



外来生物入侵已成为威胁全球生物多样性、生态安全和社会经济的重大环境问题之一(朱丽和马克平, 2010; Diagne et al, 2021)。随着全球经济一体化的飞速发展, 全球贸易、国际旅游等活动打破了物种扩散的天然地理屏障, 使其能够在不同地理和生态区域之间交叉转移(朱丽和马克平, 2010; Kumar & Singh, 2020)。如何准确地模拟入侵物种的空间分布范围对生物入侵的风险评估及防治措施的制定至关重要。在大尺度的空间转移过程中, 气候条件是决定外来物种能否形成入侵的主要因素(Pearson & Dawson, 2003)。外来物种在到达新生境的初期更容易在与其气候生态位相似的环境中形成入侵(Bustamante et al, 2020), 然而随着入侵时间的增加, 外来物种的生态位在空间和时间上是否具有保守性不仅成为生物入侵理论研究关注的焦点(Liu et al, 2020), 也是能否准确预测入侵物种分布范围的关键(朱丽和马克平, 2010)。一方面, 有学者认为大多数入侵物种的气候生态位具有高度的保守性(Liu et al, 2020), 尤其是对陆地入侵植物而言, 其气候生态位发生实质性变化的现象非常罕见(Petitpierre et al, 2012)。另一方面, 也有学者认为外来物种在入侵过程中其生态位的变化与其引进方式和跨越的地理范围相关, 人为引进的栽培物种与洲际入侵物种的生态位变化最为明显(Atwater et al, 2018), 并且由于区域环境的差异性, 同一入侵物种在不同区域间其生态位的变化具有显著的差异性(Christina et al, 2020)。
使用物种分布模型预测物种适宜分布区的一个关键假设是所研究物种的生态位在空间和时间上是保守的(朱丽和马克平, 2010)。有研究表明, 尽管大多数入侵物种的气候生态位具有保守性(Liu et al, 2020), 但仍有部分入侵物种在到达新生境后摆脱了制约其在原产地种群分布的限制因素(如, 扩散限制、竞争、捕食), 并且入侵物种本身较高的生态可塑性以及到达新生境后面临新的自然选择过程(Müller-Schärer et al, 2004), 均有可能使得入侵物种在入侵区域产生适应性的生态位漂移(王运生等, 2008)。如薇甘菊(Mikania micrantha)在入侵至泰国、尼泊尔等地后倾向于向干旱的生境扩散, 而入侵至印度尼西亚的种群更倾向于向寒冷的生境扩散(Banerjee et al, 2019)。因此, 在模拟入侵物种空间分布格局时应充分考虑其生态位的变化。此外, 入侵物种的扩散能力对于理解和预测物种扩散动态也至关重要(Thuiller et al, 2008), 在入侵物种适宜分布区的模拟中, 即使只粗略地考虑物种的扩散能力, 也可以明显降低模型预测的不确定性(Engler & Guisan, 2009)。因此, 在使用物种分布模型预测入侵物种的适宜分布区之前, 评估入侵物种生态位在时间和空间上的变化特征, 在预测未来气候变化情景下入侵物种的适宜分布范围时加入物种扩散能力参数, 不仅有助于理解入侵物种的生态位变化趋势, 还可以更真实地预测物种的扩散动态与入侵能力。
紫茎泽兰(Ageratina adenophora)是菊科紫茎泽兰属(Ageratina)多年生草本或半灌木植物, 原产于墨西哥。因其具有极强的繁殖能力、传播能力、适应能力、侵占能力和抗逆性, 现已广泛分布于热带、亚热带地区的30多个国家, 严重干扰当地的生物多样性、破坏当地生态平衡。并且由于其枝叶有毒, 给入侵地区的人类及牲畜健康造成了严重的威胁, 成为世界公认的恶性有毒入侵植物(强胜, 1998)。紫茎泽兰自1935年在我国云南南部首次发现以来(鲁萍等, 2005), 经过几十年的适应、传播, 已扩散至云南、四川、西藏、广西、贵州、湖北等地, 其分布的海拔上限也从最初的1,800 m扩大至2,500 m (Wang & Wang, 2006; 土艳丽等, 2018), 并且在未来有进一步向高海拔区域迁移的趋势(Gu et al, 2021)。过去十几年里有关紫茎泽兰的研究主要集中于其生物学特性、入侵机制、潜在分布及防除等方面(王翀等, 2014; 李霞霞等, 2017)。然而, 紫茎泽兰在入侵我国后其气候生态位的时空变化格局如何? 过去几十年该植物在我国的空间分布格局以及在考虑物种扩散能力的前提下, 未来气候变化情景下该物种在我国的入侵风险如何尚不明确。本研究基于紫茎泽兰在我国及其原产地的分布点及气候数据, 通过生态位分析及物种分布模型模拟, 旨在探讨: (1)紫茎泽兰在入侵至我国后其气候生态位的时空变化特征; (2)紫茎泽兰在我国的潜在分布范围及主要的限制因子; (3)在物种扩散能力的约束下, 预测紫茎泽兰在未来气候变化情景下的扩散动态。
1 材料与方法
1.1 物种分布记录和环境变量
紫茎泽兰在我国的分布信息来源于相关文献记录(附录1)和国家标本资源共享平台(NSII,
一般来说, 非生物的气候和地形因素被认为是影响入侵物种在大尺度上分布的关键因素。此外, 物种相互作用的强度也是影响入侵物种空间分布的重要因素之一(朱耿平等, 2014)。将生物相互作用加入到物种分布模型中可以更好地模拟入侵物种的空间分布格局(Meier et al, 2010)。有研究表明, 在区域和大陆尺度上, 植被生产力可能与生物相互作用的强度呈正相关(Early & Keith, 2019), 因此本研究使用平均净初级生产力(net primary productivity, NPP)数据代替研究区内的生物相互作用(Quiroga et al, 2021)。最终本研究选择气候、地形及代替生物相互作用的NPP 3种数据用于模拟紫茎泽兰在我国的时空分布格局, 其中生物气候变量(1970-2000)与海拔变量均来自WorldClim (
1.2 气候生态位分析
