


生物多样性和生产力是生态系统的核心指标, 是实现生态系统服务与功能的根本所在(Tilman et al, 2014)。在全球气候变化的大环境下, 生物多样性锐减, 生态系统保护面临空前困境(Chapin et al, 2000)。因此, 保护生物多样性, 提高和恢复生态系统功能具有重要意义(Wilsey & Potvin, 2000)。在生物多样性和生态系统功能的关系中, 多样性和生产力之间的关系及其维持机制是全球科学家所关注的核心问题之一(Loreau et al, 2001; 贺金生等, 2003)。大量研究表明物种多样性与生产力呈正相关(Wilson et al, 1990; Tilman & Downing, 1994; Ouyang et al, 2016; 吴初平等, 2018), 但负相关(Liang et al, 2007)、驼峰关系(Loreau et al, 2001)和不相关(Grace et al, 2016)也有报道。森林生态系统中生物量和生产力之间既有联系又有区别, 有研究表明, 在森林生态系统中生产力高的物种其碳储存能力通常要低于生产力低的物种(吴初平等, 2018), 且生物量和生产力对物种丰富度的响应也有差异(Cardinale et al, 2006)。综上, 关于天然林中物种多样性对生产力和生物量的影响及其维持机制仍存在较大争议(Zhang & Chen, 2015; Fotis et al, 2017)。为了加深对复杂自然生态系统中多样性与生产力关系和机制的理解, 亟需进一步加强多样性与生产力和生物量关系的研究。
同时, 群落结构多样性对森林生产力的影响也非常重要(Liang et al, 2007; Lei et al, 2009; Long & Shaw, 2010 )。结构多样性通常被定义为空间分布、物种多样性和树木尺寸变化(如树木高度和直径)的一种或多种组合(Lei et al, 2009)。复杂的结构可以提高树木对光的捕获和利用效率, 并提高树木利用水和土壤养分的效率, 还可以促进森林生态系统中生物量的积累(Hardiman et al, 2011; Wang et al, 2011)。结构多样性除了能表征群落的空间分布, 也可以反映群落总体的生物多样性, 不同物种间的固有差异以及同种或异种个体间的不对称竞争有助于树木在空间中形成多元配置, 提高资源利用效率和生产力(Yachi & Loreau, 2007; Lei et al, 2009)。研究表明, 个体之间的差异、立地密度、林龄、最大径级和大树密度等林分结构属性都会改变森林生物量和生产力格局的多个方面(Paoli et al, 2008; Slik et al, 2013; Zhang & Chen, 2015; Yuan et al, 2018)。物种多样性和林分结构属性的变化也直接关系到林分的生长和管理实践(Kant, 2002; Liang et al, 2005; Lei et al, 2009)。综上所述, 群落结构多样性对生产力和生物量的影响机制亟待进一步阐明。
植物群落的分布格局是不同尺度上气候、土壤、地形等环境因子综合作用的结果(Ricklefs et al, 1999)。植被的生产力也受到物种组成和地理环境等因素的影响(Doherty et al, 2011; Zhang et al, 2012)。土壤肥力和地形的差异都是影响植物群落结构和生产力的关键因素(Perroni-Ventura et al, 2006; Long et al, 2012)。研究表明高的土壤养分利用率可以促进植物快速生长, 进而对树木的生产力具有积极效应(Quesada et al, 2012; Prado-Junior et al, 2016)。相比于土壤贫瘠的森林, 土壤肥沃的森林中生物多样性对生产力的影响更大(Paquette & Messier, 2011)。此外, 海拔、坡度和坡向等地形特征造成的环境差异也会影响树种的分布、生物量的积累和生产力的变化( Valencia et al, 2009; Murphy et al, 2015)。由于环境条件、时空尺度和群落类型等因素的差异, 物种多样性和生产力之间的关系尚未得到一致的结论(Willig, 2011)。因此, 复杂的天然林生态系统中生物多样性与生产力的关联及其与环境因素的关系有待进一步阐明(谭凌照等, 2017)。
鼎湖山地处中国南亚热带典型季风气候区, 其地带性植被常绿阔叶林具有400多年的保护历史(方运霆等, 2003; 张亚茹等, 2014)。近年来该区域受气候变化等多重因素的影响出现了氮沉降增加和磷限制加剧等问题, 亟需加强对该生态系统生物多样性与生物量和生产力关系的研究(方运霆等, 2005; 黄文娟等, 2009; Lie et al, 2019)。分析南亚热带常绿阔叶林树木多样性与生产力和生物量的关联及其影响因素, 对于南亚热带常绿阔叶林生产力的提高和生物多样性的保护尤为重要。本研究以鼎湖山南亚热带常绿阔叶林群落为研究对象, 结合多种统计方法分析和评估树木多样性与生物量和生产力之间的关联及其影响因素。主要探讨以下问题: (1)生产力是否受物种多样性和林分结构多样性的影响? (2)生物量是否受物种多样性和林分结构多样性的影响? 是否与物种多样性和林分结构多样性对生产力的影响一致? (3)环境因子如何影响物种多样性和林分结构多样性?
1 材料与方法
1.1 研究地概况
鼎湖山国家级自然保护区位于广东省肇庆市鼎湖区(23.16°-23.19° N, 112.51°-112.56° E), 总面积1,155 ha, 主要地形为丘陵和低山, 最高峰鸡笼山海拔1,000.3 m, 属中国南亚热带季风湿润型气候(熊鑫等, 2016)。年平均气温22.5℃, 最冷月(1月)和最热月(7月)平均气温分别为13.8℃和28.8℃。降水量1,714 mm, 4-9月为湿季, 10月至翌年3月为旱季, 干湿季分明。年平均蒸发量1,115 mm, 年平均相对湿度82%, 灾害性天气为寒潮和台风。土壤母质主要为泥盆纪砂岩、砂页岩、页岩和石英砂岩, 主要土壤类型为赤红壤和红壤。该地区主要的植被是南亚热带常绿阔叶林, 乔木层优势种主要为木荷(Schima superba)、锥(Castanopsis chinensis)、鼎湖血桐(Macaranga sampsonii)、香楠(Aidia canthioides)等。灌木层优势种主要为柏拉木(Blastus cochinchinensis)、九节(Psychotria rubra)、罗伞树(Ardisia quinquegona)等。草本层优势种主要是淡竹叶(Lophatherum gracile)、芒萁(Dicranopteris dichotoma)、沙皮蕨(Hemigramma decurrins)等。
1.2 样地设置与群落调查
本研究选取鼎湖山南亚热带常绿阔叶林为研究对象, 该森林样地位于低山中坡, 具有400多年的保护历史, 处于演替顶极群落阶段, 郁闭度约95%。样地面积为1 ha (100 m × 100 m), 群落调查时先将样地划分为100个10 m × 10 m的II级样方, 每个II级样方再划分为4个5 m × 5 m的III级样方(共400个)。对样地内所有胸径(DBH) ≥ 1 cm的木本植物进行调查, 记录其种名、胸径、树高、生长状态和空间坐标, 每个植株给定一个唯一的编号以便后续进行追踪测定。2010年和2015年我们在样地内共调查了8,084棵木本植物, 隶属37科60属。
1.3 土壤取样和地形指标测定
在每个III级样方内0-30 cm深的土壤中利用土钻采集5个样品(对角线上), 去掉上层浮土, 混合均匀。取一部分迅速装于密封袋内, 剩余部分装在土壤袋中带回实验室测定土壤含水量(WC)、有机碳(C)、pH值(pH)、全氮(N)、铵态氮(NH4+)、速效磷(AP)和速效钾(AK) 7个指标(刘光菘等, 1996)。用标尺、重锤和激光笔配合测定各III级样方边缘的相对高差, 以整个样地海拔最低的一边为基线, 推算每个样地的相对海拔。以样地最低点作为零点, 推算出每个III级样方的顶角在样地中的相对海拔。每个III级样方的相对海拔值由四个顶角坐标的平均值表示。每个样地的地形变量包括相对海拔(RA)、东西坡度(SEW)、南北坡度(SNS)、东西相对高差(RDEW)和南北相对高差(RDNS) (表1)。
表1 南亚热带常绿阔叶林样地基础变量信息统计表
Table 1
| 变量 Variables | 范围 Range | 平均值 Mean | 标准偏差 SD |
|---|---|---|---|
| 物种丰富度 Species richness (R) | 6.00-21.50 | 12.86 | 3.69 |
| Shannon-Wiener多样性指数 Shannon-Wiener diversity index (H') | 1.23-2.68 | 2.07 | 0.31 |
