被子植物隐性雌雄异株性系统的多样性、系统演化及进化意义
何花, 谭敦炎, 杨晓琛
生物多样性
2024, 32 ( 6):
24149-.
DOI: 10.17520/biods.2024149
隐性雌雄异株指植株表型为非雌雄异株, 但形态上两性花通常雄性不育或雌性不育, 植株个体实际仅行使雌性或雄性功能。两性花植株向雌雄异株的转变在被子植物中广泛存在, 隐性雌雄异株作为潜在的中间阶段对于理解植物性系统的演化具有重要意义。据APG IV分类系统, 这些物种在被子植物的22目36科65属约221种植物中有过报道, 分别约占被子植物总目数的34.4%、总科数的8.7%、总属数的0.5%和总种数的0.1%; 多为分布于热带或亚热带地区依靠生物传粉的木本植物, 且至少存在花蜜或花粉作为访花报酬。该性系统形态学的性表达类型多样: 可分为雄花两性花异株(类型I)、两性花植株(类型II)、雌花两性花异株(类型III)及其他类型(类型IV) 4种, 分别约占已报道隐性雌雄异株物种数的48.9%、47.5%、2.7%和0.9%。从系统发育看, 该性系统在被子植物木兰分支、单子叶植物分支及真双子叶植物分支均有分布, 且主要分布于较为进化的核心真双子叶植物类群中。隐性雌雄异株物种不育性器官的进化意义存在多种假说, 包括祖先假说、遗传约束假说、促进风媒花粉落置假说、传粉者吸引假说、拟态与欺骗假说等, 但相关实证研究较少。本文对隐性雌雄异株物种不同类群的性表达形式、不育性器官败育类型进行了归纳, 并对具该性系统的类群在被子植物中的分布与系统演化进行了分析与总结, 同时对有关隐性雌雄异株物种不育性器官进化意义的5个假说进行了介绍与评价, 最后对今后的相关研究方向进行了展望。以期为深入研究被子植物隐性雌雄异株性系统的进化式样与机制研究提供理论资料。
| 目 Order | 科 Family | 属 Genus | 种 Species | 参考文献 Reference | 性表达类型I Sex expression type I 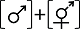 | | 木兰目 Magnoliales | 木兰科 Magnoliaceae | 拟单性木兰属 Parakmeria | 峨眉拟单性木兰 P. omeiensis | 余道平等, 2019 | | 棕榈目 Arecales | 棕榈科 Arecaceae | 海枣属 Phoenix | 海枣 P. dactylifera | 陆静等, 2021 | | 毛茛目 Ranunculales | 毛茛科 Ranunculaceae | 唐松草属 Thalictrum | T. macrostylum, T. polygamum, 柔毛唐松草 T. pubescens | Melampy & Hayworth, 1980; Davis SL, 2002; Penny, 2018 | | 昆栏树目 Trochodendrales | 昆栏树科 Trochodendraceae | 昆栏树属 Trochodendron | 昆栏树 T. aralioides | Keng, 1959 | | 五桠果目 Dilleniales | 五桠果科 Dilleniaceae | 锡叶藤属 Tetracera | 锡叶藤属14种 Tetracera sp. 1-14 | Kubitzki & Baretta-Kuipers, 1969 | | 葡萄目 Vitales | 葡萄科 Vitaceae | 葡萄属 Vitis | 夏葡萄 V. aestivalis, 河岸葡萄 V. riparia, V. vinifera ssp. silvestris | Brantjes, 1978; Kevan et al, 1985, 1988 | | 金虎尾目 Malpighiales | 藤黄科 Clusiaceae | 藤黄属 Garcinia | G. brasiliensis | Leal et al, 2012 | | 红厚壳科 Calophyllaceae | 南美杏属 Mammea | M. acuminata, M. africana, 马米杏 M. americana, M. sp. aff. bongo, M. eugenioides var. subsessilifolia, M. punctata, M. cf. punctata, M. siamensis | Dunthorn, 2004 | | 金虎尾科Malpighiaceae | 折背藤属 Madagasikaria | M. andersonii | Davis CC, 2002 | | 辐翅藤属 Rhynchophora | R. phillipsonii | Anderson WR, 2001 | | 豆目 Fabales | 豆科 Fabaceae | 皂荚属 Gleditsia | 南美皂荚 G. amorphoides | Cerino et al, 2018 | | 桃金娘目 Myrtales | 桃金娘科 Myrtaceae | 子楝树属 Decaspermum | 五瓣子楝树 D. parviflorum | Kevan & Lack, 1985 | | 番樱桃属 Eugenia | E. albanensis, E. capensis, E. cf. mossambicensis, E. erythrophylla, E. natalitia, E. simii, E. umtamvunensis, E. verdoorniae, E. woodii, E. zeyheri, E.zuluensis, 番樱桃属3种 Eugenia sp. 1-3 | van Wyk & Dedekind, 1985;