ecospat软件包提供的基于主成分分析(principal component analysis, PCA)的方法可以在连续的空间中分析物种的生态位, 已被广泛应用于外来入侵物种生态位动态的研究中(di Cola et al, 2017; Liu et al, 2020)。本研究基于紫茎泽兰的分布数据及9个生物环境变量, 首先基于紫茎泽兰在原产地与入侵地的物种分布点构建最小凸多边形(minimum convex polygon, MCP), 然后对该最小凸多边形按照该物种每年平均扩散距离(20 km)做缓冲处理, 用于空间比对及模型构建和评估(范靖宇等, 2019)。最后使用ecospat软件包量化紫茎泽兰在入侵至我国后其气候生态位的时空变化特征, 为消除边缘气候对结果的影响, 使用75%的置信区间作为消除边缘气候的阈值(Duchoslav et al, 2020)。在分析时, 首先对原产地与入侵地的分布记录进行随机分配, 通过生态位等价性检验(niche equivalency tests)来计算紫茎泽兰生态位重叠区域是否恒定。其次通过生态位相似性检验(niche similarity tests)计算紫茎泽兰在原产地与入侵地的生态位重叠区域是否高于其中一个地区生态位与从另一个区域随机选择的生态位之间的重叠区域, 两种检验均使用100次重复(孙燕等, 2017)。最后通过生态位稳定性(niche stability, NS, 在入侵地区和原产地都出现的生态位空间)及生态位扩张(niche expansion, NE, 仅出现在入侵地区的生态位空间), 评估该物种在原产地与入侵地的气候生态位差异。
1.3 物种分布模型的构建与评估
最大熵模型(MaxEnt)因其仅依据存在数据就能推测环境变量下最大熵的可能分布, 具有运算速度快、预测结果好等优点, 成为目前应用最广泛的物种分布模型(Yu et al, 2014; Phillips et al, 2017)。本研究利用ENMeval 2.0优化包调用MaxEnt模型建立最优模型进行紫茎泽兰适宜分布区的模拟(Kass et al, 2021)。为提高模型结果的准确性与模型转移能力, 本研究通过优化调控倍频(regularization multiplier, RM)和特征组合(feature combination, FC)两个参数选择最优模型用于紫茎泽兰分布区的模拟(朱耿平和乔慧捷, 2016), 选择ΔAICc = 0 (Akaike information criterion correction)的模型作为最优模型(Muscarella et al, 2014)。在模型构建过程中, 利用dismo程序包中的randomPoints函数生成1,000个伪存在点数据用于模型模拟, 选取75%的样点数据用作模型训练, 剩余25%的样点数据用于模型验证并进行10次重复。利用受试者操作特征曲线下面积(the area under the receiver operating characteristic curve, AUC)评估模型结果的准确性。AUC的取值范围为[0, 1], 值越大表示模型的准确性越高, AUC > 0.5表示模型的模拟效果好于随机, AUC ≥ 0.9表示模型的模拟效果极好(胡忠俊等, 2015)。模型结果使用cloglog作为输出格式, 因为cloglog变换更适合于存在概率的估算, 比logistic变换具有更强的理论依据(Phillips et al, 2017)。最后使用dismo软件包中的threshold函数选择特异性和敏感性之和最大时(maximum training sensitivity plus specificity, MaxSS)的阈值作为紫茎泽兰适宜分布与不适宜分布二元分类结果的划分阈值。多元环境相似度面(MESS)和最不相似变量(MoD)分析用于评估不同区域气候相似性及最不相似变量。MESS值为负值时, 说明该点至少一个环境变量的值超出了参考范围, 提取相似度最低的气候因子, 得到最不相似变量(张丹等, 2022)。
1.4 考虑扩散能力预测未来气候变化情景下紫茎泽兰的分布范围
选择2021-2040时段下SSP1-2.6情景模拟紫茎泽兰未来在我国的潜在分布范围, 并基于MigClim软件包加入紫茎泽兰的扩散能力作为约束条件, 模拟其在未来气候变化情景下的扩散动态。有研究表明自然条件下紫茎泽兰在过去几十年里的最小、最大和平均扩散速度分别为5.8、26.6和16.6 km/年(Wang & Wang, 2006), 也有研究表明极端气候条件下紫茎泽兰的扩散速度最高可达60 km /年(杨蓉西, 2003)。本研究基于短距离扩散模型与长距离扩散模型相结合的方法模拟紫茎泽兰在未来的扩散动态(Engler et al, 2012)。几个重要参数设置如下: (1)紫茎泽兰的初始分布(iniDist): 本研究使用紫茎泽兰在1971-2020年间所有的分布点作为初始分布数据。(2)扩散内核(dispKernel): 物种繁殖体到达不同空间距离的概率。植物种子到达不同空间距离的概率随距离的增加呈负指数分布, 因此本研究中该参数基于如下公式计算(Engler & Guisan, 2009):
其中, Pseed表示植物种子到达空间距离
以上数据处理、物种潜在分布区建模及生态位分析均基于R 3.6.3 (R Core Team, 2020)完成。
2 结果
2.1 紫茎泽兰在中国的气候生态位及其时空变化
生态位分析结果表明, 在时间尺度上, 紫茎泽兰在中国的气候生态位与其原产地的气候生态位高度重叠(图1), 并且与原产地的气候生态位相比具有很高的稳定性(NS > 0.85, 表1)。尤其是在入侵初期(1980年之前), 气候生态位与其在原产地的气候生态位基本一致且生态位稳定性指数最高(NS = 0.995, 图1, 表1)。随着入侵时间的增加, 紫茎泽兰在中国的气候生态位逐渐变宽且发生了低水平的生态位扩张(NE < 0.141, 图1, 表1), 到2020年, 紫茎泽兰在中国的气候生态位扩张指数达最大(NE = 0.141, 表1)。具体表现为: 随着时间的推移该物种在中国更倾向于潮湿(高最暖季降水量)、寒冷(低最干季平均温)且昼夜温差小(低平均日较差)的生境(图2), 并且超出了其在原产地已有的气候生态位。在空间上, 分布在西藏、广西和重庆的紫茎泽兰所占据的气候生态位仍与其在原产地的气候生态位重叠且生态位稳定性最高, 但是分布在云南、贵州和四川的紫茎泽兰所占据的气候生态位与其原产地的气候生态位相比略有扩张, 尤其是分布在纬度最高的四川的种群其生态位扩张最大(NE = 0.168, 图1, 表2)。
