
|
|
||
|
被子植物隐性雌雄异株性系统的多样性、系统演化及进化意义
生物多样性
2024, 32 (6):
24149-.
DOI: 10.17520/biods.2024149
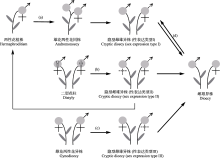
隐性雌雄异株指植株表型为非雌雄异株, 但形态上两性花通常雄性不育或雌性不育, 植株个体实际仅行使雌性或雄性功能。两性花植株向雌雄异株的转变在被子植物中广泛存在, 隐性雌雄异株作为潜在的中间阶段对于理解植物性系统的演化具有重要意义。据APG IV分类系统, 这些物种在被子植物的22目36科65属约221种植物中有过报道, 分别约占被子植物总目数的34.4%、总科数的8.7%、总属数的0.5%和总种数的0.1%; 多为分布于热带或亚热带地区依靠生物传粉的木本植物, 且至少存在花蜜或花粉作为访花报酬。该性系统形态学的性表达类型多样: 可分为雄花两性花异株(类型I)、两性花植株(类型II)、雌花两性花异株(类型III)及其他类型(类型IV) 4种, 分别约占已报道隐性雌雄异株物种数的48.9%、47.5%、2.7%和0.9%。从系统发育看, 该性系统在被子植物木兰分支、单子叶植物分支及真双子叶植物分支均有分布, 且主要分布于较为进化的核心真双子叶植物类群中。隐性雌雄异株物种不育性器官的进化意义存在多种假说, 包括祖先假说、遗传约束假说、促进风媒花粉落置假说、传粉者吸引假说、拟态与欺骗假说等, 但相关实证研究较少。本文对隐性雌雄异株物种不同类群的性表达形式、不育性器官败育类型进行了归纳, 并对具该性系统的类群在被子植物中的分布与系统演化进行了分析与总结, 同时对有关隐性雌雄异株物种不育性器官进化意义的5个假说进行了介绍与评价, 最后对今后的相关研究方向进行了展望。以期为深入研究被子植物隐性雌雄异株性系统的进化式样与机制研究提供理论资料。  View image in article
图2
具隐性雌雄异株性系统的分类群在被子植物中的系统分布。系统发育树参照APG IV (
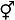 表示两性花, 虚线表示该性别无功能, [ ]表示植株个体。 表示两性花, 虚线表示该性别无功能, [ ]表示植株个体。
正文中引用本图/表的段落
根据APG IV分类系统(APG IV, 2016), 隐性雌雄异株性系统在被子植物的木兰类分支、单子叶分支和真双子叶植物分支中均有报道(图2)。木兰类分支和单子叶分支中, 分别仅出现于木兰科和棕榈科; 而真双子叶植物分支中, 在34科中有过隐性雌雄异株报道: 杜鹃花目5科, 金虎尾目、龙胆目和唇形目各3科, 无患子目、锦葵目、石竹目和伞形目各2科, 毛茛目、昆栏树目、五桠果目、葡萄目、豆目、蔷薇目、桃金娘目、山茱萸目、茶茱萸目、茄目、紫草目和菊目各有1科(表1, 图2)。从系统位置看, 具隐性雌雄异株性系统的科、属和种大多集中于被子植物较为进化的核心真双子叶植物分支, 且主要出现在分化相对较晚的桃金娘科、仙人掌科、茜草科、茄科等类群中。
具隐性雌雄异株性系统的36科中, 约69.4%的科内仅有1个属被报道, 但在以下科中有至少2个属存在隐性雌雄异株性系统: 茜草科(9属)、仙人掌科(7属)、茄科(6属)、桃金娘科(3属)、金虎尾科(3属)、无患子科(2属)、石竹科(2属)、猕猴桃科(2属)、杜鹃花科(2属)、菊科(2属)、马钱科(2属) (图2)。这表明隐性雌雄异株性系统在被子植物的不同科中多次独立起源。在属一级水平, 96.9%的属内隐性雌雄异株物种具有相同的性表达类型, 3.1%的属存在2种性表达, 如茜草科木巴戟属(Morinda) (Sugawara et al, 2011; Liu et al, 2012)和茄科茄属(Solanum) (Anderson et al, 2015; Ndem-Galbert et al, 2021)中同时存在形态上的雄花两性花异株和两性花植株2种性表达(图2)。
具隐性雌雄异株性系统的36科中, 约69.4%的科内仅有1个属被报道, 但在以下科中有至少2个属存在隐性雌雄异株性系统: 茜草科(9属)、仙人掌科(7属)、茄科(6属)、桃金娘科(3属)、金虎尾科(3属)、无患子科(2属)、石竹科(2属)、猕猴桃科(2属)、杜鹃花科(2属)、菊科(2属)、马钱科(2属) (
具隐性雌雄异株性系统的36科中, 约69.4%的科内仅有1个属被报道, 但在以下科中有至少2个属存在隐性雌雄异株性系统: 茜草科(9属)、仙人掌科(7属)、茄科(6属)、桃金娘科(3属)、金虎尾科(3属)、无患子科(2属)、石竹科(2属)、猕猴桃科(2属)、杜鹃花科(2属)、菊科(2属)、马钱科(2属) ( Pollination of androdioecious Xerospermum intermedium Radlk. (Sapindaceae) in a rain forest 2 1982 ... Sex expression types of cryptic dioecy and their distribution in angiosperms
具隐性雌雄异株性系统的36科中, 约69.4%的科内仅有1个属被报道, 但在以下科中有至少2个属存在隐性雌雄异株性系统: 茜草科(9属)、仙人掌科(7属)、茄科(6属)、桃金娘科(3属)、金虎尾科(3属)、无患子科(2属)、石竹科(2属)、猕猴桃科(2属)、杜鹃花科(2属)、菊科(2属)、马钱科(2属) (
具隐性雌雄异株性系统的36科中, 约69.4%的科内仅有1个属被报道, 但在以下科中有至少2个属存在隐性雌雄异株性系统: 茜草科(9属)、仙人掌科(7属)、茄科(6属)、桃金娘科(3属)、金虎尾科(3属)、无患子科(2属)、石竹科(2属)、猕猴桃科(2属)、杜鹃花科(2属)、菊科(2属)、马钱科(2属) ( Functional dioecy in Morinda parvifolia (Rubiaceae), a species with stigma-height dimorphism 3 2012 ... Sex expression types of cryptic dioecy and their distribution in angiosperms
本文的其它图/表
|

{kind=link}