van der Merwe et al, 2005 | | 无患子目 Sapindales | 无患子科 Sapindaceae | 干果木属 Xerospermum | X. intermedium | Appanah, 1982 | | 石竹目 Caryophyllales | 石竹科 Caryophyllaceae | 冰漆姑属 Honckenya | H. peploides var. major | Tsukui & Sugawara, 1992 | | 山茱萸目 Cornales | 蓝果树科 Nyssaceae | 蓝果树属 Nyssa | 云南蓝果树 N. yunnanensis | Sun et al, 2009 | | 杜鹃花目 Ericales | 柿科 Ebenaceae | 柿属 Diospyros | 君迁子 D. lotus | 陆静等, 2021 | | 山矾科 Symplocaceae | 山矾属 Symplocos | S. celastrinea, S. crenata, S. itatiaiae, S. lanceolata, S. mosenii, S. rhamnifolia, S. saxatilis, S. tetrandra, S. variabilis, S. inopinata, S. kleinii, S. occulta, 微毛山矾 S. wikstroemiifolia | Aranha Filho et al, 2009, 2013; Wang & Hu, 2011 | | 猕猴桃科 Actinidiaceae | 猕猴桃属 Actinidia | 软枣猕猴桃 A. arguta, 中华猕猴桃 A. chinensis, 美味猕猴桃 A. deliciosa var. deliciosa, 毛花猕猴桃 A. eriantha, 绵毛猕猴桃 A. fulvicoma var. lanata, 革叶猕猴桃 A. rubricaulis var. coriacea, 对萼猕猴桃 A. valvata, 葛枣猕猴桃 A. polygama | unpublished | | 水东哥属 Saurauia | S. bullosa, S. brachybotrys, S. excelsa, S. humboldtiana, S. tomentosa, S. putumayonis, S. ursina, S. veraguensis | Soejarto, 1969; Cane, 1993 | | 伞形目 Apiales | 毛柴木科 Pennantiaceae | 毛柴木属 Pennantia | 毛柴木属3种 Pennantia sp. 1-3 | Baylis, 1977 | | 五加科 Araliaceae | 南洋参属 Polyscias | P. dzumacensis, P. microbotrys, P. pancheri, P. subincisa | Schlessman et al, 1990 | | 紫草目 Boraginales | 紫草科 Boraginaceae | 破布木属 Cordia | C. crenata, C. cf. grandicalyx, C. sinensis | Pätzold et al, 2023 | | 龙胆目 Gentianales | 茜草科 Rubiaceae | 木巴戟属 Morinda | 鸡眼藤 M. parvifolia | Liu et al, 2012 | | 唇形目 Lamiales | 木樨科 Oleaceae | 梣属 Fraxinus | 维斯谷归化种群中的花梣 Fraxinus ornus in naturalized Vis Valley populations | Vernet et al, 2016 | | 茄目 Solanales | 茄科 Solanaceae | 茄属 Solanum | S. asymmetriphyllum, S. carduiforme, S. cataphractum, S. cowiei, S. cunninghamii, S. dioicum, S. leopoldense, S. ossicruentum, S. petraeum, S. polygamum, S. tudununggae, S. vansittartensis, S. sejunctum | Anderson & Symon, 1989; Martine et al, 2013; Anderson et al, 2015; Ndem-Galbert et al, 2021 | 性表达类型II Sex expression type II 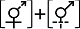 | | 蔷薇目 Rosales | 蔷薇科 Rosaceae | 蔷薇属 Rosa | 草原玫瑰 R. setigera | Kevan et al, 1990 | | 桃金娘目 Myrtales | 桃金娘科 Myrtaceae | 多香果属 Pimenta | 多香果 P. dioica | Boyd & Benkeblia, 2011 | | 无患子目 Sapindales | 无患子科 Sapindaceae | 槭属 Acer | 红花槭 A. rubrum | Batra, 1985 | | 楝科 Meliaceae | 杜楝属 Turraea | T. dargentiana | Baider & Florens, 2016 | | 锦葵目 Malvales | 瑞香科 Thymelaeaceae | 荛花属 Wikstroemia | 夏威夷岛和可爱岛中荛花属种群
