


数千年来文明的发展伴随着人类对生态系统的不断改变。进入到人类世(Anthropocene)时期以来, 人类活动规模和强度的不断增加对全球生态系统的组成、结构和功能都产生了深远的影响(Steffen et al, 2007)。2019年美国生态学会成立百年之际将“变化世界中的生态学”定为主题, 反映了生态系统对全球变化的响应和反馈成为当前生态学的核心议题之一。
生态系统可能表现出多种变化轨迹, 典型的有: (1)线性变化, (2)具有临界阈特征的非线性变化, 以及(3)具有迟滞(hysteresis)特征的突变(Scheffer et al, 2001)。其中具有迟滞特征的生态系统突变近年来受到了广泛关注。这种变化的共性特征为: 原先稳定的生态系统会在短时间内发生难以预料的急剧退化甚至崩溃, 退化后的系统状态也具有较高的稳定性且往往难以恢复(Scheffer et al, 2001)。近年来频发的大规模森林丧失、草原退化、湖泊水生植被消亡和珊瑚礁白化等事件表明, 生态系统一旦发生这种突变, 将威胁到整个区域乃至全球的生态安全, 并引发严重社会问题。生态系统的突变吸引了学界和公众的重点关注, 在英国生态学会所列出的当前“100个生态学基础问题”中, 就有多个问题与生态系统的突变密切相关(Sutherland et al, 2013)。
近年来生态文明建设与可持续发展已被提升到国家战略地位。在生态系统退化与修复的理论和实践研究中, 对生态系统突变的关注程度不断增加。本文试图对生态系统多稳态和突变的相关研究历史和部分前沿进展进行梳理, 以期为我国生态系统突变的应对与管理提供参考。
1 理论基础
许多不同类型的生态系统在特定条件下都可能发生状态突变。这些生态系统虽然具有极大差异, 但突变却往往遵循相似的轨迹(Scheffer et al, 2001): 随着外部环境条件(如降水、营养条件等)的逐渐变化, 生态系统状态(如植被盖度、生物量等)在初期可以保持相对稳定, 表现出相对较小的变幅; 但在越过某临界阈值后, 生态系统状态变量则在短时间内发生大幅变化(如生物量大幅下降), 表现出突变特征; 而且突变后的生态系统也可以保持相对稳定, 难以恢复到突变前的状态。这种变化常称为稳态转换(regime shift), 一般表现为突变前后状态维持的时间远长于突变发生的时间。
多稳态(alternative stable states)理论在20世纪70年代早期开始出现于生态学研究中, 在解释上述生态系统突变行为的机理上获得了大量理论和实证证据的支持(Scheffer et al, 2001; Beisner et al, 2003; Levin et al, 2012)。多稳态理论的数学基础可以追溯到分岔理论(bifurcation theory)和突变理论(catastrophe theory)。在基于微分方程描述的动力系统(dynamical systems)中, 有时参数的微小连续变化可以导致系统行为(或拓扑属性)在性质上发生突然变化, 称为分岔(bifurcation)。分岔行为有多种类型。其中一种相对简单的鞍结点分岔(saddle-node bifurcation, 离散系统中又称为fold bifurcation)描述了随着参数变化, 系统的多个平衡点(equilibrium)合并进而消失的过程。例如图1A中所示的系统, 随着控制参数(横坐标)逐渐变化, 系统的稳定平衡点(stable equilibrium, 实线上)和不稳定平衡点(unstable equilibrium, 虚线上)在F1或F2 (分岔点)处合并进而消失。
图1
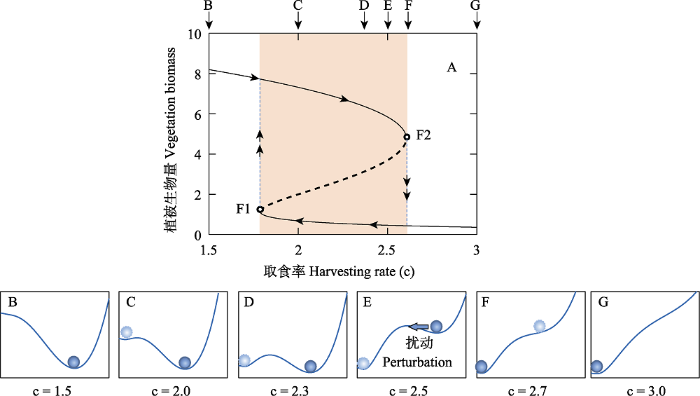
图1
基于放牧模型的多稳态理论的图示(修改自Kéfi et al, 2013)
Fig. 1
The graphic model of the alternative stable states based on grazing model (Adapted from Kéfi et al, 2013)
| Box 1 放牧系统简单模型 |
|---|
| Noy-Meir (1975)提出的放牧系统简单模型是生态系统多稳态的经典理论模型。该动力系统模型仅考虑植物生长和食草作用两种基本过程, 由如下微分方程表示: $\frac{\text{d}V}{\text{d}t}=rV\left( 1-\frac{V}{K} \right)-c\frac{{{V}^{p}}}{{{V}^{p}}+{{V}_{0}}^{p}}$ (1) 其中, $rV\left( 1-\frac{V}{K} \right)$刻画了植物生物量的逻辑斯蒂增长过程, 而$c\frac{{{V}^{p}}}{{{V}^{p}}+{{V}_{0}}^{p}}$刻画了食草动物的消费过程。V是植物生物量, r为植物的内禀增长率(这里取值为1), K为植物的环境承载力(这里取值为10), c为食草动物的最大取食率, $\frac{{{V}^{p}}}{{{V}^{p}}+{{V}_{0}}^{p}}$为Holling的III型功能响应函数(S型曲线, V0为半饱和参数, p这里取值为2)。 如 稳态转换可以用“山谷中的小球”形象表示。如 |