图1
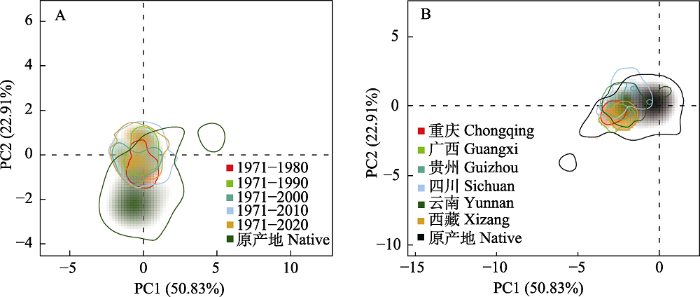
图1
不同时间(A)及空间(B)上紫茎泽兰在我国的气候生态位的变化特征
Fig. 1
Climate niche dynamics of Ageratina adenophora in China in different temporal (A) and spatial (B)
表1 不同时期入侵地与原产地紫茎泽兰气候生态位的稳定性与生态位扩张
Table 1
| 1971-1980 | 1971-1990 | 1971-2000 | 1971-2010 | 1971-2020 | |
|---|---|---|---|---|---|
| 生态位稳定性 Niche stability (NS) | 0.995 | 0.953 | 0.909 | 0.881 | 0.859 |
| 生态位扩张 Niche expansion (NE) | 0.005 | 0.047 | 0.091 | 0.119 | 0.141 |
图2
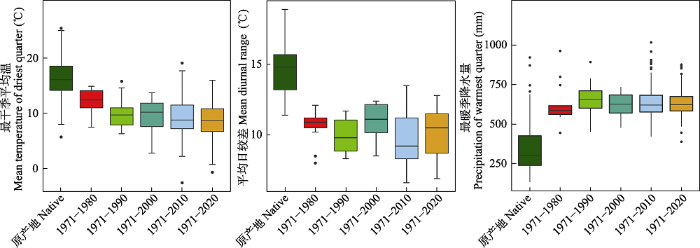
图2
紫茎泽兰在原产地与我国主要的生物气候因子差异
Fig. 2
Differences of the three bioclimatic variables of Ageratina adenophora between native and China
表2 与原产地相比不同入侵区域紫茎泽兰气候生态位的稳定性与生态位扩张
Table 2
| 云南 Yunnan | 广西 Guangxi | 贵州 Guizhou | 四川 Sichuan | 重庆 Chongqing | 西藏 Xizang | |
|---|---|---|---|---|---|---|
| 生态位稳定性 Niche stability (NS) | 0.959 | 0.990 | 0.902 | 0.832 | 1.000 | 1.000 |
| 生态位扩张 Niche expansion (NE) | 0.041 | 0.010 | 0.098 | 0.168 | 0 | 0 |
2.2 紫茎泽兰的适宜分布区变化及主要限制因子
基于ΔAIC = 0的标准选择最优模型用于不同时期紫茎泽兰适宜分布区的模拟, 结果表明所有模型的AUC值均大于0.93 (标准差SD ≤ 1.16), 说明模型预测结果准确且不同重复之间的稳定性较好(附录4)。
根据模型预测结果, 紫茎泽兰在中国的适宜分布范围逐年增加(图3, 附录5), 其中1980年之前, 紫茎泽兰在中国的适宜分布范围主要集中在云南、四川南部、海南西南部及西藏南部的零星区域, 总面积约241,730.5 km2 (图4, 附录5)。1980年以后紫茎泽兰开始向东北扩散至贵州西南及四川北部等地, 并且在西藏的适宜分布区也逐渐增加(图3, 4)。到2020年紫茎泽兰的适宜分布范围达到最大, 面积约为655,434.98 km2 (图4, 附录5), 主要分布在云南大部分、四川中部及南部、贵州西南部及广西西部(图4)。2010年之前紫茎泽兰在中国的适宜分布区的重心一直表现为向东北迁移的特点, 但是2010年以后呈现出向西南迁移的趋势(图4)。
图3
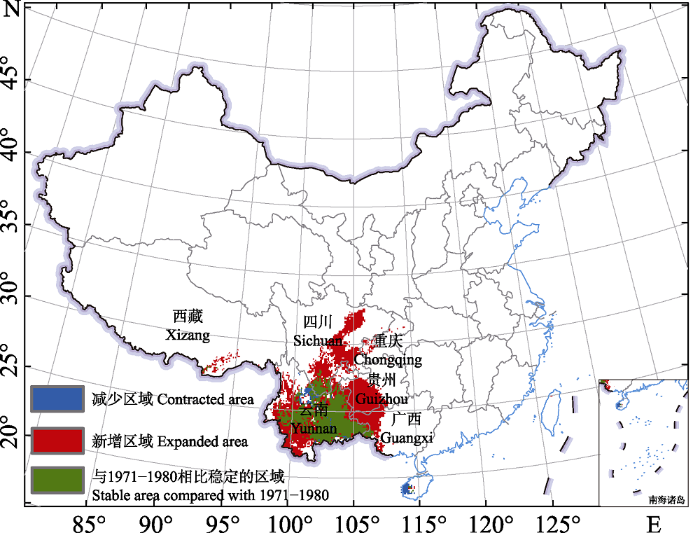
图3
过去50年紫茎泽兰潜在分布整体变化趋势
Fig. 3
Total change trend of potential distribution of Ageratina adenophora in the past 50 years
图4
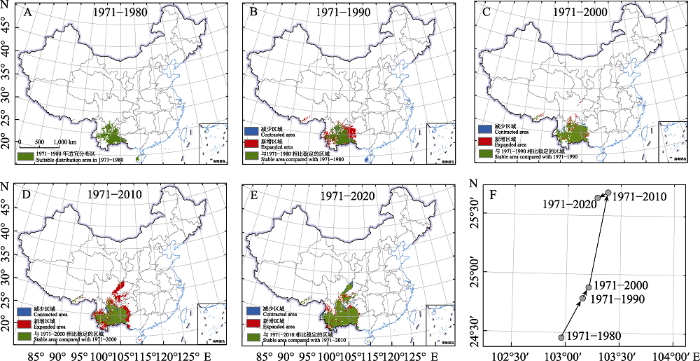
图4
不同时期紫茎泽兰在中国的潜在分布(A-E)及其重心变化(F)
Fig. 4
Potential distribution (A-E) and center of gravity (F) of Ageratina adenophora change during different periods in China
图5
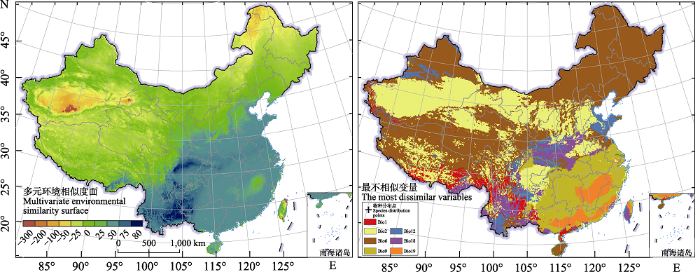
图5
影响紫茎泽兰分布的7个生物气候变量的多元环境相似度面(MESS)和最不相似变量(MoD)。
Bio 1: 年平均温度; Bio 2: 平均日较差; Bio 6: 最冷月最低温; Bio 9: 最干季平均温; Bio 12: 年平均降水量; Bio 18: 最暖季降水量; Bio 19: 最冷季降水量。
Fig. 5
Multivariate environmental similarity surface (MESS), the most dissimilar variable (MoD) of seven bioclimatic variables that affect the distribution of Ageratina adenophora.
Bio 1, Annual mean temperature; Bio 2, Mean diurnal range; Bio 6, Min temperature of coldest month; Bio 9, Mean temperature of driest quarter; Bio 12, Annual precipitation; Bio 18, Precipitation of warmest quarter; Bio 19, Precipitation of coldest quarter.