Wikstroemia populations on Kauai, Hawaii | Mayer & Charlesworth, 1991 | | 锦葵科 Malvaceae | 非洲芙蓉属 Dombeya | D. ciliata, D. delislei, D. elegans, D. ficulnea, D. formosa, D. pilosa, D. punctata, D. reclinata | Humeau et al, 1999, 2000;
Le Péchon et al, 2013 | | 石竹目 Caryophyllales | 仙人掌科 Cactaceae | 旗号掌属 Consolea | C. corallicola, C. millspaughii, 鱼鳞掌 C. moniliformis, C. nashii, C. picardae, C. rubescens, C. spinosissima | Rebman & Pinkava, 2001; Strittmatter et al, 2006, 2008 | | 鹿角柱属 Echinocereus | 赤花虾 E. coccineus, E. mombergerianus, E. pacificus, 多刺虾鹿角柱 E. polyacanthus | Hernández-Cruz et al, 2018 | | 仙人掌属 Opuntia | O. quitensis, O. robusta, O. stenopetala | Anderson EF, 2001; Hernández-Cruz et al, 2019 | | 摩天柱属 Pachycereus | 武伦柱 P. pringlei | Fleming, 1989 | | 木麒麟属 Pereskia | P. portulacifolia, P. marcanoi, P. quisqueyana, P. zinniiflora | Areces-Mallea, 1992 | | 杜鹃花目 Ericales | 杜鹃花科 Ericaceae | 南白珠属 Pernettya | P. rigida | Anderson et al, 2000 | | 菊目 Asterales | 菊科 Asteraceae | 假泽兰属 Mikania | 假泽兰属5种 Mikania 5 sp. 1-5 | Holmes, 1991 | | 茶茱萸目 Icacinales | 茶茱萸科 Icacinaceae | 假柴龙树属 Nothapodytes | 臭味假柴龙树 N. nimmoniana | Sharma et al, 2010 | | 龙胆目 Gentianales | 茜草科 Rubiaceae
| 舒榄属 Bertiera | B. borbonica | Pailler et al, 1998b | | 弯管花属 Chassalia | C. corallioides | Pailler et al, 1998a | | 牛眼棠属 Cordiera | C. concolor | Judkevich et al, 2022 | | | 靛榄属 Genipa | 靛榄 G. americana | Judkevich et al, 2022 | | 蓝茜树属 Randia | R. calycina, R. heteromera | Judkevich et al, 2022 | | 木巴戟属 Morinda | 须弥巴戟 M. villosa | Sugawara et al, 2011 | | 玉叶金花属 Mussaenda | 短裂玉叶金花 M. breviloba, 尾裂玉叶金花 M. caudatiloba, 仁昌玉叶金花 M. chingii, 展枝玉叶金花 M. divaricata, 楠藤 M. erosa, 海南玉叶金花 M. hainanensis, 粗毛玉叶金花 M. hirsutula, 红毛玉叶金花 M. hossei, 广西玉叶金花 M. kwangsiensis, 广东玉叶金花 M. kwangtungensis, 疏花玉叶金花 M. laxiflora, 长瓣玉叶金花 M. longipetala, 多毛玉叶金花 M. mollissima, 小玉叶金花 M. parviflora, M. pilosissima, 屏边玉叶金花 M. pingbianensis, 玉叶金花 M. pubescens, 白花玉叶金花 M. pubescens var. alba, 贡山玉叶金花 M. treutleri, 玉叶金花属5种 Mussaenda sp. 1-5 | 邓小芳, 2007; Li et al, 2010; 陈湜, 2013 | | 沙九节属 Pagamea | P. duckei | Terra-Araujo et al, 2012 | | 九节属 Psychotria | 九节 P. rubra | Watanabe et al, 2013 | | 马钱科 Loganiaceae | 髯管花属 Geniostoma | G. borbonica | Humeau et al, 2003 | | 顶髯管花属 Labordia | L. cyrtandrae, L. degeneri, L. fagraeoidea, L. hedyosmifolia, L. hirtella, L. hosakana, L. lydgatei, L. pumila, L. sessilis, L.venosa, L. waialealae, L. waiolani | Motley & Carr, 1998 | | 夹竹桃科 Apocynaceae | 萝芙木属 Rauvolfia | R. sellowii | Souto & Koch, 2018 | | 唇形目 Lamiales | 马鞭草科 Verbenaceae | 琴木属 Citharexylum | C. myrianthum | Rocca & Sazima, 2006 | | 唇形科 Lamiaceae | 紫珠属 Callicarpa | C. glabra, C. nishimurae, C. subpubescens | Kawakubo, 1998 | | 茄目 Solanales | 茄科 Solanaceae | 山盏茄属 Deprea | D. paneroi | Sawyer & Anderson, 2000 | | 巫灵木属 Dunalia | D. solanacea | Hepper, 1991 | | 枸杞属 Lycium | L. arenicolum, L. horridum, L. tetrandrum, L. villosum | Spies et al, 1993 | | 茄目 Solanales | 茄科 Solanaceae | 茄属 Solanum | S. appendiculatum, S. campanulatum, S. confertiseriatum, S. crotonoides, S. conocarpum | Knapp et al, 1998;
Anderson et al, 2015 | | 串钟花属 Symonanthus | S. aromaticus, S. bancroftii | Haegi, 1981 | | 睡茄属 Withania | W. aristata, W. coagulans | Sawyer & Anderson, 2000; Anderson et al, 2006 | 性表达类型III Sex expression type III 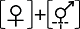 | | 金虎尾目 Malpighiales | 金虎尾科Malpighiaceae | 异金英属 Spachea | S. membranacea | Steiner, 1985 | | 石竹目 Caryophyllales | 石竹科Caryophyllaceae | 洋漆姑属 Schiedea | S. adamantis | Mayer & Charlesworth, 1991; Sakai et al, 1997 | | 仙人掌科 Cactaceae | 圆柱掌属 Cylindropuntia | C. wolfii | Ramadoss et al, 2022 | | 裸萼球属 Gymnocalycium | 罗星丸 G. bruchii | Delbón et al, 2024 | | 杜鹃花目 Ericales | 山榄科 Sapotaceae | 星苹果属 Chrysophyllum | C. gonocarpum | Judkevich et al, 2023 | | 杜鹃花科 Ericaceae | 岩梨属 Epigaea | 岩梨 E. repens | Clay & Ellstrand, 1981 | 性表达类型IV Sex expression type IV (1) 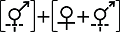 (2) (2) 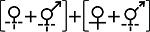 | | 菊目 Asterales | 菊科 Asteraceae | 蜂斗菜属 Petasites | (1)蜂斗菜 P. japonicas; (2)毛裂蜂斗菜 P. tricholobus | Sakai et al, 2008; Yu et al, 2011 |
View table in article
表1
隐性雌雄异株的性表达类型及其在被子植物中的分布
正文中引用本图/表的段落
据Mayer和Charlesworth (1991)的报道, 被子植物中隐性雌雄异株性系统主要出现在五桠果科、桃金娘科、茄科、菊科等18科21属78种植物中(原茶茱萸科3种毛柴木属(Pennantia)植物现属毛柴木科): 其中, 五桠果科22种, 约占28.2% (其中8种水东哥属(Saurauia)植物现属猕猴桃科); 桃金娘科16种, 约占20.5%; 茄科10种, 约占12.8%; 菊科5种, 约占6.4%。此后, 又在唇形科(Kawakubo, 1998)、金虎尾科(Davis CC, 2002)、马钱科(Humeau et al, 2003)、红厚壳科(Dunthorn, 2004)、蓝果树科(Sun et al, 2009)、藤黄科(Leal et al, 2012)、山矾科(Aranha Filho et al, 2013)、锦葵科(Le Péchon et al, 2013)、柿科(Akagi et al, 2014)、木樨科(Vernet et al, 2016)、豆科(Cerino et al, 2018)、夹竹桃科(Souto & Koch, 2018)、棕榈科(Torres et al, 2018)、木兰科(余道平等, 2019)、茜草科(Judkevich et al, 2022)、山榄科(Judkevich et al, 2023)和紫草科(P?tzold et al, 2023)等类群中发现了隐性雌雄异株性系统。至此, 隐性雌雄异株性系统已在被子植物的22目36科65属约221个物种有过报道(表1), 分别约占被子植物总目数(64)的34.4%和总科数(416)的8.7% (APG Ⅳ, 2016)、总属数(14,158)的0.5%和总种数(344,397)的0.1% (World Flora Online, https://www.worldfloraonline.org/, 数据截止日期为2024年6月)。
