
|
|
||
|
鸟类迁徙对图们江下游湿地声景时间格局的影响
生物多样性
2023, 31 (1):
22337-.
DOI: 10.17520/biods.2022337
声景生态学是一个相对较新和快速发展的研究领域, 被动声学监测技术和声学指数已经成为研究湿地鸟类和声景多样性的重要方法。本研究评价了鸟类迁徙对中国东北图们江流域下游湿地声景日、月和季节变化的影响。我们从2020年11月至2021年12月在图们江下游敬信湿地设置10个采样点, 获得91,988条时长5 min的有效音频, 计算了声音复杂度指数(acoustic complexity index, ACI)、生物声学指数(bioacoustic index, BIO)、声音均匀度指数(acoustic evenness index, AEI)和标准化声景差异指数(normalized difference soundscape index, NDSI)以及1-11 kHz频段的功率谱密度(power spectral density, PSD)。结果表明, 声学指数对鸟类迁徙活动敏感, 其中2个迁徙期声景(2-4月和10-11月)都以1-2 kHz雁类白天的叫声为主, NDSI显著降低, 1-2 kHz的PSD显著升高, 但雁类向北迁徙时几个声学指数变化更为敏感, 有效地捕获了迁徙峰值, 表明不同季节鸟类迁徙模式存在差异。非迁徙期声景由夏候鸟、蛙类和昆虫发声为主, 4种声学指数和PSD随月份呈现不同的动态特征, 反映了声景的多样性和复杂性, 其中5-7月声景以2-11 kHz的夏候鸟鸣唱(呈现显著高的黎明和鸣行为)和2-3 kHz的蛙类鸣叫为主, 8-9月声景以2-3 kHz、4-5 kHz和6-10 kHz频段的夜间昆虫鸣叫为主, 12月至次年1月仅记录到少量的鸟类发声活动。综上所述, 图们江下游湿地声景呈现明显的日和月变化规律, 多种声学指数联合使用可以有效地监测迁徙鸟类物候的变化, 特别是追踪春季雁类向北迁徙的时间和规模。随着全球气候变暖, 我们的结果强调声景监测与声学指数的应用可成为监测迁徙鸟类群落对气候变化响应的有效方法。 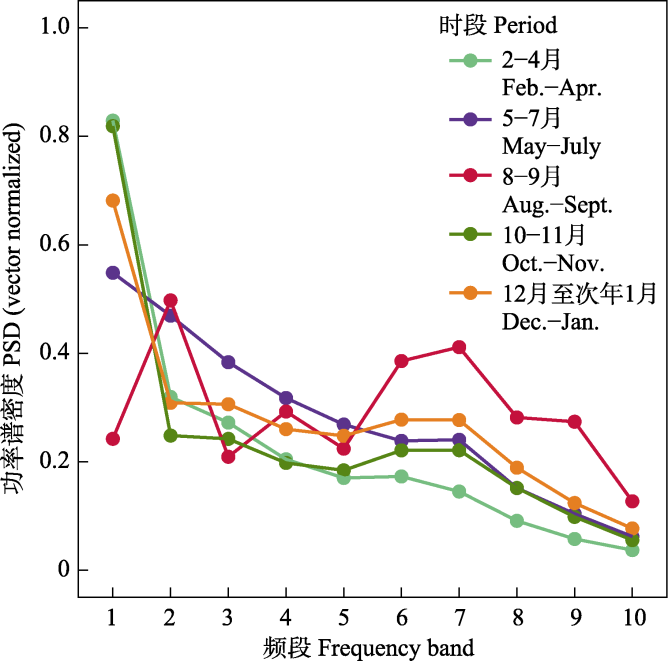 View image in article
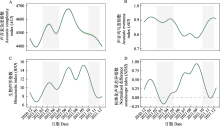
图4
五个时段的功率谱密度(PSD)归一化值。PSDi表示频段i的功率谱密度, 即i到(i + 1) kHz频段的功率谱密度。
正文中引用本图/表的段落
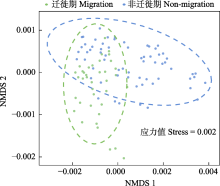
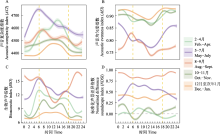
10台自动录音设备共采集了115,360条时长5 min的音频。根据风雨声信号的第二四分位数阈值共筛选出91,988条有效录音(附录1)。NMDS和ANOSIM分析结果表明, 鸟类迁徙期与非迁徙期声景格局间存在显著差异(stress = 0.002, R = 0.19, P = 0.001) (图3)。ACI、BIO、AEI、NDSI值在全年5个时段间均存在显著差异(所有P < 0.001, 表2)。Kruskal-Wallis检验结果显示迁徙期的NDSI显著低于非迁徙期(P < 0.001) (表2), 而功率谱密度曲线显示迁徙期的PSD1明显高于非迁徙期(图4), 表明迁徙鸟类显著增强了湿地声景中1-2 kHz频段的声信号。
2-4月, ACI和BIO呈现小的峰值(图5A、C), NDSI值为全年最低(图5D)但PSD1最高(图4), 此时声景中被占用的频段少, 由高振幅和窄频段(1-2 kHz频段)声信号主导。经人工识别, 1-2 kHz频段的声信号主要由旅鸟豆雁和白额雁等产生(以下统称雁类), 以及来自鹤类等其他类群少量的贡献(图2a)。