2.3 未来气候变化对紫茎泽兰潜在入侵风险的可能影响
紫茎泽兰在2021-2040时段下的适宜分布范围将进一步增加(约增加78,458.81 km2, 附录5)。具体而言, 与目前的适宜分布范围相比, 紫茎泽兰新增的大片适生区主要分布在四川东北部、重庆南部、贵州西部等区域。而且从目前的扩散速度来看, 未来的20年中, 紫茎泽兰能够扩散到其绝大部分潜在适生区, 只有西藏东南部区域因受到扩散限制而无法达到(图6)。
图6
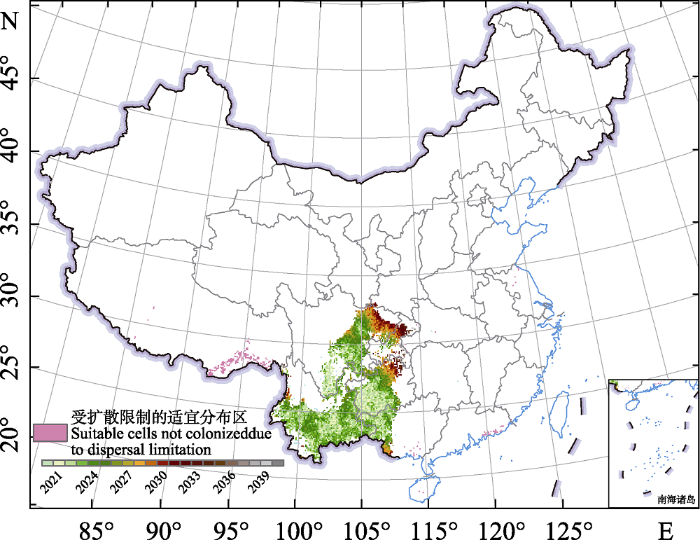
图6
紫茎泽兰在2021-2040年的潜在扩散动态
Fig. 6
Potential dispersal dynamics of Ageratina adenophora from 2021 to 2040
3 讨论
3.1 紫茎泽兰入侵我国后气候生态位虽略有扩张但整体保持较高的稳定性
本研究结果表明, 紫茎泽兰入侵到我国初期(1980年之前), 其气候生态位与原产地相比表现出高度的保守性, 随着时间的推移其气候生态位虽略有扩张, 但仍保持较高的稳定性(NS > 0.83, 图1, 表1, 2), 这与紫茎泽兰在亚洲气候生态位稳定性较高的研究结果相似(Datta et al, 2019), 也与目前大多数入侵植物的生态位具有保守性的研究结果一致(Liu et al, 2020)。同时由于本研究在分析紫茎泽兰气候生态位变化时并未考虑气候变化的影响, 而事实上过去几十年中我国西南地区呈暖干化趋势(苏秀程等, 2014), 因此紫茎泽兰在我国气候生态位的稳定性可能更高。对大多数外来物种而言, 形成入侵是一个引入-归化-入侵的连续过程, 在初始阶段通常是在与其原产地气候相似的地区形成入侵 (Banerjee et al, 2019), 因为只有在与其原产地气候条件相似的生境中才更有利于其建立能够自我维持的种群(朱丽和马克平, 2010), 并且这个过程往往要持续数十年时间(鞠瑞亭等, 2012)。在这段时间内入侵物种的生态位往往表现出高度的保守性, 同时也是入侵物种在形态、物候等方面产生适应性进化的重要时期, 随后便进入种群暴发和入侵范围迅速扩张阶段(Dietz & Edwards, 2006; 朱丽和马克平, 2010)。然而紫茎泽兰是无融合生殖的三倍体植物, 产生适应性进化的能力较低(Bala & Gupta, 2014), 并且在对云南紫茎泽兰种群遗传结构的研究中并未发现本地适应的基因型(宋紫玲, 2019 ( 宋紫玲 (2019) 道路廊道和森林景观格局对紫茎泽兰种群及其遗传结构的影响. 硕士学位论文, 云南大学, 昆明.)), 也就是说紫茎泽兰可能并未发生适应性的遗传变异而扩展其生态位, 因此其在扩散过程中仍保持了较高的生态位稳定性。
3.2 自入侵以来紫茎泽兰在我国的适生区逐渐扩大且主要表现出向东、向北的迁移趋势
紫茎泽兰自中缅边境传入我国云南以后其适宜分布范围显著增加(图4, 附录4), 1990年之前主要分布在我国云南、四川南部、贵州西南部及广西西部, 随后持续向北及东北继续扩散, 到2010年已经广泛分布在云南除西北以外的绝大部分地区、四川南部、重庆西南部、贵州西南部及广西西部, 且其分布区的最北端延伸到四川北部(图4)。在扩散方向与扩散时间上, 本研究的结果与以往对紫茎泽兰入侵扩散过程的研究结果基本一致, 如1998年金沙江-长江特大洪水促进了紫茎泽兰在川渝地区的扩散, 使得其沿金沙江-长江向东北方向扩散, 到2000年后扩散至重庆西南部的屏山县、宜宾市及中部的长寿县等地。在分布范围上, 本研究预测的紫茎泽兰的适生区范围略大于前人研究的潜在分布范围(王翀等, 2014), 如海拔较高的四川南部及贵州北部。这可能是由于前期有关紫茎泽兰的分布数据较少, 模型对于该物种气候生态位的考虑不足,导致其模拟的适生区范围较小, 事实上本研究也发现在向高纬度扩散的过程中, 紫茎泽兰对低温的耐受性也随之扩大, 尤其是分布在纬度最高的四川的种群其生态位扩张最为明显(图1, 表2)。紫茎泽兰目前的分布区域主要受西南季风控制, 常年盛行西南风, 而其种子细小并且顶端具冠毛适于风力传播(Zhu et al, 2020), 因此主要表现出向东北扩散的特点。然而自2010年以后该物种的潜在分布区重心开始向西南迁移(图3), 多元环境相似度分析表明温度条件, 尤其是最冷月最低温, 可能是限制紫茎泽兰继续向北扩散的主要因素(图4)。紫茎泽兰喜温暖湿润的环境且对低温敏感, 在全球范围内其适宜生境的最低温通常是在‒11℃以上(Datta et al, 2017), 而青藏高原及中国北方地区年平均最低温往往低于这个值, 因此限制了紫茎泽兰向北进一步扩散。除上述地区外, 西藏南部地区也有该物种孤立的、小范围的适生区, 主要分布在吉隆沟、樟木沟及藏南地区, 但是由于该区域内地形复杂且多高山, 而西藏腹地的低温环境也不适合紫茎泽兰的生存, 因此限制了紫茎泽兰在西藏的扩散。已有研究表明分布在西藏的紫茎泽兰是通过吉隆樟木口岸从尼泊尔传入, 而并非从云南扩散而来。本研究中也并未在西藏、云南、四川三省交界区域检测到紫茎泽兰的适生区, 这也进一步佐证了分布在西藏的紫茎泽兰种群不是从云南扩散而来的结论(土艳丽等, 2018)。
3.3 未来紫茎泽兰的分布范围可能进一步扩大且有能力扩散至整个适生区
未来气候变化情景下, 紫茎泽兰在我国的适宜分布范围将进一步增加, 增幅约为111.97%, 如四川东北部、重庆南部、贵州北部及广西西部(图5), 到2040年紫茎泽兰基本可以扩散至其整个适宜分布区(图6)。一方面是因为紫茎泽兰具有极强的繁殖能力、能够依赖多种媒介(如风、水、动物)进行传播且其扩散速度极快(强胜, 1998; 杨蓉西, 2003; Wang & Wang, 2006), 使得其能够迅速扩散至适宜分布区; 另一方面是因为紫茎泽兰在我国所占据的气候生态位没有达到饱和(Wang & Wang, 2006; 王瑞, 2006, 图1), 华南及中南地区的气候条件本就十分适合紫茎泽兰生长, 加上未来我国南方的气候表现出气温升高、降水增加的模式(向竣文等, 2021), 也为紫茎泽兰的入侵提供了良好的生境条件。未来气候变化情景下紫茎泽兰受扩散限制无法达到的适宜分布区主要是分布在藏南地区的小范围区域(图6), 这可能是因为目前紫茎泽兰在西藏的分布范围十分集中且主要分布在河谷当中(土艳丽等, 2018), 山脉的阻隔限制了紫茎泽兰在这些区域的扩散, 再加上西藏地区海拔较高, 由于海拔升高所引起的垂直地带性气候变冷也进一步限制了紫茎泽兰在该区域的扩散(王瑞, 2006)。本研究模拟紫茎泽兰未来扩散动态时使用的可持续发展情景是较为理想的增温情景, 事实上未来全球增温可能高于该情景的设定, 因此在未来气候变化情景下低温对紫茎泽兰的限制也会进一步释放, 其在我国的扩散可能比本研究预测的更为严重。
基于物种分布模型预测入侵物种的分布范围是入侵物种防治的重要手段, 明确入侵物种分布范围变化及其扩散趋势可以警示入侵物种可能会在哪些地区入侵并造成危害, 尤其模拟入侵发生初始阶段其潜在分布范围对于入侵物种控制措施的制定至关重要。本研究也发现生物入侵的初始阶段其气候生态位具有高度的保守性, 因此在早期可以更准确地预测入侵物种的分布范围, 进而为入侵物种的防治提供参考。然而随着入侵时间的增加, 入侵物种的生态位是否仍具有保守性目前尚处于争论之中, 因此, 在使用物种分布模型预测入侵物种潜在分布范围之前, 对目标物种生态位的时空动态格局进行检验不仅有助于对入侵物种生态位变化的理解, 还可以更准确地预测其分布范围。此外, 在使用物种分布模型时尽可能地考虑物种的扩散能力, 不仅可以细化入侵物种未来的扩散趋势, 而且可为濒危珍稀物种保护区的优化提供参考。
附录 Supplementary Material
附录1 紫茎泽兰在我国分布点的经纬度及其来源
Appendix 1 Longitude, latitude and reference of the distribution points of Ageratina adenophora in China
附录2 不同时期紫茎泽兰分布点空间分布及数量
Appendix 2 Spatial distribution and number fof distribution points of Ageratina adenophora in different periods
附录3 用于紫茎泽兰生态位建模的7个生物气候因子及其相关系数
Appendix 3 Pearson’s correlation coefficients (r) of the seven bioclimatic variables for the niche modeling of Ageratina adenophora
附录4 不同时期用于紫茎泽兰入侵分布区模拟的最佳模型参数及AUC值
Appendix 4 The optimal model parameters and AUC values for predicting the potential distribution regions of Ageratina adenophora in different periods
附录5 不同时段紫茎泽兰的潜在分布面积
Appendix 5 Potential distribution area of Ageratina adenophora in different periods
附录6 不同时期各环境因子对紫茎泽兰潜在分布范围的贡献率
Appendix 6 Contribution rate of all the environmental variables to the potential distribution area of Ageratina adenophora in different periods