| Simpson多样性指数 Simpson diversity index (D) | 0.62-0.91 | 0.82 | 0.06 |
| Pielou均匀度指数 Pielou evenness index (E) | 0.59-0.98 | 0.82 | 0.08 |
| 胸高断面积标准偏差 Basal area standard deviation (SDba) | 14.83-4,696.75 | 333.41 | 678.16 |
| 胸高断面积变异系数 Basal area coefficient of variation (CVba) | 86.49-2,069.74 | 486.98 | 371.29 |
| 胸高断面积基尼系数 Basal area Gini coefficient (GCba) | 0.55-1.00 | 0.82 | 0.10 |
| 树高标准偏差 Height standard deviation (SDh) | 1.49-8.40 | 4.14 | 1.77 |
| 树高变异系数 Height coefficient of variation (CVh) | 28.73-147.37 | 79.32 | 30.73 |
| 树高基尼系数 Height Gini coefficient (GCh) | 0.18-0.54 | 0.35 | 0.08 |
| 坡度(南北) Slop degree (Northsouth) (SNS) (°) | 0.00-89.61 | 10.52 | 11.86 |
| 坡度(东西) Slop degree(Eastwest) (SEW) (°) | 4.72-39.00 | 26.79 | 5.78 |
| 相对高差(南北) Relative dispersion (Northsouth) (RDNS) (m) | 0.00-3.07 | 0.95 | 0.83 |
| 相对高差(东西) Relative dispersion (Eastwest) (RDEW) (m) | 0.39-4.09 | 2.58 | 0.62 |
| 相对海拔 Relative altitude (RA) (m) | 4.50-66.20 | 33.42 | 15.47 |
| 土壤含水量 Water content (WC) | 0.20-0.30 | 0.24 | 0.02 |
| pH值 pH value | 3.74-4.05 | 3.85 | 0.06 |
| 有机碳 Organic carbon (C) (mg/g) | 18.79-42.75 | 26.18 | 4.37 |
| 全氮 Total nitrogen (TN) (mg/g) | 1.61-3.62 | 2.14 | 0.33 |
| 铵态氮 Ammonium nitrogen (NH4+) (mg/g) | 0.00-0.01 | 0.01 | 0.00 |
| 速效磷 Available phosphorus (AP) (mg/g) | 0.00-0.01 | 0.00 | 0.00 |
| 速效钾 Available potassium (AK) (mg/g) | 0.08-0.23 | 0.12 | 0.03 |
1.4 多样性测度
1.4.1 物种多样性
式中, S为样方中物种数量, 选取2015年和2010年同一样方的平均物种数; Pi为群落中第i个物种的重要值。利用重要值(importance value, IV)来反映每个物种在群落中的相对重要性, 计算公式为: IV = (相对多度 + 相对优势度)/2。
1.4.2 结构多样性
式中, baj是样方内按大小升序第j棵树的胸高断面积, hj是样方内按大小升序第j棵树的树高(m),
1.5 生物量和生产力的计算
式中, DBH为胸径(cm), a和b是回归系数。考虑到地形和土壤数据是两年份之间的调查数据, 本研究的生物量采用2015年和2010年的平均生物量。考虑到样方内起始的生物量可能会反过来影响样方内生物量的变化值, 本研究用生物量的相对变化量表示群落的初级生产力(文中统称为“生产力”)。为了保证数据的正态性, 对生物量和生产力的数据进行对数转换。Biomass2015为2015年调查时样方内所有个体生物量之和, Biomass2010为2010年调查时样方内所有个体生物量之和, t为从2010年到2015年的时间间隔。
1.6 数据统计和分析
首先, 分别对物种多样性、结构多样性、土壤养分和地形因子进行主成分分析(principal component analysis, PCA), 再运用一元回归模型检验树木多样性与生物量和生产力之间的关系。其次, 利用Pearson相关性分析解释物种多样性、结构多样性、生产力、生物量、土壤养分和地形因子变化之间的相关关系, 统计显著性水平设为P < 0.05。最后, 运用方差分解(variation partitioning analysis, VPA)解释每组因子的单独效应和共同效应, 利用韦恩图表示各因子对生物量和生产力变化的相对贡献。同时, 利用结构方程模型(structural equation modeling, SEM)检验土壤养分和地形因子通过物种多样性和结构多样性对生产力和生物量的直接和间接效应。采用最大似然法对结构方程模型进行拟合, 利用比较拟合指数(CFI)、渐进残差均方和平方根(RMSEA)、显著性概率值(P)评价模型优度, 拟合优度的临界值为CFI > 0.9, RMSEA < 0.08, P > 0.05, AIC越小越好(温纯和金光泽, 2019)。数据整理与计算使用Excel 2016, 结构方程模型构建采用AMOS 21.0, 统计分析及作图在R 3.3.3软件中完成。
2 结果
2.1 树木多样性与生产力和生物量之间的关系
利用主成分分析对物种多样性指标和结构多样性指标进行聚类分析(表2)。结果表明物种多样性第一主轴的解释率为73.4%, 第二主轴的解释率为25.5% (图1a)。结构多样性第一主轴的解释率为70.4%, 第二主轴的解释率为15.5% (图1b)。其第一主轴载荷量均大于0.7, 说明第一个主轴能代表物种多样性和结构多样性的变化。树木多样性与生物量和生产力的一元线性回归结果显示, 在南亚热带常绿阔叶林中生产力随着物种多样性的增加而极显著增加(R2 = 0.12, P < 0.01, 图1c), 而生物量随着物种多样性的增加极显著减少(R2 = 0.10, P < 0.01, 图1c)。相反, 生产力随着结构多样性的增加而减少(R2 = 0.20, P < 0.001, 图1d), 生物量随着结构多样性的增加而极显著增加(R2 = 0.52, P < 0.01, 图1d)。
表2 物种多样性、结构多样性、地形和土壤养分主成分的特征值和贡献率
Table 2
| 类型 Class | 成分 Component | 特征值 Eigenvalue | 方差贡献率 Contribution rate of variance (%) | 方差累积贡献率 Cumulative contribution rate of variance (%) |
|---|---|---|---|---|
| 物种多样性 Species diversity | 1 | 2.93 | 73.45 | 73.45 |
| 2 | 1.02 | 25.52 | 98.97 | |
| 3 | 0.03 | 0.76 | 99.73 | |
| 4 | 0.01 | 0.27 | 100 | |
| 结构多样性 Structural diversity | 1 | 4.22 | 70.37 | 70.37 |
| 2 | 0.93 | 15.54 | 85.91 | |
| 3 | 0.47 | 7.78 | 93.69 | |
| 4 | 0.25 | 4.19 | 97.88 | |
| 5 | 0.07 | 1.11 | 98.99 | |
| 6 | 0.06 | 1.01 | 100 | |
| 地形 Topography | 1 | 2.77 | 55.35 | 55.35 |
| 2 | 1.49 | 29.77 | 85.12 | |
| 3 | 0.61 | 12.13 | 97.25 | |
| 4 | 0.13 | 2.66 | 99.91 | |
| 5 | 0.00 | 0.09 | 100 | |
| 土壤养分 Soil nutrients | 1 | 3.51 | 50.17 | 50.17 |
| 2 | 1.37 | 19.62 | 69.79 | |
| 3 | 0.84 | 11.98 | 81.77 | |
| 4 | 0.65 | 9.22 | 90.99 | |
| 5 | 0.42 | 6.05 | 97.04 | |
| 6 | 0.13 | 1.89 | 98.93 | |