动力系统分岔行为的详细数学解释可参见相关教材(Strogatz, 2018), 本文主要从直观角度来阐述现实生态系统的多稳态理论解释。一般而言, 某些生态系统在给定的外部环境条件(如温度、降水、营养条件、放牧强度等, 对应于动力系统的参数, 即图1A中的横坐标)下可能存在多种稳定状态(简称稳态, 对应于动力系统的多个稳定平衡点, 即图1A中的实线; 稳态之间由虚线所示的不稳定平衡点分隔开来)。当生态系统处于其中一种稳态, 随着外部环境条件(一般为慢变量)的逐渐变化(如图1中沿箭头所示方向), 到达F1或F2分岔点后, 该稳态消失, 系统状态(一般为快变量)落在另一种稳态上。由于多种稳态之间为不连续分布(如图1中两条实线所示的两种稳态为不连续分布), 这种稳态转换通常表现为突变。由于F1和F2分岔点在环境梯度上的位置具有明显差异, 稳态转换表现出明显的迟滞特征。迟滞特征解释了为何即使环境条件改善到退化(正向稳态转换)发生时的状况, 生态系统也难以恢复(逆向稳态转换) (Beisner et al, 2003; Scheffer, 2009)。
对于真实的生态系统而言, 干扰(如火烧、病虫害、飓风、洪水等)的发生几乎不可避免。当系统存在多稳态时, 即使外部环境条件保持不变, 当干扰的强度足够大时也可能驱动系统状态跨越不稳定平衡点而落入到另一种稳态上并表现出突变行为。这种稳态转换可用“山谷中的小球”概念模型进行直观描述(图1B-G): 小球处于谷底(表征动力系统中的吸引域, basin of attraction)表示系统处于稳态, 受到小幅干扰时系统偏离稳态(小球处于山坡位置), 但干扰消除后可恢复到稳态(小球恢复到谷底位置); 当扰动足够强烈时, 小球可能越过山脊(不稳定平衡点)进入相邻的山谷, 即系统落入另一种稳态(Scheffer et al, 2001, 2012a)。
概括而言, 稳态转换的发生可能有两种不同的内在机理: 一是外部环境条件的改变导致系统“内在”稳定性丧失(势能景观发生变化, 主要表现为吸引域变小), 二是干扰导致系统跨越不稳定平衡点(势能景观未发生变化)。在现实生态系统中, 两种变化往往同时发生: 环境变化导致系统的稳定性下降, 导致干扰更容易触发稳态转换。
为何生态系统会表现出多稳态特征?一般认为, 围绕生态系统状态变量产生的正反馈(positive feedback)是产生多稳态的必要但非充分条件 (Scheffer, 2009)。正反馈环路的形成可能由生物之间的互作或生物与环境之间的互作而反馈于自身。通常只有当正反馈作用的强度足够高时, 生态系统才可能表现出图1所示的多稳态特征(Kéfi et al, 2016)。从直观上来解释, 系统在负反馈作用下可以保持局部稳定并形成稳态; 但在全局角度上, 正反馈作用可以对系统稳态的微小偏离产生放大效应, 从而倾向于使得整个系统失稳(DeAngelis et al, 1986)。生态系统中具体的正反馈机制将在小节3中结合现实案例进行详述。
2 识别方法与早期预警
2.1 识别方法
在理论研究中, 利用模型识别系统的多稳态以及分岔行为相对容易, 本文不再赘述。对于现实生态系统而言, 也许永远无法(像在数学和模型研究中那样)严格证明多稳态是否存在, 但可以通过实验、观测和模型等途径来推测生态系统特征与行为是否符合多稳态理论预测(Scheffer & Carpenter, 2003; Schröder et al, 2005)。
在严格控制的微宇宙实验条件下, 相关研究以浮游生物、细菌等微生物为对象, 通过扰动实验等方法发现微生态系统在趋近突变临界点的行为符合(鞍结点)分岔理论的预期, 获得了多稳态的有力证据(Drake & Griffen, 2010; Dai et al, 2012, 2013; Veraart et al, 2012)。此外, 以浅水湖泊作为中宇宙尺度的全湖实验(whole-lake experiments)表明其动力学特征与多稳态的理论预测吻合(Carpenter et al, 2011)。一般而言, 在宏观时空尺度上开展控制实验通常较为困难, 基于小尺度控制实验和实地格局观测(自然实验)的研究途径较为常见。如果在实验/自然生态系统中出现以下模式, 则可能暗示该系统存在多稳态(Scheffer & Carpenter, 2003; Schröder et al, 2005; Scheffer, 2009):
(1)生态系统的关键状态变量在相似环境条件下表现出多峰分布特征。“稳态”并不意味着生态系统会到达绝对的稳定。现实生态系统受到多种外部扰动或季节波动而一直表现出变化特征; 另一方面, 即使环境保持稳定, 许多生态系统由于内在作用也可能表现出持续波动特征(如混沌、极限环、奇异吸引子等动力学特征)。但生态系统往往围绕在特定状态(吸引子, attractor)附近波动, 此时生态系统的稳态是一种动态的“体系” (regime); 相应地, 状态变量表现出的多个分布(一般为正态分布)可能是多个稳态体系存在的表征, 其峰值(众数)的位置对应于吸引子。当数据量充分的时候, 可以通过“势能分析” (potential analysis)来构建系统的稳定性景观(stability landscape, 也称势能景观, potential landscape)用于直观/定量表征系统多稳态的吸引域(Livina et al, 2010; Hirota et al, 2011; Xu et al, 2016; Scheffer et al, 2018)。
(2)生态系统状态在时间序列上短期内发生剧烈变化。这种特征是生态系统发生稳态转换的一种可能表现形式。但需要注意的是, 许多情况下系统状态的急剧变化可能是由于外部环境条件的突变(有时某些外部环境因子的剧变未必容易观测到)或其他因素导致, 其本质并不是动力系统的分岔行为。虽然有相关的时间序列分析方法可以通过数据本身来推测系统行为是否是稳态转换, 但往往对数据质量(时间序列的长度和分辨率)有较高的要求。
(3)时间序列或空间格局上的状态突变之前表现出某些早期预警信号。理论上, 动力系统趋近鞍结点分岔点之前, 系统会表现出临界慢化(critical slowing down)现象。研究发现基于临界慢化的早期预警信号在许多现实生态系统发生稳态转换之前表现出明显的指示特征(详见小节2.2)。