致谢
特别感谢中国科学院地理科学与资源研究所王兆锋老师对数据分析提出的宝贵意见, 石河子大学马松梅老师及张丹对文章修改提出的宝贵建议, 以及新疆大学李晓辰博士对数据收集提供的帮助。
参考文献
spThin: An R package for spatial thinning of species occurrence records for use in ecological niche models
DOI:10.1111/ecog.01132 URL [本文引用: 1]
Climatic niche shifts are common in introduced plants
Meiotic behaviour in two triploids: Erigiron annus and Eupatorium adenophorum (Asteraceae)
Spatio-temporal patterns of climatic niche dynamics of an invasive plant Mikania micrantha Kunth and its potential distribution under projected climate change
DOI:10.3389/fevo.2019.00291 URL [本文引用: 2]
A classification system for predicting invasiveness using climatic niche traits and global distribution models: Application to alien plant species in Chile
DOI:10.3897/neobiota.63.50049 URL [本文引用: 1]
Climatic niche shift of an invasive shrub (Ulex europaeus): A global scale comparison in native and introduced regions
DOI:10.1093/jpe/rtz041 URL [本文引用: 1]
ecospat: An R package to support spatial analyses and modeling of species niches and distributions
DOI:10.1111/ecog.02671 URL [本文引用: 1]
Processes affecting altitudinal distribution of invasive Ageratina adenophora in western Himalaya: The role of local adaptation and the importance of different life-cycle stages
Niche expansion of the invasive plant species Ageratina adenophora despite evolutionary constraints
High and rising economic costs of biological invasions worldwide
DOI:10.1038/s41586-021-03405-6 URL [本文引用: 1]
Recognition that causal processes change during plant invasion helps explain conflicts in evidence
DOI:10.1890/0012-9658(2006)87[1359:RTCPCD]2.0.CO;2 URL [本文引用: 1]
Intricate distribution patterns of six cytotypes of Allium oleraceum at a continental scale: Niche expansion and innovation followed by niche contraction with increasing ploidy level
DOI:10.3389/fpls.2020.591137 URL [本文引用: 1]
Geographically variable biotic interactions and implications for species ranges
DOI:10.1111/geb.12861
[本文引用: 1]

The challenge Understanding how biotic interactions affect species' geographical ranges, biodiversity patterns and ecological responses to environmental change is one of the most pressing challenges in macroecology. Extensive efforts are underway to detect signals of biotic interactions in macroecological data. However, efforts are limited by bias in the taxa and spatial scale for which occurrence data are available and by difficulty in ascribing causality to co-occurrence patterns. Moreover, we are not necessarily looking in the right places; analyses are largely ad hoc, depending on availability of data, rather than focusing on regions, taxa, ecosystems or interaction types where biotic interactions might affect species' geographical ranges most strongly. Unpicking biotic interactions We suggest that macroecology would benefit from the recognition that abiotic conditions alter two key components of biotic interaction strength: frequency and intensity. We outline how and why variation in biotic interaction strength occurs, explore the implications for species' geographical ranges and discuss the challenges inherent in quantifying these effects. In addition, we explore the role of behavioural flexibility in mediating biotic interactions potentially to mitigate impacts of environmental change. New data We argue that macroecology should take advantage of "independent" data on the strength of biotic interactions measured by other disciplines, in order to capture a far wider array of taxa, locations and interaction types than are typically studied in macroecology. Data on biotic interactions are readily available from community, disease, microbial and parasite ecology, evolution, palaeontology, invasion biology and agriculture, but most are yet to be exploited within macroecology. Integrating biotic interaction strength data into macroecology Harmonization of data across interdisciplinary sources, taxa and interaction types could be achieved by breaking down interactions into elements that contribute to frequency and intensity. This would allow quantitative biotic interaction data to be incorporated directly into models of species distributions and macroecological patterns.