隐性雌雄异株物种形态学性表达和功能性性表达不一致, 仅从花表型很难判断实际繁殖功能。根据已有研究报道, 隐性雌雄异株物种形态学性表达可分为雄花两性花异株(类型I)、两性花植株(类型II)、雌花两性花异株(类型III)及其他类型(类型IV) 4种, 分别约占已报道隐性雌雄异株物种数的48.9%、47.5%、2.7%和0.9% (表1)。
(1)雄花两性花异株(类型I)。这是隐性雌雄异株最常见的形态学性表达类型(Mayer & Charlesworth, 1991), 即雄花植株只产生表型与功能一致的雄花, 而两性花植株其形态上的两性花雄蕊不育, 实际为功能性的雌花。如昆栏树科昆栏树(Trochodendron aralioides) (Keng, 1959)、桃金娘科五瓣子楝树(Decaspermum parviflorum) (Kevan & Lack, 1985)、五加科Polyscias dzumacensis (Schlessman et al, 1990)、猕猴桃科葛枣猕猴桃(Actinidia polygama) (Kawagoe & Suzuki, 2004)等物种(表1, 图1A)。该类型被报道约108个物种, 约96.3% (104/108)为木本植物; 主要生长于热带或亚热带生物群落中(81.2%, 69/85, 23种无相关信息, 下同); 大都依赖生物媒介进行传粉(89.9%, 71/79), 其余10.1% (8/79)的物种为生物和非生物传粉共存; 该类型物种多不产花蜜(74%, 37/50), 主要以花粉作为传粉者访花报酬(雌性植株不育雄蕊提供花粉的物种约占87.6%, 92/105) (附录1)。
(2)两性花植株(类型II)。即形态均为两性花的植株, 部分植株的所有两性花雄性不育而成为功能性的雌花、部分植株的所有两性花则是雌性不育而成为功能性的雄花, 从而导致了隐性雌雄异株。如无患子科红花槭(Acer rubrum) (Batra, 1985)、仙人掌科武伦柱(Pachycereus pringlei) (Fleming, 1989)、杜鹃花科Pernettya rigida (Anderson et al, 2000)、茜草科玉叶金花(Mussaenda pubescens) (Li et al, 2010)、锦葵科Dombeya formosa (Le Péchon et al, 2013)等物种(表1, 图1B)。该类型被报道约105个物种, 均为木本植物(97/97, 8个物种无相关信息, 下同); 主要生长于热带或亚热带生物群落中(94.6%, 87/92), 大都依赖生物媒介进行传粉(98.7%, 75/76), 仅1种为非生物传粉。雌性植株不育雄蕊多不提供花粉(72.6%, 61/84), 主要以花蜜作为传粉者访花报酬(89.5%, 51/57) (附录1)。
(3)雌花两性花异株(类型III)。即雌花植株只产生表型与功能一致的雌花, 而形态学两性花植株实际雌蕊不育, 成为功能性雄花植株。如杜鹃花科岩梨(Epigaea repens) (Clay & Ellstrand, 1981)、石竹科Schiedea adamantis (Sakai et al, 1997)、山榄科Chrysophyllum gonocarpum (Judkevich et al, 2023), 其形态学两性花雌蕊不育(表1)。该类型被报道约6个物种, 均为木本植物; 主要生长于热带或亚热带生物群落中(83.3%, 5/6); 3种为生物传粉, 剩余3种未报道传粉媒介; 已报道访花报酬的3种均存在花蜜(附录1)。
(4)其他类型(类型IV)。该类型性表达形式相对复杂, 目前仅报道了菊科蜂斗菜属(Petasites) 2个物种(表1)。如毛裂蜂斗菜(P. tricholobus)所有植株均产生形态上的两性花和雌花, 但功能性态为雌性的植株中所有两性花不育, 仅雌花行使雌性功能, 而功能性态为雄性的植株中所有两性花及雌花的雌蕊均不育, 仅两性花的雄蕊正常行使雄性功能(Yu et al, 2011)。蜂斗菜(P. japonicus)和毛裂蜂斗菜的雌性植株性表达一致, 但雄性植株上只产生雌蕊不育的形态上的两性花, 并无雌花(Sakai et al, 2008) (表1)。被报道的2个物种均为生长于温带生物群落中的生物传粉的多年生草本植物; 其不育雄蕊均不产生花粉, 但毛裂蜂斗菜的雌性植株中存在花蜜, 蜂斗菜花蜜信息未见有报道(附录1)。
根据APG IV分类系统(APG IV, 2016), 隐性雌雄异株性系统在被子植物的木兰类分支、单子叶分支和真双子叶植物分支中均有报道(图2)。木兰类分支和单子叶分支中, 分别仅出现于木兰科和棕榈科; 而真双子叶植物分支中, 在34科中有过隐性雌雄异株报道: 杜鹃花目5科, 金虎尾目、龙胆目和唇形目各3科, 无患子目、锦葵目、石竹目和伞形目各2科, 毛茛目、昆栏树目、五桠果目、葡萄目、豆目、蔷薇目、桃金娘目、山茱萸目、茶茱萸目、茄目、紫草目和菊目各有1科(表1, 图2)。从系统位置看, 具隐性雌雄异株性系统的科、属和种大多集中于被子植物较为进化的核心真双子叶植物分支, 且主要出现在分化相对较晚的桃金娘科、仙人掌科、茜草科、茄科等类群中。