(4)实验中生态系统的最终状态(稳态体系)依赖于初始条件。如图1所示, 在给定环境条件下, 具有多稳态的生态系统最终表现为哪种稳态取决于其初始状态落在不稳定平衡点的上方还是下方。如果实验条件下能够发现这种初始条件依赖现象, 则暗示系统可能具有多稳态特征。
(5)干扰可以触发系统转变为明显不同的(类似稳态体系的)状态。如前所述, 对于存在多稳态的系统, 当干扰强度足够大时, 可以推动系统跨越不稳定平衡点落在另外的稳态上。因此现实生态系统的这种行为可以视为多稳态的证据之一。
(6)系统的正向和逆向变化存在迟滞现象。现实生态系统如果随着环境条件的正向和逆向变化发生状态剧变的临界条件具有明显差别, 也可以视为多稳态的证据之一(如图1所示)。
需要注意的是, 由于上述模式都可能由其他机制驱动形成, 因此都不能视为多稳态的绝对证据。但如果多种模式同时存在于某个生态系统中, 则多稳态的存在具有较高的可能性。
2.2 早期预警信号
生态系统突变前的状态往往变化幅度相对较小, 因而从表面状态很难判断突变的发生。另外, 在到达稳态转换临界阈前, 小幅干扰即可触发稳态转换, 意味着随机发生的干扰可以在接近临界阈前的任意时间触发突变, 因此几乎无法精确预测生态系统的突变发生事件, 但可以根据系统行为特征评估系统发生突变的风险(或脆弱性)。根据多稳态理论, 随着外部环境条件的逐渐变化, 生态系统趋近分岔点时会出现临界慢化特征, 表现为吸引域变小(如图1所示, 山谷的宽度、深度以及坡度变小), 扰动后恢复到稳态的时间变长。此时表现为系统的“弹性” (resilience, 有国内学者翻译为恢复力或韧性, 用于表征生态系统在环境变化时维持自身完整性和功能的能力)下降, 系统跃迁到另一种稳态的风险增加(Scheffer et al, 2009, 2012a, 2015a; Dakos et al, 2015)。
近年来, 寻找可指示各种生态系统突变的“通用”早期预警信号(generic early-warning signals)研究取得了一系列重要进展(Scheffer et al, 2009, 2012a, 2015a; Dakos et al, 2015)。理论研究表明, 当接近临界阈时, 不同系统具有某些共通的行为特征, 主要表现在扰动发生后恢复到稳态的过程将变得缓慢, 即系统弹性下降, 其数学基础是动力系统中雅克比矩阵的特征值趋于0 (即临界慢化现象)。这种变化可以被多个指标有效捕捉到, 主要包括: 系统受到扰动后的恢复时间变长、系统状态参量在时间序列上的自相关程度和方差上升、系统频谱发生红移等。因此, 扰动后的恢复时间、时间序列上的一阶自相关系数、时序方差(或变异系数)等都可以成为指示突变的重要早期预警信号。如果外部环境频繁波动, 接近临界阈时系统在不同状态之间来回切换(状态“闪烁”, flickering), 对于这种系统, 时序偏斜度(三阶矩)、翘度(四阶矩)、概率分布的不连续性也是有效的早期预警信号。这些指标在诸多现实生态系统中都得到了有效的验证, 但由于需要较长时间序列数据的支持, 在生态系统管理中的实用性有限。考虑到生态系统通常由空间单元构成, 相比单一的状态参量(每个时间点通常只有一个数据), 充分利用空间格局数据可能提供更多的信息用于突变预测。在这种思路的指引下, 研究发现系统状态的空间自相关程度、空间方差和偏斜度等也是明确的早期预警信号。虽然早期预警信号的理论研究取得了重要成果, 但仍存在局限性, 相关实证研究也有待深入: (1)并非所有生态系统都具有多稳态和稳态转换机制。很多生态系统未必具有多稳态, 但也表现出临界阈特征。对于这种系统而言, 部分早期预警信号仍然适用, 但必须进行针对性检验。(2)部分早期预警信号在应用于现实生态系统中可能失效, 出现假警报或未警报情况, 其原因目前尚未完全澄清。而部分早期预警信号的有效性仍存在争议。具体的早期预警信号的计算方法及其应用案例可参见相关综述(Scheffer et al, 2015a)。
3 现实案例
多稳态理论被广泛用于阐释多种生态系统(以及多种其他复杂系统)的突变机理。可能具有多稳态的生态系统包括浅水湖泊、干旱/半干旱区生态系统、森林-稀树草原生态系统和多种海洋生态系统。本文选择具有代表性的几种生态系统进行具体阐述。需要特别注意的是, 虽然多稳态理论描述的情形相对简单, 但将该理论应用到复杂的现实生态系统时, 往往伴随着诸多与具体机理、数据、方法等方面相关的复杂问题, 因此需要避免简单移植或套用理论框架。
3.1 浅水湖泊
在多稳态的早期研究中, 浅水湖泊作为代表性的生态系统受到了重点关注(Scheffer et al, 1993; Scheffer, 1997)。由于人源排放的增加, 氮、磷等营养元素的过度输入导致水体富营养化, 浅水湖泊通常表现出由沉水植物占主导的“草型状态” (通常表现为水体浑浊度较低的清水状态)迅速退化为浮游藻类占主导的“藻型状态” (通常表现为水体浑浊度较高的浊水状态); 而如果要恢复到原先状态, 必须将营养负荷削减到远低于突变前的水平, 表现出明显的迟滞特征。大量理论和实证研究表明, 浅水湖泊的这两种状态本质上很可能是多稳态的表现形式。其证据包括基于实地观测和控制实验的一系列实证(Scheffer & Carpenter, 2003): (1)浅水湖泊生态系统状态的频度分布通常表现出“双峰”特征(bi-modal distribution), 即清水状态和浊水状态频度相对较高, 而中间状态的频度相对较低(图2); (2)在长时间序列上, 清水状态和浊水状态之间的转换发生在相对较短的时间内, 表现出明显的突变特征; (3)系统状态随营养条件的变化表现出明显的迟滞特征; (4)在全湖尺度上的自然或人为扰动实验下, 湖泊也表现出明显的突变特征。在温带和亚热带湖泊中都观测到类似模式, 但不同气候条件下的模式和机理可能存在明显差异(Scheffer, 1997; Scheffer & van Nes, 2007; Pan et al, 2015)。