MigClim: Predicting plant distribution and dispersal in a changing climate
DOI:10.1111/j.1472-4642.2009.00566.x URL [本文引用: 2]
The MIGCLIM R package-seamless integration of dispersal constraints into projections of species distribution models
DOI:10.1111/j.1600-0587.2012.07608.x URL [本文引用: 1]
Selecting the best native individual model to predict potential distribution of Cabomba caroliniana in China
DOI:10.17520/biods.2018232 URL [本文引用: 1]
基于本土最优模型模拟入侵物种水盾草在中国的潜在分布
DOI:10.17520/biods.2018232
[本文引用: 1]
生态位模型预测存在不确定性, 不同模型预测结果差别较大。在生态位保守的前提下, 在本土区域构建经典生态位模型, 利用入侵地独立样本数据检验并选择最优模型, 具有独特优势, 可为入侵物种风险分析提供可靠参考。水盾草(Cabomba caroliniana)是一种恶性水生入侵杂草, 原产于南美洲, 已在我国多个省市建立种群, 本文基于本土最优模型预测其在我国的潜在分布, 以期为其风险分析和综合治理提供依据, 并通过水盾草案例探讨如何提高生态位模型预测准确性的方法。本文按时间顺序梳理了水盾草在我国的分布记录, 然后根据水盾草已有分布记录和其所关联的环境因子比较了不同地理种群所占有的气候生态空间, 测试水盾草在世界入侵过程中的现实生态位保守性。采用两组环境变量和5种算法在南美洲本土地区构建10种生态位模型, 并将其转移至我国, 基于最小遗漏率和记账错率, 利用我国(入侵地)的样本数据选择最优模型预测水盾草在我国的适宜生态空间和潜在分布。研究发现当前水盾草在我国的分布集中在东部水域充沛地区, 沿京杭运河和南水北调工程等向北扩散。生态空间比对中发现水盾草在亚洲与其他大洲所占有的生态空间具有一定的重叠, 其在我国的入侵过程中生态位是保守的。与本土空间相比, 水盾草在我国所占有的生态空间存在较大的生态位空缺, 表明水盾草在我国的潜在分布范围较大。生态位模型预测显示水盾草的适生区主要分布于我国的北京、上海、山东、浙江、江苏、安徽、湖北和湖南等省(市)。水盾草的潜在分布区多聚集在我国东南部, 该地区河流、湖泊、运河和渠道较为密集, 人类活动及自然天敌的缺乏容易助长其入侵趋势, 应在这些适宜地区开展调查, 及时发现疫情并采取相应措施。
Predicting the potential global distribution of Ageratina adenophora under current and future climate change scenarios
DOI:10.1002/ece3.7974 URL [本文引用: 1]
Cross-validation of species distribution models: Removing spatial sorting bias and calibration with a null model
Species distribution models are usually evaluated with cross-validation. In this procedure evaluation statistics are computed from model predictions for sites of presence and absence that were not used to train (fit) the model. Using data for 226 species, from six regions, and two species distribution modeling algorithms (Bioclim and MaxEnt), I show that this procedure is highly sensitive to "spatial sorting bias": the difference between the geographic distance from testing-presence to training-presence sites and the geographic distance from testing-absence (or testing-background) to training-presence sites. I propose the use of pairwise distance sampling to remove this bias, and the use of a null model that only considers the geographic distance to training sites to calibrate cross-validation results for remaining bias. Model evaluation results (AUC) were strongly inflated: the null model performed better than MaxEnt for 45% and better than Bioclim for 67% of the species. Spatial sorting bias and area under the receiver-operator curve (AUC) values increased when using partitioned presence data and random-absence data instead of independently obtained presence-absence testing data from systematic surveys. Pairwise distance sampling removed spatial sorting bias, yielding null models with an AUC close to 0.5, such that AUC was the same as null model calibrated AUC (cAUC). This adjustment strongly decreased AUC values and changed the ranking among species. Cross-validation results for different species are only comparable after removal of spatial sorting bias and/or calibration with an appropriate null model.
Simulation of Stipa purpurea distribution pattern on Tibetan Plateau based on MaxEnt model and GIS
基于MaxEnt模型和GIS的青藏高原紫花针茅分布格局模拟
气候变化对物种分布的影响是生物地理学研究的热点问题.本研究以广泛分布在青藏高原高寒草甸区域的建群种紫花针茅为研究对象,通过植物标本库查询和实地调查了解紫花针茅在青藏高原的分布情况,利用MaxEnt模型模拟紫花针茅在青藏高原历史、当前及未来的分布趋势,分析紫花针茅各历史时期的分布格局,探讨了物种分布变化的产生原因.结果表明: 青藏高寒草地的针茅属物种具有较高的物种多样性,代表性植物紫花针茅的主要分布区域为青藏高原腹地及沿喜马拉雅地区;紫花针茅的分布强烈地受最暖季降水量、最湿季降水量和年均降水量的控制.依据紫花针茅在末次冰期的分布格局以及青藏高原的地理地质特点,研究认为: 冰期来临时,羌塘以南藏北核心区和喜马拉雅西部阿里地区是紫花针茅潜在分布的核心区,此处提供了比其他地区更适合紫花针茅生存的栖息地,这些区域成为紫花针茅的避难所,当前的紫花针茅都是从这些避难所迁移并扩展开来.生物避难所的存在有助于理解青藏高原高寒植被起源和分化的相关问题.
Climate Change 2021:The Physical Science Basis
Progress of biological invasions research in China over the last decade
DOI:10.3724/SP.J.1003.2012.31148 URL [本文引用: 1]
近十年中国生物入侵研究进展
DOI:10.3724/SP.J.1003.2012.31148
[本文引用: 1]
生物入侵已对入侵区的生态环境、社会经济和人类健康造成了严重的威胁, 成为了21世纪五大全球性环境问题之一。本文回顾了2000年以来, 中国生物入侵研究领域尤其是入侵种的多样性与格局、入侵机制及生态学效应、管理与控制等方面所取得的重要进展, 讨论了需进一步加强研究的领域, 以期为进一步拓展该领域研究的广度和深度、为我国的生物入侵预警预防和科学治理提供参考。据初步研究, 中国的入侵种数量已达529种, 其中陆生植物、陆生无脊椎动物和微生物为主要入侵类群; 原产地以北美洲和南美洲为主; 经济发达和气候温暖湿润的东部和南部省份入侵态势明显较西部和北部省份严重; 随着中国经济的进一步发展, 生物入侵问题将可能更加严峻。外来种的成功入侵是其内禀优势、资源机遇和人为干扰共同作用的结果; 其中, 表型可塑性、适应性进化、天敌释放、种间互利或偏利共生和新化感作用等因素对入侵起到了关键作用。生物入侵已对中国土著生态系统的生物多样性和生态系统服务功能造成了严重影响, 打破了生态系统的固有平衡, 危害或威胁到中国的农林牧渔业生产、交通航运、环境、人类健康和公共设施安全。针对生物入侵的管理与控制, 中国加强了包括检测监测、风险分析、生物防治、扩散阻断、根治灭除和生态恢复等技术体系的研究和实施, 并初步控制了一些重要入侵种的扩张。中国生物入侵需要在全境性科学考察、生物入侵的遗传学、基因组学、生态系统影响、全球变化和管理与控制技术创新等领域进一步加强跨领域的交叉合作和系统研究。
ENMeval 2.0: redesigned for customizable and reproducible modeling of species’ niches and distributions
DOI:10.1111/2041-210X.13628 URL [本文引用: 1]
Invasive alien plant species: Their impact on environment, ecosystem services and human health
DOI:10.1016/j.ecolind.2019.106020 URL [本文引用: 1]
Progress of the research on invasive plant species Eupatorium adenophorum over the last decade
近十年入侵植物紫茎泽兰研究进展
Most invasive species largely conserve their climatic niche
Progress and prospects in research of an exotic invasive species, Eupatorium adenophorum
外来入侵种紫茎泽兰研究进展与展望
DOI:10.17521/cjpe.2005.0128
[本文引用: 1]
紫茎泽兰(Eupatorium adenophorum)是20世纪40年代经中缅边境传入我国的一种外来入侵种,原产于中美洲的墨西哥和哥斯达黎加,现已在我国南方和西南地区广泛分布,并且其蔓延速度极快,引起了社会各界的广泛关注。近年来,随着研究的不断深入和拓展,新的研究成果不断涌现,使紫茎泽兰成功入侵的机理性问题不断被揭示出来。该文简要介绍了目前我国关于紫茎泽兰研究的几个热点问题,这些问题主要围绕着紫茎泽兰的分布和预测、入侵扩散机制以及防除方法3 个方面展开。其中以紫茎泽兰作为典型外来入侵种来研究其入侵扩散机制的工作最多。该文就目前的研究进展做一综述,并提出今后的研究建议。
The impacts of climate change on the potential distribution of Haloxylon ammodendron
气候变化对梭梭植物适宜分布的影响
Biotic and abiotic variables show little redundancy in explaining tree species distributions
DOI:10.1111/j.1600-0587.2010.06229.x URL [本文引用: 1]
Evolution in invasive plants: Implications for biological control
DOI:10.1016/j.tree.2004.05.010 URL [本文引用: 1]
Predicting the impacts of climate change on the distribution of species: Are bioclimate envelope models useful?