| 7 | 0.07 | 1.07 | 100 |
图1
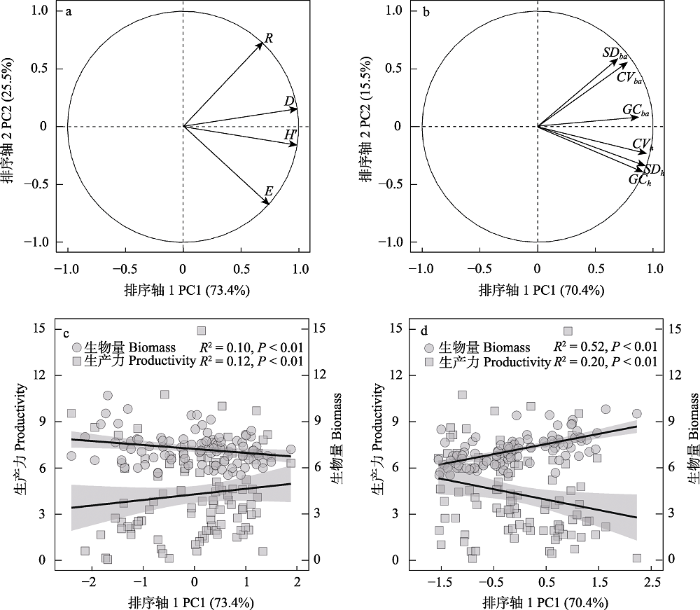
图1
物种多样性和结构多样性与生物量和生产力之间的相关关系。(a) 4个物种多样性指标的第一轴和第二轴载荷量; (b) 6个结构多样性指标的第一轴和第二轴载荷量; (c)物种多样性第一主轴载荷量与生物量和生产力之间的关系; (d)结构多样性第一主轴载荷量与生物量和生产力之间的关系。缩写代表含义同
Fig. 1
Relationship between species diversity and structural diversity with plant biomass and productivity. (a) Loadings on the first and second axes of four species diversity indexes; (b) Loadings on the first and second axes of six structural diversity indexes; (c) Relationship between loadings on the first axe of species diversity and plant biomass and productivity; (d) Relationship between loadings on the first axe of structural diversity and plant biomass and productivity. Abbreviations have the same as in
2.2 环境因子与树木多样性、生产力和生物量的相关性
Person相关性分析结果显示, 生产力与土壤含水量显著负相关(P < 0.05), 而生物量与地形和土壤养分均没有显著的相关关系(图2)。物种丰富度与有机碳、速效磷和速效钾呈显著负相关(P < 0.05), 与坡度(南北)和相对高差(南北)极显著负相关(P < 0.01), 与全氮呈负相关(P<0.001)。Shannon-Wiener多样性指数与坡度(南北)显著负相关(P < 0.05), 与全氮极显著负相关(P < 0.01), 与铵态氮极显著正相关(P < 0.01)。Simpson多样性指数与相对高差(南北)显著负相关(P < 0.05), 与铵态氮显著正相关, 与坡度(南北)和全氮极显著负相关(P < 0.01)。Pielou均匀度指数与土壤含水量和铵态氮显著正相关(P < 0.05)。胸高断面积变异系数与土壤含水量和铵态氮显著负相关(P < 0.05)。胸高断面积基尼系数与速效磷呈显著负相关(P < 0.05)。树高标准偏差与相对海拔、土壤含水量和速效磷呈显著负相关(P < 0.05); 树高变异系数与有机碳、铵态氮、速效磷和速效钾呈显著负相关(P < 0.05), 与土壤含水量极显著负相关(P < 0.01)。树高基尼系数与相对海拔、土壤含水量、有机碳和速效磷呈显著负相关(P < 0.05) (图2)。
图2
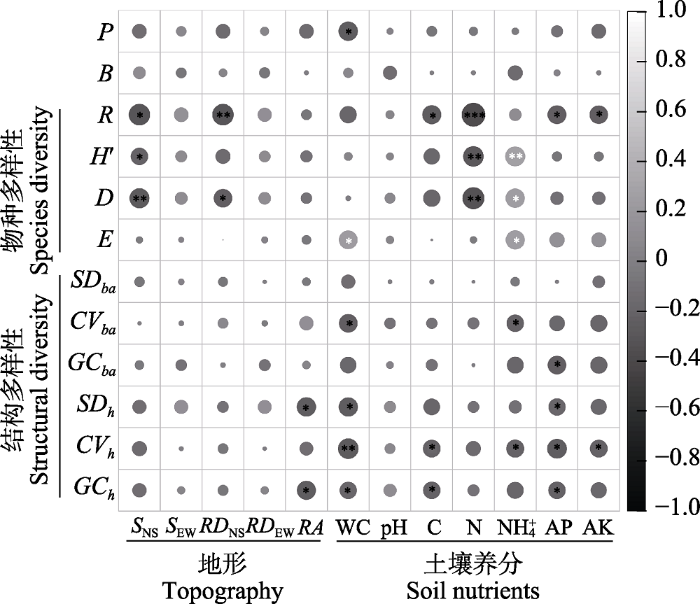
图2
南亚热带常绿阔叶林树木多样性、生物量和生产力与土壤养分和地形因子之间的Pearson相关关系。P: Productivity; B: Biomass; 其余缩写代表含义同
Fig. 2
Pearson correlation between tree species diversity, plant biomass, plant productivity and soil nutrients, topographic factors in subtropical evergreen broad‐leaved forest. The letter B and P mean biomass and productivity, and other abbreviations have the same as in
2.3 环境因子对生产力和生物量的直接和间接影响
方差分解分析结果表明, 结构多样性对生产力的单独效应的解释率最大, 为5.21%; 结构多样性和物种多样性共同效应的解释率次之, 为3.53%; 地形与结构多样性共同效应和土壤养分与结构多样性共同效应的解释率较小, 分别为1.09%和0.19% (图3a)。结构多样性对生物量单独效应的解释率最大, 为35.39%; 结构多样性和物种多样性共同效应的解释率次之, 为13.66%; 地形与结构多样性共同效应和土壤养分与结构多样性共同效应的解释率较小, 分别为3.52%和3.49% (图3b)。进一步利用结构方程模型进行分析, 结果表明各变量分别解释生产力和生物量的变化的34%和52% (图3c)。物种多样性对生产力有显著的直接影响(P < 0.05), 反映作用大小的路径系数为0.18。结构多样性对生物量具有直接影响(P < 0.001, 路径系数为0.72)。各变量分别解释了物种多样性和结构多样性变化的29%和5%。地形和土壤养分对物种多样性具有显著的直接影响, 路径系数分别为‒0.23和‒0.26; 土壤养分对结构多样性具有显著的直接影响, 路径系数为‒0.22。结构多样性对物种多样性具有极显著影响, 路径系数为‒0.48; 生物量对生产力具有极显著影响, 路径系数为‒0.50 (图3c)。
图3
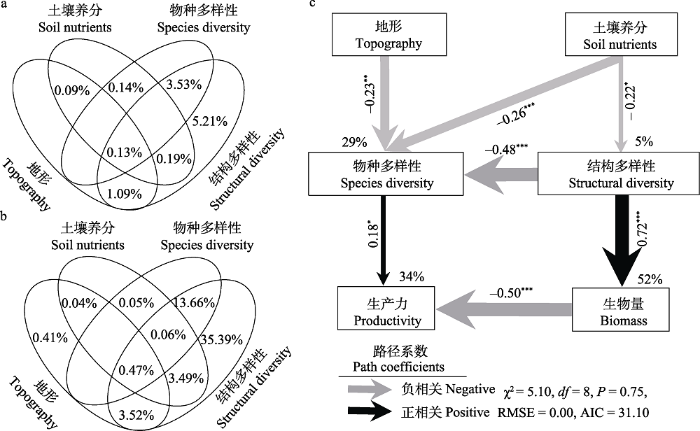
图3
物种多样性、结构多样性、地形和土壤养分对生产力和生物量的影响。(a)方差分解解释各因子对生产力的单独效应和共同效应; (b)方差分解解释各因子对生物量的单独效应和共同效应; (c)结构方程模型(SEM)解释地形和土壤养分通过物种多样性和结构多样性对生产力的直接和间接影响(N = 100)。SEM考虑了所有可能的路径, R2表示所解释的方差比例。箭头上的数字表示标准化的路径系数。箭头宽度表示路径系数的强度。*, P < 0.05, **, P < 0.01, ***, P < 0.001。
Fig. 3
The impact of species diversity, structural diversity, topography and soil nutrients on plant productivity and biomass. (a) Variation partitioning analysis explains the pure and shared effect of factors on plant productivity. (b) Variation partitioning analysis explains the pure and shared effect of factors on plant biomass. (c) Structural equation model (SEM) reveals the direct and indirect effects of species diversity, structural diversity, topography and soil nutrients on plant productivity and biomass (N = 100). The SEM considered all plausible pathways, to increase the degrees of freedom, R2 indicates the proportion of variance explained. The numbers on the arrows indicate standardized path coefficients. The arrow width is proportional to the strength of the path coefficients. *, P < 0.05, **, P < 0.01, ***, P < 0.001.
3 讨论
3.1 物种多样性与生产力和生物量的关系