图2
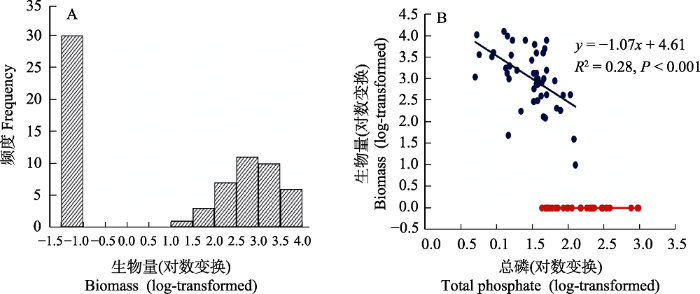
图2
长江浅水湖群沉水植物生物量的不连续分布可能是浅水湖泊生态系统的多稳态证据。(A)沉水植物生物量的频度分布表现出不连续的多峰分布(修改自王海军, 2007); (B)在水体总磷含量的梯度上沉水植物的生物量表现出不连续特征(修改自Wang et al, 2014)。
Fig. 2
Discontinuous distribution of submerged plant biomass in the Yangtze shallow lakes as suggestive evidence of alternative stable states of shallow lake ecosystems. (A) Submerged macrophyte biomass shows multi-modal frequency distribution (Adapted from Wang et al, 2007); (B) Two regimes of submerged plant biomass are present along the phosphorus gradient (Adapted from Wang et al, 2014).
浅水湖泊中存在若干种可能导致多稳态形成的正反馈机制(Scheffer & van Nes, 2007), 例如: (1)沉水植物通过抵御风浪降低沉积物再悬浮、与藻类竞争光和营养、化感作用以及为浮游动物提供庇护所进而抑制藻类等途径提升水体透明度, 进而为沉水植物自身的进一步发展提供优良的条件, 加强了清水态的稳定性。(2)沉水植物退化后, 沉积物再悬浮加强, 藻类因竞争和牧食压力降低而大量发展, 透明度进一步降低, 抑制沉水植物的恢复, 加强了浊水态的稳定性。基于长江亚热带浅水湖群的系列研究提出了一些新的补充机制: 沉水植物死亡后水体中氮浓度变化不大而磷浓度大幅上升, 这为缺磷的水体提供了很好的营养来源, 进一步促进了藻类发展(Wang et al, 2014); 沉水植物死亡后, 原先摄食水草上附着藻类和无脊椎动物的鱼类将转向在底部摄食, 进而加大沉积物的再悬浮(Pan et al, 2015)。这两种机制均可增加水体的浊度进而抑制沉水植物的恢复, 这可能是导致浅水湖泊生态系统弹性下降的一种重要机理。此外, 浅水湖泊中沉水植物具有不同的功能性状(如形态特征和化学计量内稳性等)可能对于维持湖泊清水态的能力产生差异(反馈作用强度不同), 在湖泊富营养化过程中通过改变沉水植物群落结构也可能降低生态系统弹性(Su et al, 2019a, b)。
近年来我国学者对以浅水湖泊为代表的湿地生态系统多稳态和稳态转换现象和机理开展了系统性的研究工作, 详见相关综述(冯剑丰等, 2009; 李玉照等, 2013; 吴庆龙等, 2013; 赵磊等, 2014; 赵雁捷等, 2016; 李蕙等, 2017)。
3.2 陆地植被
经典理论(如群落演替的顶极理论)认为在生物地理尺度上, 植被类型主要决定于气候、土壤等宏观环境因子。但近年来越来越多的证据表明, 在某些条件下, 不同植被类型可能表现为多稳态特征。在热带地区, 基于全球尺度数据的分析结果表明, 树木盖度(Hirota et al, 2011; Staver et al, 2011)和冠层高度(Xu et al, 2016, 2018; Hantson et al, 2017; Scheffer et al, 2018)的频度分布表现出显著的不连续多峰分布(multi-modal distribution)格局; 同时, 基于树木胸径、树皮厚度、比叶面积等植物性状(Dantas et al, 2013, 2016)的地面观测结果也反映出一致的多峰分布格局, 这些分布的众数(峰值)分别反映了典型热带森林、稀树草原和荒漠/草原植被特征。这种在渐变的环境梯度上植被表现出的不连续分布特征与多稳态理论预测相吻合。此外, 实地观测结果发现, 热带森林在受到干旱、火烧等干扰后可能退化为具有稀树草原特征的稳定生态系统, 也与多稳态理论预测相符(Flores et al, 2016, 2017; Staal et al, 2018b; van Nes et al, 2018)。
理论模型与实证研究表明, 火烧干扰所形成的正反馈机制可能在塑造热带植被的多稳态中扮演了关键角色。局部尺度上, 热带森林中广泛存在的正反馈机制为: 林冠层郁闭→林下透光度低→林下草本生长受限→草本可燃生物量不足→火烧难以发生→树木冠层保持郁闭(系统表现为郁闭森林状态)。但严重的干旱或火烧干扰一旦发生并导致林冠层盖度显著下降, 则会产生以下正反馈环路: 林冠层稀疏→林下透光度增加→促进林下草本生长→草本可燃生物量高→火烧易于发生→乔木幼苗受火影响死亡率上升→乔木更新不良→林冠层保持稀疏(系统表现为稀树草原状态) (Staal & Flores, 2015; Staal et al, 2016; van Nes et al, 2018)。在较大空间尺度上也存在重要的正反馈机制, 例如植被盖度的增加可以通过促进本区域水分循环等方式促进森林的维持(Staal et al, 2020)。