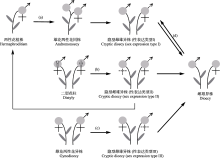
(1)已有对被子植物隐性雌雄异株演化关系的研究主要集中于茜草科的一些类群中, 而对已报道其他类群的系统位置及其演化关系的研究仍然较少, 那么隐性雌雄异株是如何进化而来? 处于进化的何种阶段? 这些还有待于进一步探究。Mayer和Charlesworth (1991)认为隐性雌雄异株正处于由两性花植株向雌雄异株进化的过渡阶段。已有研究为两性花(二型花柱)经隐性雌雄异株向雌雄异株进化提供了证据支持(邓小芳, 2007; 陈湜, 2013; 段婷婷, 2016), 但隐性雌雄异株物种表型性态多样(表1), 不同形态学性表达的隐性雌雄异株物种其进化途径可能不同, 而关于其他进化途径(图3)尚缺乏有力的实验数据支撑。此外, 对隐性雌雄异株性系统进化稳定性的研究还相对缺乏。开展隐性雌雄异株与两性花植株、雌雄异株、雄花两性花异株或雌花两性花异株的比较研究, 将有助于认识维持隐性雌雄异株的选择动力及深入理解其起源与进化。
♂表示雄花, ♀表示雌花, 表示两性花; 虚线表示该性别无功能; [ ]表示植株个体. ... An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV 4 2016 ... 据Mayer和Charlesworth (1991)的报道, 被子植物中隐性雌雄异株性系统主要出现在五桠果科、桃金娘科、茄科、菊科等18科21属78种植物中(原茶茱萸科3种毛柴木属(Pennantia)植物现属毛柴木科): 其中, 五桠果科22种, 约占28.2% (其中8种水东哥属(Saurauia)植物现属猕猴桃科); 桃金娘科16种, 约占20.5%; 茄科10种, 约占12.8%; 菊科5种, 约占6.4%.此后, 又在唇形科(Kawakubo, 1998)、金虎尾科(Davis CC, 2002)、马钱科(Humeau et al, 2003)、红厚壳科(Dunthorn, 2004)、蓝果树科(Sun et al, 2009)、藤黄科(Leal et al, 2012)、山矾科(Aranha Filho et al, 2013)、锦葵科(Le Péchon et al, 2013)、柿科(Akagi et al, 2014)、木樨科(Vernet et al, 2016)、豆科(Cerino et al, 2018)、夹竹桃科(Souto & Koch, 2018)、棕榈科(Torres et al, 2018)、木兰科(余道平等, 2019)、茜草科(Judkevich et al, 2022)、山榄科(Judkevich et al, 2023)和紫草科(P?tzold et al, 2023)等类群中发现了隐性雌雄异株性系统.至此, 隐性雌雄异株性系统已在被子植物的22目36科65属约221个物种有过报道(表1), 分别约占被子植物总目数(64)的34.4%和总科数(416)的8.7% (APG Ⅳ, 2016)、总属数(14,158)的0.5%和总种数(344,397)的0.1% (World Flora Online, https://www.worldfloraonline.org/, 数据截止日期为2024年6月). ...
♂表示雄花, ♀表示雌花, 表示两性花; 虚线表示该性别无功能; [ ]表示植株个体. ... Three new species of Symplocos sect. Hopea (Symplocaceae) from Brazil 2 2013 ... 据Mayer和Charlesworth (1991)的报道, 被子植物中隐性雌雄异株性系统主要出现在五桠果科、桃金娘科、茄科、菊科等18科21属78种植物中(原茶茱萸科3种毛柴木属(Pennantia)植物现属毛柴木科): 其中, 五桠果科22种, 约占28.2% (其中8种水东哥属(Saurauia)植物现属猕猴桃科); 桃金娘科16种, 约占20.5%; 茄科10种, 约占12.8%; 菊科5种, 约占6.4%.此后, 又在唇形科(Kawakubo, 1998)、金虎尾科(Davis CC, 2002)、马钱科(Humeau et al, 2003)、红厚壳科(Dunthorn, 2004)、蓝果树科(Sun et al, 2009)、藤黄科(Leal et al, 2012)、山矾科(Aranha Filho et al, 2013)、锦葵科(Le Péchon et al, 2013)、柿科(Akagi et al, 2014)、木樨科(Vernet et al, 2016)、豆科(Cerino et al, 2018)、夹竹桃科(Souto & Koch, 2018)、棕榈科(Torres et al, 2018)、木兰科(余道平等, 2019)、茜草科(Judkevich et al, 2022)、山榄科(Judkevich et al, 2023)和紫草科(P?tzold et al, 2023)等类群中发现了隐性雌雄异株性系统.至此, 隐性雌雄异株性系统已在被子植物的22目36科65属约221个物种有过报道(表1), 分别约占被子植物总目数(64)的34.4%和总科数(416)的8.7% (APG Ⅳ, 2016)、总属数(14,158)的0.5%和总种数(344,397)的0.1% (World Flora Online, https://www.worldfloraonline.org/, 数据截止日期为2024年6月). ...