本研究结果表明物种多样性与生产力呈显著正相关关系, 这与早期生产力与物种多样性关系的研究结果相似(黄小荣等, 2018; Liu et al, 2019)。这可能由于鼎湖山南亚热带常绿阔叶林群落郁闭度较高, 林龄超过200年, 生态位分化合理, 使得空间资源能够被充分利用, 进而增加了植被的生产力(Loreau et al, 2001; Chen et al, 2016)。并且以往研究表明物种丰富度高的群落更偏向于增加高产物种进入群落的机会, 而本研究鼎湖山南亚热带常绿阔叶林已经处于群落演替后期, 高产物种已经形成了稳定的种间关系, 因此高物种多样性也是植被生产力增加的原因之一(Vile et al, 2006; Cadotte, 2017)。此外, 鼎湖山南亚热带常绿阔叶林中群落物种多样性与生物量呈负相关, 该结果与在浙江定海次生林生物量与物种多样性的结果并不一致(吴初平等, 2018)。这可能由于本研究群落处于近演替顶极阶段(邹顺等, 2018), 在演替后期南亚热带常绿阔叶林群落结构较为稳定, 群落的总生物量主要取决于群落中的高碳储量的物种, 高碳储量的物种竞争性导致更多的生物量积累(Potter & Woodall 2014; Ouyang et al, 2016)。而鼎湖山常绿阔叶林以往研究显示, 过去30年间群落中大径级个体和不抗旱植物个体数减少, 小径级个体和抗旱或速生植物个体数增加, 使得群落中总生物量呈现下降的趋势, 这可以用来解释本研究中物种多样性与生物量的负相关关系(邹顺等, 2018)。相关研究也表明当生物量超过一定水平时, 物种丰富度与生物量呈显著的负相关(郭志华等, 2002)。此外, 本研究显示鼎湖山南亚热带常绿阔叶林生物量与生产力具有负相关关系, 这与早期在该地区的研究结果一致(Prado-Junior et al, 2016)。这可能是由于环境因素对生物量和生产力关系的影响(Ma et al, 2010), 本研究群落处于演替后期阶段, 林分内养分循环处于稳定甚至关闭的状态, 小径级个体虽有生长, 但大径级个体生物量在群落中占主导地位, 而研究区域内大径级个体受到环境因素的影响个体数减少, 导致群落生物量与生产力之间表现出负相关关系。
3.2 结构多样性与生产力和生物量的关系
早期研究表明, 群落结构变异在一定程度上反映了物种在资源利用上的生态位互补, 是促进生产力的一个重要机制(谭凌照等, 2017)。而鼎湖山南亚热带常绿阔叶林结构多样性与生物量呈显著的正相关关系, 与生产力却呈显著的负相关关系。本研究中结构多样性与生产力的关系与大多数结果不一致(即结构多样性促进生产力) (谭凌照等, 2017; Lei et al, 2009)。这可能是由研究样地所处的演替阶段、环境条件和群落结构差异所导致的。南亚热带常绿阔叶林受到气候条件变化和自然干扰的影响, 群落整体、群落垂直层次内和垂直层次之间的正相关关系逐渐减弱, 物种间的互补性不足, 对资源的利用效率下降, 导致群落结构多样性与生产力呈现负相关关系。这可能是由于鼎湖山常绿阔叶林群落更新率和死亡率上升, 大径级个体占据主要地位, 虽有小径级个体快速更新提升群落生产力, 但仍然不能够弥补大径级个体死亡对群落所造成的影响(邹顺等, 2018)。群落结构多样性与生物量的正向关系可以在一定程度上表明结构多样性是促进个体生物量积累的。一方面群落结构与资源的充分利用具有密切关系, 竞争和资源的充分利用决定了物种分布格局的多样化; 另一方面群落结构与物种组成有密切的关系, 高碳储量的物种占据着群落主要地位(Potter & Woodall 2014; Ouyang et al, 2016)。因此, 南亚热带常绿阔叶林群落结构多样性是促进群落生物量积累的。此外, 本研究显示结构多样性与生产力和生物量的关系与物种多样性与生产力和生物量的关系呈相反的趋势, 这可能是由于本研究中结构多样性对物种多样性具有反向作用, 群落属于近演替顶极, 群落结构较为稳定, 而近年来该群落受环境影响个体不断死亡造成群落巨大波动, 影响了物种组成, 使结构多样性与物种多样性呈现出负相关关系(邹顺等, 2018)。
3.3 环境因子对多样性的影响
地形和土壤养分的变化决定了物种分布、森林碳库组成和功能的多个方面。自然群落中的物种组成是在漫长的演化过程中, 物种与环境之间相互依赖、相互作用, 从而适应当地环境条件所形成的特定群落结构, 是对环境响应的综合反映(Ricklefs et al, 1999)。我们的研究结果表明地形对植被生产力和生物量无显著的直接影响, 但对物种多样性具有显著的负影响, 这与吴初平等(2018)在浙江定海次生林地的研究具有一定的差异。当然, 群落结构特性也受区域特征和群落发展阶段的影响(方精云等, 2004; Jafari et al, 2004)。本研究中地形对物种多样性的影响主要是由于样地的斑块状特征, 坡度呈西南-东北走向, 从南到北坡度逐渐增加, 而物种多样性逐渐下降, 导致地形与物种多样性呈现显著的负相关关系。
土壤和植被的养分状况常作为衡量环境对植被生长养分限制状况的指标。在热带和亚热带地区, 由于土壤高度风化以及土壤中磷的有效性低等特点, 土壤养分常常成为该地区限制植物生长的主要因素(Vitousek & Howarth, 1991)。本研究中南亚热带常绿阔叶林高土壤有机碳和土壤有效磷含量的环境条件下, 群落结构复杂; 而高土壤氮含量环境条件下, 物种多样性较低, 这与早期研究具有相似的结果(冯健等, 2021)。这可能是由于南亚热带常绿阔叶林物种多样性和结构多样性主要受土壤有效磷和氮沉降的影响, 植物生长所需要的土壤有效磷主要来源于土壤, 植被中土壤有效磷含量越多, 结构多样性越复杂。并且南亚热带常绿阔叶林由于氮沉降导致物种多样性逐渐降低(鲁显楷等, 2008)。因此, 南亚热带常绿阔叶林土壤养分对物种多样性和结构多样性有明显的影响。
综上, 本研究以未经人为干扰的中国南亚热带地区的地带性顶极群落为研究对象, 基于森林生态系统乔木层群落调查数据, 结合样地的地形和土壤养分, 研究了南亚热带常绿阔叶林树木多样性和生产力之间的关联及其影响因素。主要结论如下: (1)物种多样性对生产力有最大直接效应, 结构多样性对生物量有最大直接效应; (2)在南亚热带常绿阔叶林中物种多样性在解释生产力方面要优于结构多样性, 而结构多样性在解释生物量积累方面要优于物种多样性; (3)物种多样性和结构多样性均受地形和土壤因子的影响。因此, 在南亚热带常绿阔叶林的经营和管理过程中充分考虑物种多样性和结构多样性对生产力和生物量的影响, 将有助于更好的保持南亚热带常绿阔叶林的结构多样性, 以维持演替顶级群落的生物多样性和快速生长。
参考文献
Functional traits explain ecosystem function through opposing mechanisms
DOI:10.1111/ele.12796
PMID:28639274
[本文引用: 1]

The ability to explain why multispecies assemblages produce greater biomass compared to monocultures, has been a central goal in the quest to understand biodiversity effects on ecosystem function. Species contributions to ecosystem function can be driven by two processes: niche complementarity and a selection effect that is influenced by fitness (competitive) differences, and both can be approximated with measures of species' traits. It has been hypothesised that fitness differences are associated with few, singular traits while complementarity requires multidimensional trait measures. Here, using experimental data from plant assemblages, I show that the selection effect was strongest when trait dissimilarity was low, while complementarity was greatest with high trait dissimilarity. Selection effects were best explained by a single trait, plant height. Complementarity was correlated with dissimilarity across multiple traits, representing above and below ground processes. By identifying the relevant traits linked to ecosystem function, we obtain the ability to predict combinations of species that will maximise ecosystem function.© 2017 John Wiley & Sons Ltd/CNRS.