此外, 动物食草作用、阿利效应、土壤条件、霜冻等因素都可能在特定条件下介导形成正反馈机制, 进而导致植被系统多稳态的形成(van Langevelde et al, 2003; Xu et al, 2015a; Kéfi et al, 2016; Joshi et al, 2020)。
近期研究在寒带和温带地区植被中也发现了与热带植被相似的系统状态不连续多峰分布格局(Scheffer et al, 2012b, 2014, 2015b; Xu et al, 2015b, d), 暗示全球尺度上植被的多稳态可能是一种普遍存在的特征。对于寒带植被, 有研究猜测火烧、病虫害暴发、冻原融化等生态过程可能产生重要的正反馈机制从而产生多稳态(Scheffer et al, 2012b; Abis & Brovkin, 2019), 但相关实证和机理研究仍有待深入开展。
宏观尺度上陆地植被的多稳态研究对于深入理解和预测全球变化的生态后果具有重要价值。多稳态理论预测, 寒带暖化加剧和热带干旱加剧等全球气候变化可能导致一系列生态系统发生稳态转换, 热带雨林退化、寒带森林丧失、冻原加速融化等宏观生态系统一旦发生突变, 可能在短期内导致地表和地下碳库的加速释放, 并对全球气候变化产生重要影响。
3.3 干旱区生态系统
在干旱胁迫环境条件下, 水分通常为最重要的限制性生态因子。在微生境尺度上, 植物生长可以通过冠层遮荫减少蒸散失水, 同时通过根系作用促进下方土壤疏松, 从而增加降水入渗和横向地表径流来源的水分下渗, 最终提升土壤水分有效性; 此外, 植物地上部分的生物量也可以截留风力搬运的(含有养分的)沉积物, 而根际微生物活动也能够促进养分有效性。总体上, 植物生长有助于其下方土壤水分和养分条件的改善, 从而有利于维持自身生长, 形成局部正反馈作用(Holmgren et al, 1997; Holmgren & Scheffer, 2010)。反之, 一旦植被丧失, 土壤失去植被保护作用后, 土壤侵蚀程度增加, 土壤表面形成硬质结皮增加地表径流而减少水分下渗, 进一步形成不利于植物生长的微生境条件, 在这种正反馈作用下, 裸地状态得以稳定维持。这些由植物-土壤相互作用介导的正反馈机制导致微生境尺度上形成两种稳态: 植被斑块状态与裸地斑块状态(D’Odorico et al, 2007), 在干旱区, 这两种状态在空间上通常镶嵌分布(受到尺度依赖的反馈机制作用), 成为干旱区植被景观的标志性特征(van de Koppel et al, 1997; Rietkerk et al, 2004)。对此类正负反馈形成的有序空间格局的进一步理解, 读者可参阅3.4以及本期有关自组织(self-organization)理论的详细阐述(葛振鹏和刘权兴, 2020)。
此外, 植物个体间的局部促进作用(如保育植物作用, nurse plant effect)所产生的正反馈作用的强度如果足够大, 可能导致景观水平上系统产生植被覆盖状态与裸地状态的多稳态特征(Xu et al, 2015a, c)。例如, 在荒漠化区域中, 植物个体间通过土壤条件改善、遮荫效应和食草动物防御等机制产生的促进作用能够形成正反馈环路, 促进植被覆盖的增加, 发达的植物根系将沙丘固定, 最终形成稳定的固定沙丘状态; 而在缺少植物定植的流动沙丘上, 植物幼苗易被流沙掩埋而具有较高的死亡率, 植物生长受限使得沙丘缺少根系固定作用, 这种正反馈机制使得沙丘表现出持续的流动性。流动沙丘与固定沙丘之间的稳态转换受到气候变化和放牧活动等过程驱动, 环境重建数据表明, 在长时间尺度上, 稳态转换在两种沙丘类型之间可能反复发生(Xu et al, 2020); 而植被在退化和恢复过程中都可能表现出突变特征, 并在到达突变点前表现出早期预警信号(Chen et al, 2018)。
3.4 自组织生态系统
如前所述, 正反馈机制是生态系统产生多稳态的必要条件。正相互作用存在于许多生态系统的正反馈环路, 而正相互作用也是许多生态系统自组织空间格局产生机理之一(并非唯一机制) (Kéfi et al, 2016)。在自组织系统中, 正反馈能使原来平稳的生态系统, 在环境胁迫下发生突变并产生有序的空间自组织格局(表现为规则的空间斑图)。无论原来的生态系统是单稳态还是双稳态系统, 图灵(Turing)理论(以尺度依赖的反馈为核心机理)框架下的自组织发生均能够导致多稳态的涌现(图3) (van de Koppel & Crain, 2006)。此外, 规则斑图本身的特征变化有时也可以作为指示生态系统趋近稳态转换临界点的早期预警信号(Rietkerk et al, 2004)。
图3
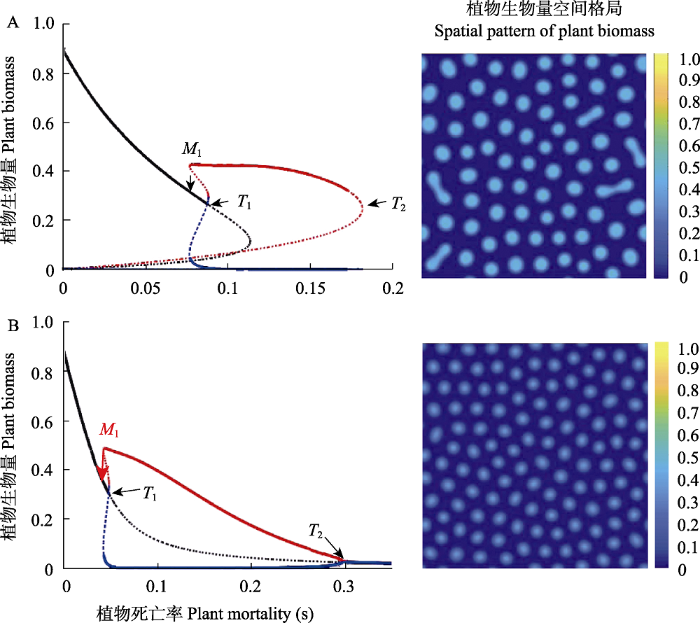
图3
一个淡水沼泽湿地生态系统中产生的植物空间自组织格局与稳态转换。当系统中存在促进作用时可能同时产生多稳态和自组织空间格局特征。(A)和(B)分别是
Fig. 3
Spatial self-organization and alternative stable states of the plants in a freshwater wetland ecosystem. Facilitative effects may give rise to alternative stable states and spatial self-organization signature in ecosystems. (A) and (B) are regime shift and regular spatial patterns generated by the model 2 and 3 in table 1, respectively. M and T are bifurcation points (Adapted from van de Koppel & Crain, 2006).
