
|
|
||
|
被子植物隐性雌雄异株性系统的多样性、系统演化及进化意义
生物多样性
2024, 32 (6):
24149-.
DOI: 10.17520/biods.2024149
隐性雌雄异株指植株表型为非雌雄异株, 但形态上两性花通常雄性不育或雌性不育, 植株个体实际仅行使雌性或雄性功能。两性花植株向雌雄异株的转变在被子植物中广泛存在, 隐性雌雄异株作为潜在的中间阶段对于理解植物性系统的演化具有重要意义。据APG IV分类系统, 这些物种在被子植物的22目36科65属约221种植物中有过报道, 分别约占被子植物总目数的34.4%、总科数的8.7%、总属数的0.5%和总种数的0.1%; 多为分布于热带或亚热带地区依靠生物传粉的木本植物, 且至少存在花蜜或花粉作为访花报酬。该性系统形态学的性表达类型多样: 可分为雄花两性花异株(类型I)、两性花植株(类型II)、雌花两性花异株(类型III)及其他类型(类型IV) 4种, 分别约占已报道隐性雌雄异株物种数的48.9%、47.5%、2.7%和0.9%。从系统发育看, 该性系统在被子植物木兰分支、单子叶植物分支及真双子叶植物分支均有分布, 且主要分布于较为进化的核心真双子叶植物类群中。隐性雌雄异株物种不育性器官的进化意义存在多种假说, 包括祖先假说、遗传约束假说、促进风媒花粉落置假说、传粉者吸引假说、拟态与欺骗假说等, 但相关实证研究较少。本文对隐性雌雄异株物种不同类群的性表达形式、不育性器官败育类型进行了归纳, 并对具该性系统的类群在被子植物中的分布与系统演化进行了分析与总结, 同时对有关隐性雌雄异株物种不育性器官进化意义的5个假说进行了介绍与评价, 最后对今后的相关研究方向进行了展望。以期为深入研究被子植物隐性雌雄异株性系统的进化式样与机制研究提供理论资料。 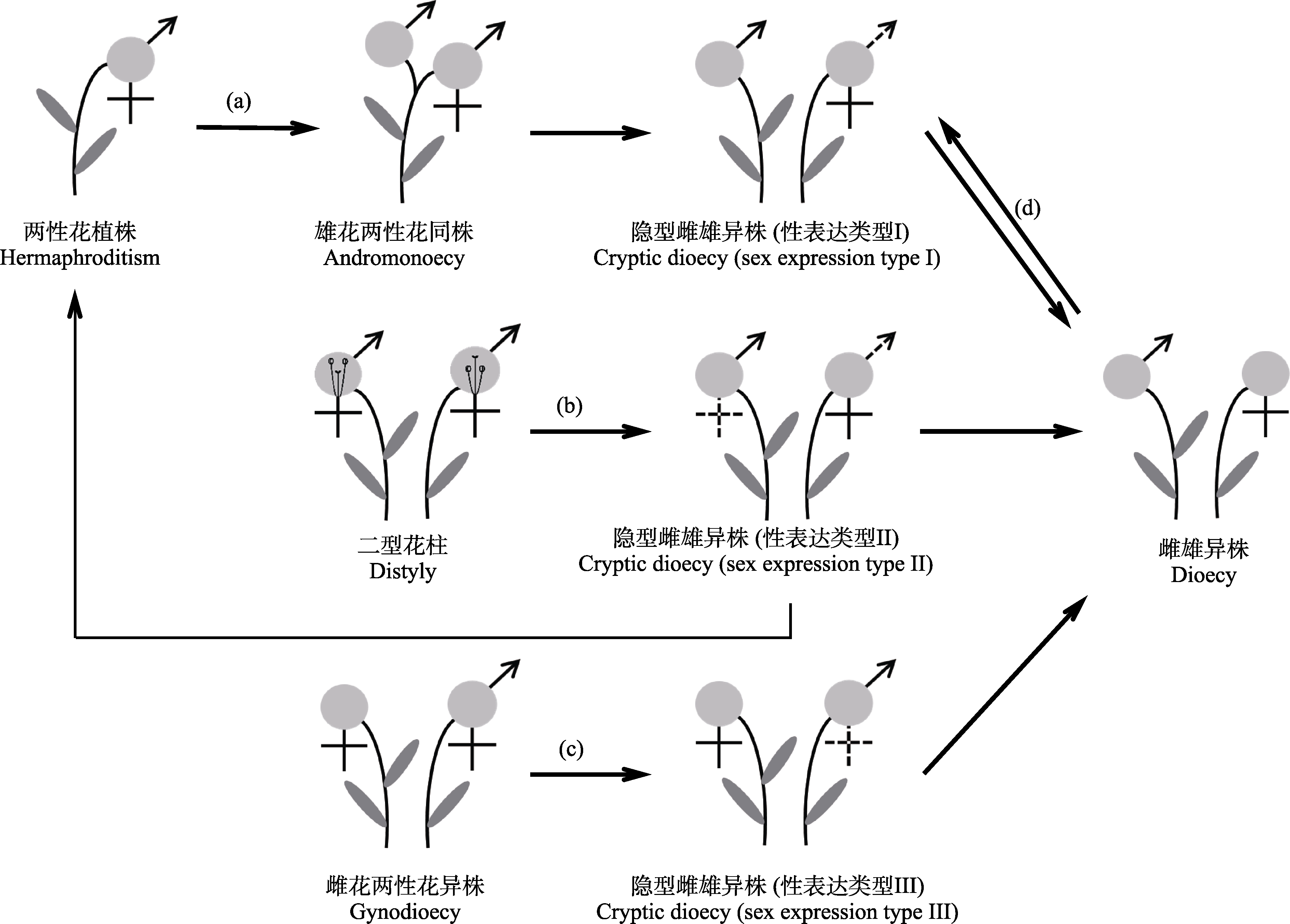 View image in article
图3