♂表示雄花, ♀表示雌花, 表示两性花; 虚线表示该性别无功能; [ ]表示植株个体. ... Functional dioecy in Gleditsia amorphoides (Fabaceae) 3 2018 ... 据Mayer和Charlesworth (1991)的报道, 被子植物中隐性雌雄异株性系统主要出现在五桠果科、桃金娘科、茄科、菊科等18科21属78种植物中(原茶茱萸科3种毛柴木属(Pennantia)植物现属毛柴木科): 其中, 五桠果科22种, 约占28.2% (其中8种水东哥属(Saurauia)植物现属猕猴桃科); 桃金娘科16种, 约占20.5%; 茄科10种, 约占12.8%; 菊科5种, 约占6.4%.此后, 又在唇形科(Kawakubo, 1998)、金虎尾科(Davis CC, 2002)、马钱科(Humeau et al, 2003)、红厚壳科(Dunthorn, 2004)、蓝果树科(Sun et al, 2009)、藤黄科(Leal et al, 2012)、山矾科(Aranha Filho et al, 2013)、锦葵科(Le Péchon et al, 2013)、柿科(Akagi et al, 2014)、木樨科(Vernet et al, 2016)、豆科(Cerino et al, 2018)、夹竹桃科(Souto & Koch, 2018)、棕榈科(Torres et al, 2018)、木兰科(余道平等, 2019)、茜草科(Judkevich et al, 2022)、山榄科(Judkevich et al, 2023)和紫草科(P?tzold et al, 2023)等类群中发现了隐性雌雄异株性系统.至此, 隐性雌雄异株性系统已在被子植物的22目36科65属约221个物种有过报道(表1), 分别约占被子植物总目数(64)的34.4%和总科数(416)的8.7% (APG Ⅳ, 2016)、总属数(14,158)的0.5%和总种数(344,397)的0.1% (World Flora Online, https://www.worldfloraonline.org/, 数据截止日期为2024年6月). ...
♂表示雄花, ♀表示雌花, 表示两性花; 虚线表示该性别无功能; [ ]表示植株个体. ... Dombeya formosa (Malvaceae s.l.): A new species endemic to La Réunion (Indian Ocean) based on morphological and molecular evidence 2 2013 ... 据Mayer和Charlesworth (1991)的报道, 被子植物中隐性雌雄异株性系统主要出现在五桠果科、桃金娘科、茄科、菊科等18科21属78种植物中(原茶茱萸科3种毛柴木属(Pennantia)植物现属毛柴木科): 其中, 五桠果科22种, 约占28.2% (其中8种水东哥属(Saurauia)植物现属猕猴桃科); 桃金娘科16种, 约占20.5%; 茄科10种, 约占12.8%; 菊科5种, 约占6.4%.此后, 又在唇形科(Kawakubo, 1998)、金虎尾科(Davis CC, 2002)、马钱科(Humeau et al, 2003)、红厚壳科(Dunthorn, 2004)、蓝果树科(Sun et al, 2009)、藤黄科(Leal et al, 2012)、山矾科(Aranha Filho et al, 2013)、锦葵科(Le Péchon et al, 2013)、柿科(Akagi et al, 2014)、木樨科(Vernet et al, 2016)、豆科(Cerino et al, 2018)、夹竹桃科(Souto & Koch, 2018)、棕榈科(Torres et al, 2018)、木兰科(余道平等, 2019)、茜草科(Judkevich et al, 2022)、山榄科(Judkevich et al, 2023)和紫草科(P?tzold et al, 2023)等类群中发现了隐性雌雄异株性系统.至此, 隐性雌雄异株性系统已在被子植物的22目36科65属约221个物种有过报道(表1), 分别约占被子植物总目数(64)的34.4%和总科数(416)的8.7% (APG Ⅳ, 2016)、总属数(14,158)的0.5%和总种数(344,397)的0.1% (World Flora Online, https://www.worldfloraonline.org/, 数据截止日期为2024年6月). ...
♂表示雄花, ♀表示雌花, 表示两性花; 虚线表示该性别无功能; [ ]表示植株个体. ... Floral structure of Garcinia brasiliensis in relation to flower biology and evolution 3 2012 ... 据Mayer和Charlesworth (1991)的报道, 被子植物中隐性雌雄异株性系统主要出现在五桠果科、桃金娘科、茄科、菊科等18科21属78种植物中(原茶茱萸科3种毛柴木属(Pennantia)植物现属毛柴木科): 其中, 五桠果科22种, 约占28.2% (其中8种水东哥属(Saurauia)植物现属猕猴桃科); 桃金娘科16种, 约占20.5%; 茄科10种, 约占12.8%; 菊科5种, 约占6.4%.此后, 又在唇形科(Kawakubo, 1998)、金虎尾科(Davis CC, 2002)、马钱科(Humeau et al, 2003)、红厚壳科(Dunthorn, 2004)、蓝果树科(Sun et al, 2009)、藤黄科(Leal et al, 2012)、山矾科(Aranha Filho et al, 2013)、锦葵科(Le Péchon et al, 2013)、柿科(Akagi et al, 2014)、木樨科(Vernet et al, 2016)、豆科(Cerino et al, 2018)、夹竹桃科(Souto & Koch, 2018)、棕榈科(Torres et al, 2018)、木兰科(余道平等, 2019)、茜草科(Judkevich et al, 2022)、山榄科(Judkevich et al, 2023)和紫草科(P?tzold et al, 2023)等类群中发现了隐性雌雄异株性系统.至此, 隐性雌雄异株性系统已在被子植物的22目36科65属约221个物种有过报道(表1), 分别约占被子植物总目数(64)的34.4%和总科数(416)的8.7% (APG Ⅳ, 2016)、总属数(14,158)的0.5%和总种数(344,397)的0.1% (World Flora Online, https://www.worldfloraonline.org/, 数据截止日期为2024年6月). ...