Effects of biodiversity on the functioning of trophic groups and ecosystems
DOI:10.1038/nature05202 URL [本文引用: 1]
Consequences of changing biodiversity
DOI:10.1038/35012241 URL [本文引用: 1]
Positive effects of neighborhood complementarity on tree growth in a Neotropical forest
DOI:10.1890/15-0625.1 URL [本文引用: 1]
Diversity-function relationships changed in a long-term restoration experiment
DOI:10.1890/10-1534.1 URL [本文引用: 1]
Ecological characteristics of mountains and research issues of mountain ecology
试论山地的生态特征及山地生态学的研究内容
DOI:10.17520/biods.2004003
[本文引用: 1]
山地是一个生态复杂系统,它具有特定的结构和功能,拥有丰富的生物多样性资源、水资源、矿产资源和旅游资源。开展山地生态学研究对阐明山地系统的结构与功能、山地的生态现象与过程,以及合理开发利用和保护山地资源都有极为重要的意义。本文在简要分析山地地形的主要要素对生态因子影响的基础上,对山地的生态效应进行了归纳,探讨了山地生态学应包含的主要研究内容。作者认为,在山区,地形地貌是形成山地结构和功能以及各种生态现象和过程的最根本因素,它通过改变地表的光、热、水、土、肥等生态因子而发生作用。因此,山地生态学应把地形地貌与各种生态现象和过程的相互作用作为其核心的研究内容。作者提出,山地生态学研究主要包括: 山地生态复杂性与生物多样性、山地气候变化、山地生态工程、山区可持续发展综合研究以及山地生态学研究技术与方法论等内容。
Role of forest succession on carbon sequestration of forest ecosystems in lower subtropical China
森林演替在南亚热带森林生态系统碳吸存中的作用
Response of diameter at breast height increment to N additions in forests of Dinghushan Biosphere Reserve
鼎湖山主要森林类型植物胸径生长对氮沉降增加的初期响应
Study on vegetation diversity and soil characteristics of Larix olgensis uneven-aged mixed stand in eastern Liaoning mountain region
辽东山区长白落叶松异龄混交林植被多样性和土壤特征研究
Above-ground biomass is driven by mass‐ratio effects and stand structural attributes in a temperate deciduous forest
DOI:10.1111/1365-2745.12847 URL [本文引用: 1]
Integrative modelling reveals mechanisms linking productivity and plant species richness
DOI:10.1038/nature16524 URL [本文引用: 1]
The formation and maintenance mechanisms of biodiversity and the research techniques for biodiversity
生物多样性的形成、维持机制及其宏观研究方法
The role of canopy structural complexity in wood net primary production of a maturing northern deciduous forest
The even-aged northern hardwood forests of the Upper Great Lakes Region are undergoing an ecological transition during which structural and biotic complexity is increasing. Early-successional aspen (Populus spp.) and birch (Betula papyrifera) are senescing at an accelerating rate and are being replaced by middle-successional species including northern red oak (Quercus rubra), red maple (Acer rubrum), and white pine (Pinus strobus). Canopy structural complexity may increase due to forest age, canopy disturbances, and changing species diversity. More structurally complex canopies may enhance carbon (C) sequestration in old forests. We hypothesize that these biotic and structural alterations will result in increased structural complexity of the maturing canopy with implications for forest C uptake. At the University of Michigan Biological Station (UMBS), we combined a decade of observations of net primary productivity (NPP), leaf area index (LAI), site index, canopy tree-species diversity, and stand age with canopy structure measurements made with portable canopy lidar (PCL) in 30 forested plots. We then evaluated the relative impact of stand characteristics on productivity through succession using data collected over a nine-year period. We found that effects of canopy structural complexity on wood NPP (NPPw) were similar in magnitude to the effects of total leaf area and site quality. Furthermore, our results suggest that the effect of stand age on NPPw is mediated primarily through its effect on canopy structural complexity. Stand-level diversity of canopy-tree species was not significantly related to either canopy structure or NPPw. We conclude that increasing canopy structural complexity provides a mechanism for the potential maintenance of productivity in aging forests.
Biodiversity and ecosystem productivity: Why is there a discrepancy in the relationship between experimental and natural ecosystems?
生物多样性与生态系统生产力: 为什么野外观测和受控实验结果不一致?