图3所示的一个典型的淡水沼泽湿地生态系统模型可用于说明自组织系统诱导多稳态发生的机理(表1, 图3)。该系统由植被(变量P)和枯落物(变量W)相互作用组成, 其中植物种群的增长符合一般逻辑斯谛方程。植物增长受到枯落物的非线性抑制而减少, 枯落物的生物量来自植被生命周转速率。当植物之间没有正反馈作用时(模型1), 该系统将呈现空间均匀低生物量的平衡态, 即没有自组织格局涌现。当该系统中引入直接的正反馈作用(模型2), 则系统将会出现规则的图灵斑图, 同时表现出多稳态特征: 随着植物死亡率(s)增加到达M1点时, 植被生物量会突然增加, 并且通过自组织形成有序缺口格局(图灵斑图); 随着死亡率进一步增加, 因为系统的自组织涌现功能会延长系统的持续性和稳健性, 直至到达T1点系统崩溃(图3A)。该系统过程反映出自组织过程通过延迟系统的稳态转换的临界值从而增强了系统的稳健性, 即降低了系统发生崩溃突变的风险, 这可能是自组织的重要生态功能之一(Zhao et al, 2019)。当该系统存在间接正反馈过程时(模型3), 自组织同样能够诱导稳态转换发生, 然而前面出现的临界崩溃现象消失。此时尽管系统的脆弱性降低, 但其持续性并没有得到提升(图3B)。如何通过自组织过程和行为来研究生态系统的弹性和早期预警信号依然有待进一步探索。
表1 一个淡水沼泽湿地生态系统中产生的植物空间自组织格局与稳态转换的模型
Table 1
| 模型方程 | 反馈机制 | |
|---|---|---|
| 植被生物量模型1 Vegetation biomass model 1 | $\frac{\partial P}{\partial t}=P\left( 1-P \right)-iWP-sP+{{D}_{P}}\Delta P$ | 抑制 Inhibition |
| 植被生物量模型2 Vegetation biomass model 2 | $\frac{\partial P}{\partial t}=P\left( 1-P \right)\frac{P}{P+{{k}_{2}}}-iWP-sP+{{D}_{P}}\Delta P$ | 促进 + 抑制 Facilitation + inhibition |
| 植被生物量模型3 Vegetation biomass model 3 | $\frac{\partial P}{\partial t}=P\left( 1-P \right)-\frac{{{k}_{3}}}{P+{{k}_{3}}}iWP-sP+{{D}_{P}}\Delta P$ | 促进 + 抑制 Facilitation + inhibition |
| 枯落物生物量 Wrack biomass | $\frac{\partial W}{\partial t}=sP-bW+{{D}_{W}}\Delta W$ | |
3.5 群落组成
作为群落生态学和生物多样性研究的核心问题, 群落构建的过程可能表现出多稳态特征。例如, 多稳态可能在经典的优先效应(priority effect)、历史偶然性(historical contingency)等塑造群落物种组成的作用中扮演关键角色(Fukami, 2015), 也有助于阐释环境随机作用对群落的塑造作用, 以及生态学早期的群落“整体论” (如以不连续物种组成为证据)和“个体论” (以连续物种组成为证据)争论产生的本质(Liautaud et al, 2019)。群落多稳态(alternative community states)的概念也受到了越来越多的关注。在物种多样性较低(如干旱区植物群落)或优势种具有明显差别(如岩石质潮间带生物群落)的群落中, 当通过自然实验或者控制实验途径在相似环境条件下观测到群落组成具有显著区别时, 有研究者将其视为群落多稳态存在的证据(Petraitis & Dudgeon, 2005)。但由于多稳态的直接证据通常难以获得并受到时空尺度等因素的影响, 关于特定系统中群落是否具有多稳态仍存在争议(Bertness et al, 2002)。对于物种丰富的群落(如森林生态系统), 多稳态的识别往往更为困难。
许多机理可能产生物种组成具有明显差别的群落, 但这些机理未必都与系统的分岔行为有关。对于以鞍结点分岔为基础的系统, 正反馈机制在产生群落多稳态的过程中发挥了关键性的作用, 可能的正反馈机制包括生物的环境塑造作用(如植物-土壤反馈机制)以及生物个体间的促进作用等。此外, 物种之间的竞争等负相互作用也可能产生间接的正作用并导致产生群落组成的不连续特征(Liautaud et al, 2019)。总体上, 关于群落组成与生物多样性的多稳态表征和机理研究还有待于进一步深入。
4 理论应用
多稳态理论目前已被应用于多种现实生态系统的管理实践中。特别是近年来多稳态理论对生态修复的实践产生了重要的影响。以往生态修复主要关注于恢复生态系统退化之前的环境条件, 但对于存在多稳态和迟滞特征的系统而言, 诸多研究表明在驱动系统状态自我维持的正反馈机制的作用下, 即使外部环境条件恢复后系统通常也难以复原到退化前的状态。因此通过人为干预, 打破或减弱退化系统状态的正反馈机制成为生态修复成功的关键环节(Suding et al, 2004)。
基于稳态转换理论的“休克疗法” (即去除湖泊中大部分鱼类以减弱维持浊水状态的正反馈作用, 从而促进向清水态转换的发生)在欧洲开展的众多浅水湖泊生态修复中取得了显著的成效(Jeppesen et al, 2012)。广东惠州西湖也是一个成功案例, 通过去除鱼类, 减少对沉积物的扰动来提升水体透明度, 最终达到恢复沉水植物建成健康稳态的目标(Jensen et al, 2017)。湖北枝江金湖是另外一个成功案例(该湖初始的湖水总磷浓度远高于藻-草转换的阈值(80-100 μg/L) (Wang et al, 2014)。该湖采取生态水文技术大幅降低水位(徐超等, 2020), 使得水底光强能满足沉水植物生长的需求进而实施生态工程, 在一年之内沉水植被覆盖率60%-80%, 透明度得到大幅提升。在南美洲开展的干旱区植被退化生态修复中, 研究者发现在厄尔尼诺周期内降水增加时开展人工植被恢复, 可有效推动裸地稳态向植被覆盖稳态的转换(Holmgren & Scheffer, 2001), 即借助降水丰沛的“机会窗口”通过“逆向扰动”可实现退化生态系统向健康生态系统转换。我国干旱区防风固沙工程中广泛采用的“草方格”技术(Li et al, 2004), 通过在裸沙表面覆盖秸秆而增加微生境的水分和养分有效性等途径来打破不利于植物定植的正反馈环路, 促进植物间促进作用产生的正反馈环路, 从而最终使附近景观尺度的植被覆盖和生态系统得以恢复。
除小节3中的案例外, 近年来研究发现可能发生稳态转换的生态系统还包括潮间带盐沼与滩涂湿地生态系统、珊瑚礁生态系统、海草床生态系统、海藻森林生态系统、寒区泥炭沼泽生态系统等。此外, 生态-进化过程、生态网络等系统中也可能存在多稳态和突变点。多稳态概念和框架还应用于气候环境、社会经济、生物医学等多种复杂系统中, 广泛用于研究古代文明的崩溃、人类情感交流、疾病发作、衰老与健康、可持续农业等问题。
生态系统多稳态和突变不断衍生出诸多的新兴研究热点。除了应用系统拓展、具体反馈机制, 以及早期预警信号的发现和适用性研究等方面吸引了大量关注, 近年来还与生态学中的诸多重要概念产生联系。例如, 基于复杂网络分析的研究表明, 具有突变点的不同系统之间可能存在遥相关而发生级联反应, 某些系统发生突变可能产生“多米诺”效应进而触发其他系统的突变(Rocha et al, 2018)。最近研究发现系统大小、连通性、空间相互作用、自组织过程、尺度、协同效应等一系列生态特征与过程可能与多稳态的形成机理及稳态转换的行为模式密切相关(Liu et al, 2014; Staal et al, 2016, 2018a; Cooper et al, 2020); 研究者也开始关注多稳态和稳态转换在深入理解生物多样性形成机制、生态系统功能维持等生态学核心问题中的作用(Fukami & Nakajima, 2011; Berdugo et al, 2017)。
5 讨论