DOI:10.1046/j.1466-822X.2003.00042.x URL [本文引用: 1]
Climatic niche shifts are rare among terrestrial plant invaders
DOI:10.1126/science.1215933
PMID:22422981
[本文引用: 1]
The assumption that climatic niche requirements of invasive species are conserved between their native and invaded ranges is key to predicting the risk of invasion. However, this assumption has been challenged recently by evidence of niche shifts in some species. Here, we report the first large-scale test of niche conservatism for 50 terrestrial plant invaders between Eurasia, North America, and Australia. We show that when analog climates are compared between regions, fewer than 15% of species have more than 10% of their invaded distribution outside their native climatic niche. These findings reveal that substantial niche shifts are rare in terrestrial plant invaders, providing support for an appropriate use of ecological niche models for the prediction of both biological invasions and responses to climate change.
Opening the black box: An open-source release of Maxent
DOI:10.1111/ecog.03049 URL [本文引用: 2]
The history and status of the study on crofton weed (Eupatorium adenophorum Spreng.)—A worst worldwide weed
世界性恶性杂草——紫茎泽兰研究的历史及现状
Niche dynamics in amphitropical desert disjunct plants: Seeking for ecological and species-specific influences
DOI:10.1111/geb.13215 URL [本文引用: 1]
R: A language and environment for statistical computing
Study of surface dry and wet conditions in Southwest China in recent 50 years
近50a中国西南地区地表干湿状况研究
Biological control opportunities of ragweed are predicted to decrease with climate change in East Asia
DOI:10.17520/biods.2017096
[本文引用: 1]
The control of invasive alien plants (IAP) that jeopardize our ecosystems and economy constitutes a significant challenge for natural resource management. Classical biological control referring to the introduction of specialist antagonists from the native range has proven to be a highly cost-effective management tool against IAP. A critical issue in biological control research is to guide informed decision-making on the potential spread and distribution and thus impact of biological control candidates, especially under climate change. Here we propose a biogeographic modeling approach to predict the cover of the suitable area of a plant invader in East Asia (EA) by two biological control agents and their combinations. Our study system is Ambrosia artemisiifolia, native to North America and invasive worldwide, and two North American biological control agents, Ophraella communa and Epiblema strenuana that were accidentally and deliberately introduced into East Asia (EA) in the late 20th century, respectively. Specifically, we ask: (1) what percentage of the suitable A. artemisiifolia area is also suitable for the two agents in EA, and (2) which part of the suitable A. artemisiifolia area in EA is likely to remain uncovered by these two agents, both under current and future climatic scenarios; and (3) which particular biotypes would be needed to fill in the yet uncovered part of the suitable A. artemisiifolia range in East Asia? For this, we simultaneously modelled the species distributions based on worldwide occurrences and important bioclimatic variables for the target invasive plant and its two biological control agents. Ordination techniques were used to explore climatic constraints of each species and to perform niche overlap and similarity tests with A. artemisiifolia between its native North American and introduced EA range. Our results show that O. communa has a larger overlap with the geographic range of A. artemisiifolia than E. strenuana, both under current (40.3% vs. 21.6% for O. communa and E. strenuana, respectively) and future climatic scenarios (29.8% vs. 20.3% for O. communa and E. strenuana, respectively). Importantly, climate change is expected to reduce the total geographic overlap of A. artemisiifolia by the two agents combined (42.9% vs. 29.8% for current and future climate conditions, respectively), with a higher reduction by O. communa than by E. strenuana. Our analyses also identified for which abiotic conditions to select in order to develop climatically adapted strains for particular regions, where A. artemisiifolia is presently unlikely to be covered.
气候变化预计会减少东亚地区豚草的生物防治效果
DOI:10.17520/biods.2017096
[本文引用: 1]
外来入侵植物对入侵地的生态系统与社会经济可造成严重的威胁。如何控制入侵植物对自然资源的危害, 向人类提出了极大的挑战。利用原产地的专食性天敌来控制入侵植物已被广泛证明是一种经济、可持续的生物防治手段。在全球气候变暖的背景下, 生物防治研究的关键问题是全面了解物种的潜在分布区和传播范围, 进而优化入侵植物的生物防治方案。本研究利用生物地理模型, 通过预测两种生物防治昆虫和它们的组合在东亚地区的适宜分布区, 预测豚草(Ambrosia artemisiifolia)的生物防治区域。豚草原产北美, 目前已经入侵全球多个国家和地区。20世纪末, 豚草条纹叶甲(Ophraella communa)和豚草卷蛾(Epiblema strenuana)作为豚草的生物防治昆虫从原产地引入到东亚地区。本研究旨在探讨如下问题: (1)在豚草的适宜生长分布区内, 有多少区域也同样适宜其两种天敌的生存?(2)在目前和未来的气候背景下, 有多少区域适宜豚草生长但是不适宜它的两种天敌生长?(3)在这些适宜豚草生长却不适宜两种昆虫天敌生长的区域内, 需要选择哪些特定的生物型进行投放?为此, 我们基于入侵植物和两种生物防治昆虫的全球分布记录及其分布点的重要生物气候因子, 同时模拟了入侵植物及其两种生物防治天敌在东亚地区的分布范围。排序技术被用来探索气候因子对每个物种的限制作用, 同时也用来检验豚草在北美和东亚地区的生态位重叠和相似性。结果表明, 在当前和未来的气候背景下, 相较于豚草卷蛾, 豚草条纹叶甲与豚草的地理分布范围更加吻合(当前气候: 40.3% vs. 21.6%, 未来气候: 29.8% vs. 20.3%)。气候变化可能会导致两种生物防治天敌(尤其是豚草条纹叶甲)的地理分布与豚草的地理分布的重叠区域减少(42.9% vs. 29.9%)。本研究同时提出了温度和降水等气候因子可用于为特殊区域(生物防治天敌未覆盖的豚草分布区)筛选生物防治天敌的相应株系。