DOI:10.17521/cjpe.2003.0120
[本文引用: 1]
人们担心生物多样性的空前丧失会危及到生态系统的服务功能,因此有关生物多样性-生态系统生产力这一古老命题的讨论成为当今生态学的热点议题之一。20世纪90年代以David Tilman和Shahid Naeem为代表的生态学家利用大规模的受控实验,对物种多样性与生态系统功能的诸多方面进行
Inorganic nitrogen and available phosphorus concentrations in the soils of five forests at Dinghushan, China
鼎湖山5种森林土壤的无机氮和有效磷含量
Relationship between plant functional diversity and productivity of Pinus massoniana plantations in Guangxi
DOI:10.17520/biods.2018092 URL [本文引用: 1]
广西马尾松林植物功能多样性与生产力的关系
DOI:10.17520/biods.2018092
[本文引用: 1]
探索植物多样性与生产力的关系可为森林经营与管理提供科学基础。本研究以广西4个地区的马尾松(Pinus massoniana)人工林群落为研究对象, 通过计算物种多样性、功能多样性和功能优势值, 运用相关分析、自动线性建模和方差划分等方法, 分析了多样性与生产力的关系。研究发现, 生产力与物种丰富度、Shannon指数、功能丰富度、功能均匀度极显著正相关(P < 0.01), 与物种均匀度、功能多样性、功能离散度、功能团个数、坡向显著正相关(P < 0.05), 与林龄极显著负相关(P < 0.01), 4个功能多样性参数和4个物种多样性参数两两之间皆为显著正相关; 未发现初始生物量制约生产力的提高; 方差划分最优模型中, 功能多样性参数效应、功能优势值效应和林龄效应分别解释生产力方差的56%、43%和33%, 功能多样性参数效应和功能优势值效应重叠部分高达27%; 生态位互补效应主要由功能丰富度和功能均匀度产生, 选择效应主要由生长型优势值产生; 生长型优势值为灌木的样地生产力较高, 次优种或过渡种对生态系统功能也有重要作用。以生产力为响应变量的自动线性建模最佳子集包括重要性由大到小的5个因素: 林龄、生长型优势值、功能丰富度、功能均匀度、功能团个数。建议维护森林功能多样性, 加强林下叶层植物保护, 用好功能重要的物种, 通过林下叶层的补偿性光合作用和生长竞争, 有效地提高生产力和生物多样性。
Effective environmental factors in the distribution of vegetation types in Poshtkouh rangelands of Yazd Province (Iran)
DOI:10.1016/S0140-1963(03)00077-6 URL [本文引用: 1]
The marginal cost of structural diversity of mixed uneven-aged hard maple forests
DOI:10.1139/x02-001 URL [本文引用: 1]
The relationship between tree biodiversity and biomass dynamics changes with tropical forest succession
DOI:10.1111/ele.2014.17.issue-9 URL [本文引用: 1]
Relationships between stand growth and structural diversity in spruce-dominated forests in New Brunswick, Canada
DOI:10.1139/X09-089 URL [本文引用: 5]
An evaluation of different diameter diversity indices based on criteria related to forest management planning
DOI:10.1016/j.foreco.2005.10.046 URL [本文引用: 1]
Growth and yield of all-aged Douglas-fir-western hemlock forest stands: A matrix model with stand diversity effects
DOI:10.1139/x05-137 URL [本文引用: 1]
Effects of diversity of tree species and size on forest basal area growth, recruitment, and mortality
DOI:10.1016/j.foreco.2007.02.028 URL [本文引用: 2]
Warming changes soil N and P supplies in model tropical forests
DOI:10.1007/s00374-019-01382-7 URL [本文引用: 1]
Linking understory species diversity, community-level traits and productivity in a Chinese boreal forest
DOI:10.1111/jvs.12714
[本文引用: 1]
Question The consensus has been growing over the past decade that functional traits and diversity are better to explain forest overstory diversity-productivity relationships (DPRs) than species diversity. Although the understory accounts for the majority of plant diversity in forests, it remains unclear how understory aboveground biomass production (UABP) is influenced by its species diversity and community-level functional traits. Location Great Xing'an Mountains of ortheastern China. Methods We quantified the effects of species richness, community aggregated traits (community weighted mean trait values, CWM) and functional diversity (functional dispersion, FDis) on UABP using structural equation modeling (SEM), which simultaneously accounted for the effects of overstory tree basal area, stand age, and soil fertility. Results In the full model, species richness had a negative direct, a positive indirect and no total effect on UABP. Furthermore, CWM and FDis, respectively, exhibited positive and no effect on UABP. Among the covariates, soil fertility, stand age, and overstory tree basal area had, respectively, positive, negative, and no effect on UABP. In the model without species richness, all trait variables had similar effects on UABP to those in the full model. In the richness-only model without traits, species richness, soil fertility and stand age had no effect on UABP. Conclusions Our results suggest that the selection effect largely determined understory DPRs due to the stronger effects of CWM on UABP than of FDis. Soil fertility exhibited the strongest influence on understory DPRs due to its parallel influences on traits, diversity, and productivity. The increase in resource availability induced by overstory tree litter-fall likely promoted soil fertility as the main driver of the understory DPRs. Stand age exhibited a negative effect on UABP, which may have contributed to the increases in shrub dominance and decreases in production due to limited resources.
The influence of compositional and structural diversity on forest productivity
DOI:10.1093/forestry/cpp033 URL [本文引用: 1]
Patterns of species diversity and soil nutrients along a chronosequence of vegetation recovery in Hainan Island, South China
DOI:10.1007/s11284-011-0923-3 URL [本文引用: 1]
Biodiversity and ecosystem functioning: Current knowledge and future challenges
The ecological consequences of biodiversity loss have aroused considerable interest and controversy during the past decade. Major advances have been made in describing the relationship between species diversity and ecosystem processes, in identifying functionally important species, and in revealing underlying mechanisms. There is, however, uncertainty as to how results obtained in recent experiments scale up to landscape and regional levels and generalize across ecosystem types and processes. Larger numbers of species are probably needed to reduce temporal variability in ecosystem processes in changing environments. A major future challenge is to determine how biodiversity dynamics, ecosystem processes, and abiotic factors interact.
Effects of nitrogen deposition on forest biodiversity: A review
DOI:10.1016/S1872-2032(09)60012-3 URL [本文引用: 1]
氮沉降对森林生物多样性的影响
Environmental factors covary with plant diversity-productivity relationships among Chinese grassland sites
DOI:10.1111/j.1466-8238.2009.00508.x URL [本文引用: 1]
Species associations structured by environment and land-use history promote beta-diversity in a temperate forest
DOI:10.1890/14-0695.1 URL [本文引用: 1]
Significant effects of biodiversity on forest biomass during the succession of subtropical forest in South China
DOI:10.1016/j.foreco.2016.04.020 URL [本文引用: 3]
Soil nutrients affect spatial patterns of aboveground biomass and emergent tree density in southwestern Borneo
Studies on the relationship between soil fertility and aboveground biomass in lowland tropical forests have yielded conflicting results, reporting positive, negative and no effect of soil nutrients on aboveground biomass. Here, we quantify the impact of soil variation on the stand structure of mature Bornean forest throughout a lowland watershed (8-196 m a.s.l.) with uniform climate and heterogeneous soils. Categorical and bivariate methods were used to quantify the effects of (1) parent material differing in nutrient content (alluvium > sedimentary > granite) and (2) 27 soil parameters on tree density, size distribution, basal area and aboveground biomass. Trees > or =10 cm (diameter at breast height, dbh) were enumerated in 30 (0.16 ha) plots (sample area = 4.8 ha). Six soil samples (0-20 cm) per plot were analyzed for physiochemical properties. Aboveground biomass was estimated using allometric equations. Across all plots, stem density averaged 521 +/- 13 stems ha(-1), basal area 39.6 +/- 1.4 m(2) ha(-1) and aboveground biomass 518 +/- 28 Mg ha(-1) (mean +/- SE). Adjusted forest-wide aboveground biomass to account for apparent overestimation of large tree density (based on 69 0.3-ha transects; sample area = 20.7 ha) was 430 +/- 25 Mg ha(-1). Stand structure did not vary significantly among substrates, but it did show a clear trend toward larger stature on nutrient-rich alluvium, with a higher density and larger maximum size of emergent trees. Across all plots, surface soil phosphorus (P), potassium, magnesium and percentage sand content were significantly related to stem density and/or aboveground biomass (R (Pearson) = 0.368-0.416). In multiple linear regression, extractable P and percentage sand combined explained 31% of the aboveground biomass variance. Regression analyses on size classes showed that the abundance of emergent trees >120 cm dbh was positively related to soil P and exchangeable bases, whereas trees 60-90 cm dbh were negatively related to these factors. Soil fertility thus had a significant effect on both total aboveground biomass and its distribution among size classes.