被子植物隐性雌雄异株可能的主要进化途径。(a)经雄花两性花同株进化为隐性雌雄异株性表达类型I; (b)由二型花柱直接进化为隐性雌雄异株性表达类型II; (c)由雌花两性花异株直接进化为隐性雌雄异株性表达类型III; (d)由雌雄异株直接进化为隐性雌雄异株性表达类型I。花及其性别用浅灰色圆圈表示。
正文中引用本图/表的段落
上述例证表明, 不同性表达类型的隐性雌雄异株物种其进化途径可能不同。表型性态为雄花两性花异株的隐性雌雄异株物种, 可能由两性花植株经雄花两性花同株进化而来(图3a); 表型性态为两性花的物种, 可直接由两性花(二型花柱)植株进化为隐性雌雄异株(图3b)。此外, 还有研究者认为雌花两性花异株和雌雄异株可分别进化为隐性雌雄异株性表达类型III (图3c) (Steiner, 1985)和类型I (图3d) (Penny, 2015), 但这些途径暂未证实, 关于隐性雌雄异株性系统的进化演变并未引起足够关注。
(1)已有对被子植物隐性雌雄异株演化关系的研究主要集中于茜草科的一些类群中, 而对已报道其他类群的系统位置及其演化关系的研究仍然较少, 那么隐性雌雄异株是如何进化而来? 处于进化的何种阶段? 这些还有待于进一步探究。Mayer和Charlesworth (1991)认为隐性雌雄异株正处于由两性花植株向雌雄异株进化的过渡阶段。已有研究为两性花(二型花柱)经隐性雌雄异株向雌雄异株进化提供了证据支持(邓小芳, 2007; 陈湜, 2013; 段婷婷, 2016), 但隐性雌雄异株物种表型性态多样(表1), 不同形态学性表达的隐性雌雄异株物种其进化途径可能不同, 而关于其他进化途径(图3)尚缺乏有力的实验数据支撑。此外, 对隐性雌雄异株性系统进化稳定性的研究还相对缺乏。开展隐性雌雄异株与两性花植株、雌雄异株、雄花两性花异株或雌花两性花异株的比较研究, 将有助于认识维持隐性雌雄异株的选择动力及深入理解其起源与进化。
本文的其它图/表
|

 表示两性花, 虚线表示该性别无功能, [ ]表示植株个体。
表示两性花, 虚线表示该性别无功能, [ ]表示植株个体。{kind=link}