5-7月, ACI值快速升高并达到峰值(图5A), 同时, BIO、NDSI值也显著升高, AEI显著降低(表2, 图5), PSD1明显降低且PSD2-10均升高(图4), 这可能是因为1-2 kHz频段的雁类声信号减弱, 2-11 kHz频段的发声物种占用频段增多且频段间强度差异减小, 发声活动和复杂性显著增强(图2C)。经人工识别, 夜晚的2-3 kHz频段主要为蛙类声信号, 白天2-11 kHz频段的鸟类声信号丰富。
8-9月, PSD1降至最低水平, PSD2、4和6-9较高(图4), 表明声景主要由2-3 kHz、4-5 kHz和6- 10 kHz的声信号主导。NDSI显著高于其他时段并达到峰值, 且AEI显著低于其他时段(图5D, 表2), 说明2-11 kHz频段声信号最强, 占据的频段最多。BIO显著高于其他时段, 此时ACI最低, 代表此时生物声最丰富但鸟类的发声活动减弱, 经人工识别这一时段发出连续信号的昆虫种类多且发声活动强(图2D)。
10-11月, BIO较前一时段显著降低(图5C), AEI显著升高(图5B), 表明2-11 kHz频段上的生物信号明显减少, 声景均匀度降低且被较少频段主导。PSD1上升、PSD2-10下降(图4)、NDSI显著降低(图5D), 说明声景中1-2 kHz频段声信号又一次占主导, 以南迁雁类发声为主, 但是发声活动没有2-4月雁类北归时强。
12月至翌年1月, 进入冬季后, PSD1有所下降, 但仍处于较高水平(图4)。ACI和BIO处于全年较低水平(图5), 代表此时仅有少量生物发声活动。PSD2-10较上一时段均升高(图4), NDSI显著升高且均大于0 (图5D), 表明1-2 kHz频段声信号减弱, 2-11 kHz频段声信号增多。因此, 这段时间鸟类发声活动不强, 1-2 kHz雁类发声活动大幅减弱, 声景主要成分为较少的本地鸟类声信号。
声学指数的动态变化主要由鸟类和昆虫的声学活动变化驱动, 另外也经常受哺乳动物、两栖动物和爬行动物的影响(Diepstraten & Willie, 2021; Opaev et al, 2021)。本研究通过计算声学指数和功率谱密度, 对图们江下游湿地声景的时间格局进行了综合评价, 发现研究区声景主要受候鸟迁徙和蛙类、昆虫鸣叫的影响, 表现出不同的月变化和日变化规律。图们江下游湿地作为鸟类迁徙活动中的重要停歇地, 每年春季大量鸭科的雁鸭类迁徙至此, 不仅数量大而且种类多, 优势种包括豆雁、白额雁、绿头鸭、针尾鸭、斑嘴鸭和普通秋沙鸭等, 其他稀有种包括虎头海雕、白尾海雕、丹顶鹤、东方白鹳(Ciconia boyciana)等。雁类通常停歇时间较短, 约3-5天, 鸭类相对停歇时间较长, 大约10天, 4月中下旬雁鸭类群体陆续离开(杨兴家和吴志刚, 1994)。几个声学指数反映了这一迁徙变化过程, 春季随着候鸟的到来ACI和BIO逐渐升高, 在3月上旬达到一个小的峰值, 随着鸟类陆续离开这两个指数逐渐下降(图5)。然而, NDSI值呈现相反的趋势, 并在鸟类迁徙高峰期达到全年最低值, 这是因为根据声谱图(图2), 雁类的发声频段主要在1-2 kHz, 表明春季鸟类迁徙停歇期研究区的声景主要由雁类低频声信号主导, 功率谱密度变化曲线也证实了这一点, 此时1-2 kHz频段的功率谱密度最高(图4)。每年春季先后有数十万只雁类在敬信湿地农田、草丛、湖泊中停歇, 此时它们进入发情期, 将更多能量投入到求偶鸣唱活动, 特别是白天发声强度最大。Buxton等(2016)在美国阿拉斯加的研究也发现, 当候鸟从越冬地返回并开始繁殖时有一个明显的春季声学活动峰值。
本文的其它图/表
|
{kind=link}