The effect of biodiversity on tree productivity: From temperate to boreal forests
DOI:10.1111/geb.2011.20.issue-1 URL [本文引用: 1]
Relationship between soil nutrient availability and plant species richness in a tropical semi-arid environment
DOI:10.1111/j.1654-1103.2006.tb02495.x URL [本文引用: 1]
Does biodiversity make a difference? Relationships between species richness, evolutionary diversity, and aboveground live tree biomass across U.S. forests
DOI:10.1016/j.foreco.2013.06.026 URL [本文引用: 2]
Conservative species drive biomass productivity in tropical dry forests
DOI:10.1111/1365-2745.12543 URL [本文引用: 2]
Basin-wide variations in Amazon forest structure and function are mediated by both soils and climate
DOI:10.5194/bg-9-2203-2012 URL [本文引用: 1]
Global patterns of tree species richness in moist forests: Distinguishing ecological influences and historical contingency
DOI:10.2307/3546454 URL [本文引用: 2]
Drivers of productivity and its temporal stability in a tropical tree diversity experiment
DOI:10.1111/gcb.14792
PMID:31486578
[本文引用: 1]
There is increasing evidence that mixed-species forests can provide multiple ecosystem services at a higher level than their monospecific counterparts. However, most studies concerning tree diversity and ecosystem functioning relationships use data from forest inventories (under noncontrolled conditions) or from very young plantation experiments. Here, we investigated temporal dynamics of diversity-productivity relationships and diversity-stability relationships in the oldest tropical tree diversity experiment. Sardinilla was established in Panama in 2001, with 22 plots that form a gradient in native tree species richness of one-, two-, three- and five-species communities. Using annual data describing tree diameters and heights, we calculated basal area increment as the proxy of tree productivity. We combined tree neighbourhood- and community-level analyses and tested the effects of both species diversity and structural diversity on productivity and its temporal stability. General patterns were consistent across both scales indicating that tree-tree interactions in neighbourhoods drive observed diversity effects. From 2006 to 2016, mean overyielding (higher productivity in mixtures than in monocultures) was 25%-30% in two- and three-species mixtures and 50% in five-species stands. Tree neighbourhood diversity enhanced community productivity but the effect of species diversity was stronger and increased over time, whereas the effect of structural diversity declined. Temporal stability of community productivity increased with species diversity via two principle mechanisms: asynchronous responses of species to environmental variability and overyielding. Overyielding in mixtures was highest during a strong El Niño-related drought. Overall, positive diversity-productivity and diversity-stability relationships predominated, with the highest productivity and stability at the highest levels of diversity. These results provide new insights into mixing effects in diverse, tropical plantations and highlight the importance of analyses of temporal dynamics for our understanding of the complex relationships between diversity, productivity and stability. Under climate change, mixed-species forests may provide both high levels and high stability of production.© 2019 The Authors. Global Change Biology published by John Wiley & Sons Ltd.
Large trees drive forest aboveground biomass variation in moist lowland forests across the tropics
DOI:10.1111/geb.2013.22.issue-12 URL [本文引用: 1]
Relationships between species diversity or community structure and productivity of woody-plants in a broad-leaved Korean pine forest in Jiaohe, Jilin, China
DOI:10.17521/cjpe.2016.0321 URL [本文引用: 4]
吉林蛟河阔叶红松林木本植物物种多样性及群落结构与生产力的关系
DOI:10.17521/cjpe.2016.0321
[本文引用: 4]
该研究以吉林蛟河阔叶红松林为分析对象, 比较了物种多样性和群落结构对生产力的影响, 试图为解释多样性-生产力关系、指导森林经营管理提供科学依据。研究利用11.76 hm<sup>2</sup>大样地中10 973株木本植物数据, 通过线性回归模型分析了7个物种多样性和群落结构指标与生产力的相关性, 进而利用结构方程模型比较了物种多样性和群落结构对生产力的影响。结果显示: (1)在线性回归模型中, 物种多样性和群落结构均对生产力有显著作用, 其中物种多样性均匀度指数与生产力显著负相关, 群落结构的香农指数与生产力显著正相关, 基尼系数与生产力显著负相关。(2)在结构方程模型中, 群落结构对生产力的影响比物种多样性更强。研究认为群落结构对生产力的作用比物种多样性更大, 在森林经营管理中, 提高群落结构复杂性对促进生产力具有重要的意义。
Biodiversity and stability in grasslands
DOI:10.1038/367363a0 URL [本文引用: 1]
Biodiversity and ecosystem functioning
DOI:10.1146/ecolsys.2014.45.issue-1 URL [本文引用: 1]
Dissecting biomass dynamics in a large Amazonian forest plot
DOI:10.1017/S0266467409990095 URL [本文引用: 1]
Ecosystem productivity can be predicted from potential relative growth rate and species abundance
DOI:10.1111/j.1461-0248.2006.00958.x URL [本文引用: 1]
Nitrogen limitation on land and in the sea: How can it occur?