♂表示雄花, ♀表示雌花, 表示两性花; 虚线表示该性别无功能; [ ]表示植株个体. ... Observations on flower and fruit anatomy in dioecious species of Cordia (Cordiaceae, Boraginales) with evolutionary interpretations 4 2023 ... 据Mayer和Charlesworth (1991)的报道, 被子植物中隐性雌雄异株性系统主要出现在五桠果科、桃金娘科、茄科、菊科等18科21属78种植物中(原茶茱萸科3种毛柴木属(Pennantia)植物现属毛柴木科): 其中, 五桠果科22种, 约占28.2% (其中8种水东哥属(Saurauia)植物现属猕猴桃科); 桃金娘科16种, 约占20.5%; 茄科10种, 约占12.8%; 菊科5种, 约占6.4%.此后, 又在唇形科(Kawakubo, 1998)、金虎尾科(Davis CC, 2002)、马钱科(Humeau et al, 2003)、红厚壳科(Dunthorn, 2004)、蓝果树科(Sun et al, 2009)、藤黄科(Leal et al, 2012)、山矾科(Aranha Filho et al, 2013)、锦葵科(Le Péchon et al, 2013)、柿科(Akagi et al, 2014)、木樨科(Vernet et al, 2016)、豆科(Cerino et al, 2018)、夹竹桃科(Souto & Koch, 2018)、棕榈科(Torres et al, 2018)、木兰科(余道平等, 2019)、茜草科(Judkevich et al, 2022)、山榄科(Judkevich et al, 2023)和紫草科(P?tzold et al, 2023)等类群中发现了隐性雌雄异株性系统.至此, 隐性雌雄异株性系统已在被子植物的22目36科65属约221个物种有过报道(表1), 分别约占被子植物总目数(64)的34.4%和总科数(416)的8.7% (APG Ⅳ, 2016)、总属数(14,158)的0.5%和总种数(344,397)的0.1% (World Flora Online, https://www.worldfloraonline.org/, 数据截止日期为2024年6月). ...
♂表示雄花, ♀表示雌花, 表示两性花; 虚线表示该性别无功能; [ ]表示植株个体. ... Explaining ovule sterility in Rauvolfia L. (Apocynaceae) 5 2018 ... 据Mayer和Charlesworth (1991)的报道, 被子植物中隐性雌雄异株性系统主要出现在五桠果科、桃金娘科、茄科、菊科等18科21属78种植物中(原茶茱萸科3种毛柴木属(Pennantia)植物现属毛柴木科): 其中, 五桠果科22种, 约占28.2% (其中8种水东哥属(Saurauia)植物现属猕猴桃科); 桃金娘科16种, 约占20.5%; 茄科10种, 约占12.8%; 菊科5种, 约占6.4%.此后, 又在唇形科(Kawakubo, 1998)、金虎尾科(Davis CC, 2002)、马钱科(Humeau et al, 2003)、红厚壳科(Dunthorn, 2004)、蓝果树科(Sun et al, 2009)、藤黄科(Leal et al, 2012)、山矾科(Aranha Filho et al, 2013)、锦葵科(Le Péchon et al, 2013)、柿科(Akagi et al, 2014)、木樨科(Vernet et al, 2016)、豆科(Cerino et al, 2018)、夹竹桃科(Souto & Koch, 2018)、棕榈科(Torres et al, 2018)、木兰科(余道平等, 2019)、茜草科(Judkevich et al, 2022)、山榄科(Judkevich et al, 2023)和紫草科(P?tzold et al, 2023)等类群中发现了隐性雌雄异株性系统.至此, 隐性雌雄异株性系统已在被子植物的22目36科65属约221个物种有过报道(表1), 分别约占被子植物总目数(64)的34.4%和总科数(416)的8.7% (APG Ⅳ, 2016)、总属数(14,158)的0.5%和总种数(344,397)的0.1% (World Flora Online, https://www.worldfloraonline.org/, 数据截止日期为2024年6月). ...
本文的其它图/表
|


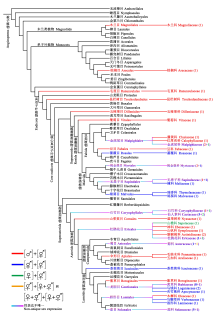
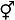 表示两性花, 虚线表示该性别无功能, [ ]表示植株个体。
表示两性花, 虚线表示该性别无功能, [ ]表示植株个体。