。6个物种的tRNA、rRNA的数目完全一致, 而在蛋白质编码基因方面, 异株荨麻原亚种、尾尖异株荨麻和甘肃异株荨麻有83个, 其余物种为82个(附录7)。各物种的质体基因组大小在146,732 bp (U. mairei)至158,078 bp (U. membranifolia)之间, 相差11,346 bp (表1), 其中, 膜叶荨麻的基因组变异最大(146,785-158,078 bp), 可能反映了该物种内基因组结构的变异, 而高原荨麻和异株荨麻原亚种等物种的基因组长度则相对稳定。
(2) Angiosperms353核基因数据集特征。质控后, 共保留291个Angiosperms353核基因。其中, 尾尖异株荨麻是保留基因数量最多的物种, 有250个基因被保留; 其次是麻叶荨麻(有244个基因)、三角叶荨麻原亚种(有227个基因); 高原荨麻保留了190个基因; 外类群中, 滇藏荨麻和膜叶荨麻由于与麻叶荨麻分支遗传距离较远, 保留的基因数量最少(表1)。
(3)遗传多样性。计算了第三极6种荨麻属植物质体基因组长度(GL)、保留核基因数(NG)、单倍型总数(Nh)、单倍型多样性(Hd)和核苷酸多样性(π) (表1)。结果显示, 麻叶荨麻、尾尖异株荨麻和滇藏荨麻质体基因组的单倍型多样性低于核基因数据集; 核基因数据的单倍型多样性均为1.00。在质体基因组层面上, 滇藏荨麻(π = 1.15 × 10?3)和膜叶荨麻(π = 1.16 × 10?3)具有较高遗传多样性; 在低拷贝直系同源核基因数据集中, 膜叶荨麻的遗传多样性(π = 3.59 × 10?4)高于其他物种, 麻叶荨麻、三角叶荨麻原亚种和羽裂荨麻的遗传多样性水平相近, 但低于其他物种。
本文的其它图/表
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||