Predicting global change impacts on plant species’ distributions: Future challenges
DOI:10.1016/j.ppees.2007.09.004 URL [本文引用: 1]
Invasion status and countermeasures of malignant invasive plant (Ageratina adenophora) in Tibet
恶性入侵植物紫茎泽兰在西藏的入侵现状及应对建议
Research on responses of Eupatorium adenophorum’s potential distribution to climate change
紫茎泽兰潜在分布对气候变化响应的研究
Historical Reconstruction of Invasion and Expansion and Potential Spread of Some Threatening Invasive Alien Species in China
我国严重威胁性外来入侵植物入侵与扩散历史过程重建及其潜在分布区的预测
Invasion dynamics and potential spread of the invasive alien plant species Ageratina adenophora (Asteraceae) in China
DOI:10.1111/j.1366-9516.2006.00250.x URL [本文引用: 4]
Application of ecologic niche models in explanation of niche shift of invasive alien species
应用生态位模型研究外来入侵物种生态位漂移
The Beijing Climate Center Climate System Model (BCC-CSM): The main progress from CMIP 5 to CMIP6
DOI:10.5194/gmd-12-1573-2019 URL [本文引用: 1]
Projection and evaluation of extreme temperature and precipitation in major regions of China by CMIP6 models
基于CMIP6的中国主要地区极端气温/降水模拟能力评估及未来情景预估
Road network features and its relationship to the dispersal and spread of Ageratina adenophora in Yunnan, China
云南路网特征及其与紫茎泽兰传播扩散的相关性分析
The badly damaging invasive plant—Eupatorium adenophorum Spreng
危害严重的外来入侵植物——紫茎泽兰
Visualizing patterns of genetic landscapes and species distribution of Taxus wallichiana (Taxaceae), based on GIS and ecological niche models
DOI:10.5814/j.issn.1674-764X.2014.03.001 URL [本文引用: 1]
The suitable distribution of alpine plant Sorbus tianschanica and its environmental driving factors
高山植物天山花楸的适宜分布及其环境驱动因子研究
Predicting potential distribution of Tibetan spruce (Picea smithiana) in Qomolangma (Mount Everest) National Nature Preserve using maximum entropy niche-based model
DOI:10.1007/s11769-011-0483-z URL [本文引用: 1]
Assessing the ecological niche and invasion potential of the Asian giant hornet
Improving ecological niche model transferability to predict the potential distribution of invasive exotic species
DOI:10.3724/SP.J.1003.2014.08178 URL [本文引用: 1]
提高生态位模型转移能力来模拟入侵物种的潜在分布
DOI:10.3724/SP.J.1003.2014.08178
[本文引用: 1]
生态位模型利用物种分布点所关联的环境变量去推算物种的生态需求, 模拟物种的分布。在模拟入侵物种分布时, 经典生态位模型包括模型构建于物种本土分布地, 然后将其转移并投射至另一地理区域, 来模拟入侵物种的潜在分布。然而在模型运用时, 出现了模型的转移能力较低、模拟的结果与物种的实际分布不相符的情况, 由此得出了生态位漂移等不恰当的结论。提高生态位模型的转移能力, 可以准确地模拟入侵物种的潜在分布, 为入侵种的风险评估提供参考。作者以入侵种茶翅蝽(Halyomorpha halys)和互花米草(Spartina alterniflora)为例, 从模型的构建材料(即物种分布点和环境变量)入手, 全面阐述提高模型转移能力的策略。在构建模型之前, 需要充分了解入侵物种的生物学特性、种群平衡状态、本土地理分布范围及物种的生物历史地理等方面的知识。在模型构建环节上, 物种分布点不仅要充分覆盖物种的地理分布和生态空间的范围, 同时要降低物种采样点偏差; 环境变量的选择要充分考虑其对物种分布的限制作用、各环境变量之间的空间相关性, 以及不同地理种群间生态空间是否一致, 同时要降低环境变量的空间维度; 模型构建区域要真实地反映物种的地理分布范围, 并考虑种群的平衡状态。作者认为, 在生态位保守的前提下, 如果模型是构建在一个合理方案的基础上, 生态位模型的转移能力是可以保证的, 在以模型转移能力较低的现象来阐述生态位分化时需要引起注意。
Effect of the Maxent model’s complexity on the prediction of species potential distributions
DOI:10.17520/biods.2016265 URL [本文引用: 1]
MaxEnt模型复杂度对物种潜在分布区预测的影响
DOI:10.17520/biods.2016265
[本文引用: 1]
生态位模型在入侵生物学和保护生物学中具有广泛的应用, 其中Maxent模型最为流行, 被越来越多地应用在预测物种的现实分布和潜在分布的研究中。在Maxent模型中, 多数研究者采用默认参数来构建模型, 这些默认参数源自早期对266个物种的测试, 以预测物种的现实分布为目的。近期研究发现, Maxent模型采用复杂机械学习算法, 对采样偏差敏感, 易产生过度拟合, 模型转移能力仅在低阈值情况下较好。基于默认参数的Maxent模型不仅预测结果不可靠, 而且有时很难解释。在本研究中, 作者以入侵害虫茶翅蝽(Halyomorpha halys)为例, 采用经典模型构建方案(即构建本土模型然后将其转移至入侵地来评估), 利用ENMeval数据包来调整本土Maxent模型调控倍频和特征组合参数, 分析各种参数条件下模型的复杂度, 然后选取最低复杂度的模型参数(即为最优模型), 综合比较默认参数和调整参数后Maxent模型的响应曲线和预测结果, 探讨Maxent模型复杂度对预测结果的影响及Maxent模型构建时所需注意事项, 以期对物种潜在分布进行合理的预测, 促进Maxent模型在我国的合理运用和发展。作者认为, 环境变量的选择至关重要, 需要综合分析其对所模拟物种分布的限制作用和环境变量之间的空间相关性。构建Maxent模型前需对物种分布采样偏差及模型的构建区域进行合理地判断, 模型构建时需要比较不同参数下模型的预测结果和响应曲线, 选取复杂度较低的模型参数来最终建模。在茶翅蝽的分析中, Maxent模型的默认参数和最优模型参数不同, 与Maxent模型默认参数相比, 采用调整参数后所构建的模型预测效果较好, 响应曲线较为平滑, 模型转移能力较高, 能够较为合理反映物种对环境因子的响应和准确地模拟该物种的潜在分布。
On the niche stasis of intercontinental invasive plants
DOI:10.3724/SP.J.1003.2010.547 URL [本文引用: 6]
洲际入侵植物生态位稳定性研究进展
DOI:10.3724/SP.J.2010.547
[本文引用: 6]
人类活动引起的大规模洲际物种交换与生物入侵, 改变了当地生态系统结构与功能, 使生物多样性受到日益严重的威胁。本文通过综合分析主要国家和地区入侵植物的地理起源, 发现洲际入侵主要包括东亚—北美、东亚—南美、欧洲—南非、欧洲—北美、欧洲—东亚、北美—大洋洲等, 这些洲际入侵造成的后果往往比陆内入侵更为严重。利用物种分布模型(SDMs)预测入侵物种潜在分布范围是有效管理和提早预防生物入侵的重要依据, 但这些模型的一个关键假定是: 入侵物种的生态位在空间和时间上是保守的、稳定的。然而, 对于远离原产地种群并能快速适应新生境的洲际入侵植物来说, 生态位可能发生显著的变化。入侵种能否在入侵地保持原有的生态位, 取决于制约其生态分布的限制因素和生态过程在不同地区间是否发生变化。本文中作者总结了洲际入侵与陆内入侵的生态与进化过程的异同点, 认为这些限制物种原产地分布的因素如扩散限制、种间互作、适应性进化、生态可塑性和种群遗传特性等均可能导致入侵物种生态位的改变。建议下一步的研究应该重视: (1)对生态位属性进行多尺度的研究, 包括时间、空间、环境或系统发育等几个方面; (2)对比生态位稳定与发生偏移的物种特性, 确定什么样的入侵物种更容易改变原有的生态位; (3)进行生态位时间动态格局研究, 探讨生态位变化的倾向、历史速率和偏移程度, 以便判定生态位变化趋势。这些研究结果将会进一步提高物种分布模型的预测能力, 有助于更为准确地揭示气候变化和物种入侵对生物多样性的影响。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}