英文文献中与稳态转换相关的名词包括“critical transition” “catastrophic shift” “regime shift” “abrupt shift” “tipping point”等。这些概念的内涵存在细微的差别, 例如“critical transition”和“catastrophic shift”通常明确指向以分岔行为为本质的动力学特征, 而后三者概念的界定相对较为宽泛, 描述的状态突变未必与分岔行为有关。由于相关术语目前在中文文献中尚未形成统一译法, 使用相关概念时预先进行内涵界定可能有助于避免混淆。
“tipping point” (中文翻译包括“临界点” “引爆点”等)在近年来文献中尤为常见。2000年出版的Malcom Gladwell的畅销书The Tipping Point极大促进了“tipping point”概念的应用。在科学研究中, “tipping point”可以定义为环境条件或系统状态的微小变化所触发的系统剧变(van Nes et al, 2016), 其内涵包含但不局限于分岔行为, 因此具有相对更为广泛的应用范围。鉴于此, 在更好的中文翻译出现之前, 本文暂将其翻译为“突变点”。突变点的具体位置可能存在不同看法。图1B山谷中的小球概念模型中, 山脊位置似乎可以从直观上理解为突变点。但如前所述, 山脊位置对应于不稳定平衡点, 在现实生态系统中出现概率极低。在图1中系统趋近分岔点F1或F2时, 生态系统状态都可以发生突变, 因此将分岔点F1或F2定义为突变点具有更为明确的理论基础(Scheffer et al, 2015a)。
多稳态理论的应用范围在近年来逐渐增加。但如前所述, 其理论基础和涉及的机理与分岔行为和正反馈具有紧密的联系, 在将多稳态概念应用到特定系统时, 应首先考虑是否存在较为明确的证据(或有根据的猜测), 并考虑其中可能存在的具体正反馈机制。尤其需要慎重考虑生态系统在时间或空间上的状态剧变是否由环境梯度或者其他机制导致, 在缺乏(小节2中所述的)多个相关证据时, 仅以生态系统状态的时空不连续特征为基础往往不足以认定系统存在多稳态。
此外, 图1B山谷中的小球概念模型由于其易于理解而得到广泛应用, 但对该概念模型的过度直观解释可能产生误解。例如, 当系统(小球)受到扰动时偏离稳态而后恢复的过程, 不能将真实世界中的小球因具有惯性而反复震荡这种现象进行简单类比(因为动力系统在状态空间中不具有惯性)来解释系统的波动。此外, 直观上山谷(吸引域)是静态的, 但实际上现实生态系统往往受到多种环境变化和扰动的影响, 吸引域也在不断变化(从直观上可以理解为山谷本身在不停地震动颠簸)。虽然目前稳定性景观的计算方法不断发展, 例如准势能(quasi-potential)分析方法(Nolting & Abbott, 2016), 但吸引域的重建一般是在一维或二维变量上, 多维变量上的吸引域刻画还缺少成熟的技术方法。总而言之, 多稳态理论目前主要基于鞍结点分岔的数学理论, 其相对简单的理论框架在应对复杂现实系统方面还存在诸多问题有待深入研究。
大量的动力系统理论研究已经揭示出许多非线性行为的数学基础以及分岔行为发生之前的系统行为变化(可以作为潜在的早期预警信号)。但对于真实世界中的生态系统而言, 在理论与实证研究之间建立坚实的联结往往需要大量的控制实验与长期观测。提升对生态系统突变(以及其他非线性行为)的预测能力仍然是当前生态学研究的重点和难点问题(Mouquet et al, 2015)。
6 结语
生态系统的多稳态和突变研究方兴未艾, 在我国生态学以及其他复杂系统(如气候系统、社会系统、微观生物系统等)研究中受到越来越多的关注。受作者知识水平所限, 本文对相关重要前沿进展的综述难免存在疏漏。仅希望本文能有助于读者开展多稳态理论与实证之间联结的相关研究和应用。
在多稳态理论的应用研究中, 特别需要关注该理论的前提假设、现实证据和模式预测。由于现实生态系统往往具有极高的复杂性并且受到研究时空尺度所限, 开展严格的控制实验通常较为困难, 对于特定生态系统而言, 推测其是否存在多稳态应避免仅仅从单一现象(如系统状态变量的变化轨迹的不连续性)存在与否而进行判断。条件满足时, 尽可能从多种证据出发, 综合借助于多种观测、实验和理论模型途径, 特别是简单理论模型(simple model)在理解现实世界中的生态现象和生态过程中往往可以发挥重要作用。当前, 在我国生态系统的多稳态和突变(以及其他生态系统的复杂非线性行为)研究中, 对于理论分析(模型)与现实系统的观测和实验的结合也日趋重视。多稳态的概念和理论基础的澄清有助于深入开展现实生态系统的突变现象和机理研究。
参考文献
Alternative tree-cover states of the boreal ecosystem: A conceptual model
Alternative stable states in ecology
Plant spatial patterns identify alternative ecosystem multifunctionality states in global drylands
Do alternate stable community states exist in the Gulf of Maine rocky intertidal zone?
Early warnings of regime shifts: A whole-ecosystem experiment
Rising variability, not slowing down, as a leading indicator of a stochastically driven abrupt transition in a dryland ecosystem
Regime shifts occur disproportionately faster in larger ecosystems
DOI:10.1038/s41467-020-15029-x
URL
PMID:32157098
[本文引用: 1]

Regime shifts can abruptly affect hydrological, climatic and terrestrial systems, leading to degraded ecosystems and impoverished societies. While the frequency of regime shifts is predicted to increase, the fundamental relationships between the spatial-temporal scales of shifts and their underlying mechanisms are poorly understood. Here we analyse empirical data from terrestrial (n = 4), marine (n = 25) and freshwater (n = 13) environments and show positive sub-linear empirical relationships between the size and shift duration of systems. Each additional unit area of an ecosystem provides an increasingly smaller unit of time taken for that system to collapse, meaning that large systems tend to shift more slowly than small systems but disproportionately faster. We substantiate these findings with five computational models that reveal the importance of system structure in controlling shift duration. The findings imply that shifts in Earth ecosystems occur over 'human' timescales of years and decades, meaning the collapse of large vulnerable ecosystems, such as the Amazon rainforest and Caribbean coral reefs, may take only a few decades once triggered.