Positive relationship between aboveground carbon stocks and structural diversity in spruce-dominated forest stands in New Brunswick, Canada
Effects of functional diversity on productivity in a typical mixed broadleaved-Korean pine forest
DOI:10.17521/cjpe.2018.0312 URL [本文引用: 1]
功能多样性对典型阔叶红松林生产力的影响
DOI:10.17521/cjpe.2018.0312
[本文引用: 1]
为比较生物量比率假说与生态位互补假说在解释生产力变异的相对重要性, 探讨生物多样性和生产力之间的关系是否受到生物和非生物因素的影响, 该研究依托小兴安岭9 hm <sup>2</sup>阔叶红松(Pinus koraiensis)林动态监测样地, 通过计算群落初始生物量、物种多样性、功能多样性、植物性状的群落加权平均值和测定环境因子, 运用线性回归模型、结构方程模型, 比较了物种多样性和功能多样性与生产力的相关性。结果表明: (1)物种多样性和功能多样性均对生产力有显著作用, 功能多样性比物种多样性与生产力的关系更为密切; (2)功能多样性指数比群落加权平均值能更好地解释生产力变异, 说明生态位互补假说更适用于解释阔叶红松林群落内生产力的变异; (3)生物多样性与生产力的关系受生物因素与非生物因素的共同作用, 相较于多样性和功能性状组成(植被质量), 初始林分生物量(植被数量)能更有效地解释生产力的变异。生物多样性与生产力关系的研究应从植被质量与植被数量同时出发, 评估生态系统过程的多种非生物和生物驱动因素, 同时维护森林功能多样性, 加强植物与土壤环境的保护, 对有效增加生产力和维持生物多样性具有重要意义。
Biomass study of the community of Castanopsis chinensis + Cryptocarya concinna + Schina superba in a southern China reserve
鼎湖山锥栗 + 黄果厚壳桂 + 荷木群落生物量及其特征
Evolution and measurement of species diversity
DOI:10.2307/1218190 URL [本文引用: 1]
Biodiversity and productivity
DOI:10.1126/science.1212453 URL [本文引用: 1]
Biodiversity and ecosystem functioning: Importance of species evenness in an old field
DOI:10.1890/0012-9658(2000)081[0887:BAEFIO]2.0.CO;2 URL [本文引用: 1]
Species diversity in relation to ultramafic substrate and to altitude in southwestern New Zealand
DOI:10.1007/BF00045131 URL [本文引用: 1]
Relationships between species richness and biomass/productivity depend on environmental factors in secondary forests of Dinghai, Zhejiang Province
DOI:10.17520/biods.2017320 URL [本文引用: 5]
浙江定海次生林内物种丰富度与生物量和生产力关系的环境依赖性
DOI:10.17520/biods.2017320
[本文引用: 5]
迄今生物多样性与生态系统功能关系的研究主要在物种组成随机配置的人工生态系统中进行, 在自然生态系统中研究较少, 且未考虑环境因子如何影响生态系统功能及其与生物多样性的关系。本研究选取亚热带广泛分布的次生林为研究对象, 利用模型拟合的方法, 探讨亚热带次生林中物种丰富度与生物量和生产力之间的关系, 以及环境因子(海拔、坡度、坡向、土层厚度)和次生林恢复时间(林龄)对生物量、生产力、物种丰富度与生物量和生产力间关系的影响。结果表明, 当不考虑环境因子时, 物种丰富度与生物量之间存在显著的线性正相关关系, 而与生产力之间存在显著的二次关系(先增加后减少的驼峰型)。当考虑环境因子时, 个体密度和土层厚度对生物量具有显著影响, 而环境因子对生产力并无显著效应。在坡度较陡、坡向朝南及土层较厚的环境条件下, 物种丰富度与生物量具有显著的线性正相关关系; 而在坡度较缓、坡向朝北及土层较薄的环境条件下, 物种丰富度不影响生物量。在较高海拔环境条件下, 生产力随物种丰富度先增加后减少(驼峰形状), 而在其他环境条件下, 生产力均不响应物种丰富度。以上结果说明自然森林生态系统中物种丰富度与生物量和生产力的关系存在差异, 且其相互间的关系依赖于环境因子。
13C and 15N isotopic signatures of plant-soil continuum along a successional gradient in Dinghushan Biosphere Reserve
DOI:10.17521/cjpe.2015.0478 URL [本文引用: 1]
鼎湖山森林演替序列植物-土壤碳氮同位素特征
DOI:10.17521/cjpe.2015.0478
[本文引用: 1]
植物群落对水分利用和养分利用的优化策略, 土壤碳周转和氮循环过程对演替变化如何响应, 森林土壤有机碳积累机制等都是森林生态学需要解决的关键问题。然而, 这些生态学过程的变化在短时间内通过传统的研究手段难以被精确观测, 碳氮同位素(<sup>13</sup>C、<sup>15</sup>N)技术的应用或许能提供更多有价值的信息。该文通过对鼎湖山森林演替序列代表性群落——马尾松(Pinus massoniana)针叶林(PF)、针阔叶混交林(MF)和季风常绿阔叶林(BF)植物-土壤碳氮同位素自然丰度的测定, 分析了叶片稳定碳同位素比率(δ<sup>13</sup>C)和稳定氮同位素比率(δ<sup>15</sup>N)与其叶片元素含量的关系, 以及叶片-凋落物-土壤δ<sup>13</sup>C、δ<sup>15</sup>N在演替水平和垂直方向上的变化特征。结果显示: 1)主要优势树种叶片δ<sup>13</sup>C与其C:N极显著正相关(p < 0.01), 凋落物和各层土壤δ<sup>13</sup>C均表现为PF > MF > BF, 沿演替方向逐渐降低; 2)叶片δ<sup>15</sup>N与叶片N含量正相关(p = 0.05), 凋落物和表层土壤(0-10 cm) δ<sup>15</sup>N沿演替方向逐渐增大; 3)不同演替阶段土壤δ<sup>13</sup>C、δ<sup>15</sup>N均沿垂直剖面呈现增大的趋势。结果表明: 南亚热带地区植物群落的发展并不一定受水分利用和氮素利用的补偿制约; δ<sup>13</sup>C自然丰度法的应用有助于森林土壤有机碳积累机制, 尤其有助于成熟森林土壤“碳汇”机制的阐释; 植物-土壤δ<sup>15</sup>N值可作为评估土壤氮素有效性和生态系统“氮饱和”状态的潜在指标。
Does complementary resource use enhance ecosystem functioning? A model of light competition in plant communities
DOI:10.1111/j.1461-0248.2006.00994.x URL [本文引用: 1]
Abiotic and biotic determinants of coarse woody productivity in temperate mixed forests
DOI:10.1016/j.scitotenv.2018.02.125 URL [本文引用: 1]
Individual size inequality links forest diversity and Above-ground biomass
DOI:10.1111/1365-2745.12425 URL [本文引用: 2]
Forest productivity increases with evenness, species richness and trait variation: A global meta-analysis
DOI:10.1111/j.1365-2745.2011.01944.x URL [本文引用: 1]
Spatial heterogeneity of soil organic carbon and total nitrogen in a monsoon evergreen broadleaf forest in Dinghushan, Guangdong, China
鼎湖山季风常绿阔叶林土壤有机碳和全氮的空间分布
Long-term (
DOI:10.17521/cjpe.2017.0171 URL [本文引用: 4]
1992-2015年鼎湖山季风常绿阔叶林群落结构动态
DOI:10.17521/cjpe.2017.0171
[本文引用: 4]
季风常绿阔叶林是中国南亚热带地区的地带性植被, 其群落结构及其动态变化直接影响着该森林能为该地区提供的生态系统服务功能质量。该文利用1992-2015年共24年的长期定点监测数据, 从群落种类组成、生物量、径级、密度等数量特征方面研究了鼎湖山季风常绿阔叶林群落结构的动态变化。结果表明: 1)到2015年该森林群落林分个体密度增加了42.7%, 总生物量减少了5.1%; 24年间基于生物量的β多样性指数群落差异为37.4%, 基于个体数的差异则高达65.6%; 2)灌木和小乔木的个体数剧增, 生物量增大, 中乔木和大乔木的个体数变化虽不显著, 但生物量显著降低; 3)小径级(胸径Aidia canthioides)、鼎湖血桐(Macaranga sampsonii)、柏拉木(Blastus cochinchinensis)等物种个体数急剧增加, 锥(Castanopsis chinensis)、木荷(Schima superba)、黄果厚壳桂(Cryptocarya concinna)等树种生物量大量减少, 白颜树(Gironniera subaequalis)、窄叶半枫荷(Pterospermum lanceifolium)等树种生物量增加, 这些物种是群落结构变化的主要贡献者。鼎湖山季风常绿阔叶林群落结构在1992-2015年发生了巨大变化, 与演替、虫害和气候变化等影响有关。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}