On soil moisture-vegetation feedbacks and their possible effects on the dynamics of dryland ecosystems
Slower recovery in space before collapse of connected populations
DOI:10.1038/nature12071
URL
PMID:23575630
[本文引用: 1]
Slower recovery from perturbations near a tipping point and its indirect signatures in fluctuation patterns have been suggested to foreshadow catastrophes in a wide variety of systems. Recent studies of populations in the field and in the laboratory have used time-series data to confirm some of the theoretically predicted early warning indicators, such as an increase in recovery time or in the size and timescale of fluctuations. However, the predictive power of temporal warning signals is limited by the demand for long-term observations. Large-scale spatial data are more accessible, but the performance of warning signals in spatially extended systems needs to be examined empirically. Here we use spatially extended yeast populations, an experimental system with a fold bifurcation (tipping point), to evaluate early warning signals based on spatio-temporal fluctuations and to identify a novel spatial warning indicator. We found that two leading indicators based on fluctuations increased before collapse of connected populations; however, the magnitudes of the increases were smaller than those observed in isolated populations, possibly because local variation is reduced by dispersal. Furthermore, we propose a generic indicator based on deterministic spatial patterns, which we call 'recovery length'. As the spatial counterpart of recovery time, recovery length is the distance necessary for connected populations to recover from spatial perturbations. In our experiments, recovery length increased substantially before population collapse, suggesting that the spatial scale of recovery can provide a superior warning signal before tipping points in spatially extended systems.
Generic indicators for loss of resilience before a tipping point leading to population collapse
Resilience indicators: Prospects and limitations for early warnings of regime shifts
Fire drives functional thresholds on the savanna-forest transition
Disturbance maintains alternative biome states
Positive Feedback in Natural Systems. Springer-Verlag, Berlin
Early warning signals of extinction in deteriorating environments
DOI:10.1038/nature09389
URL
PMID:20827269
[本文引用: 1]
During the decline to extinction, animal populations may present dynamical phenomena not exhibited by robust populations. Some of these phenomena, such as the scaling of demographic variance, are related to small size whereas others result from density-dependent nonlinearities. Although understanding the causes of population extinction has been a central problem in theoretical biology for decades, the ability to anticipate extinction has remained elusive. Here we argue that the causes of a population's decline are central to the predictability of its extinction. Specifically, environmental degradation may cause a tipping point in population dynamics, corresponding to a bifurcation in the underlying population growth equations, beyond which decline to extinction is almost certain. In such cases, imminent extinction will be signalled by critical slowing down (CSD). We conducted an experiment with replicate laboratory populations of Daphnia magna to test this hypothesis. We show that populations crossing a transcritical bifurcation, experimentally induced by the controlled decline in environmental conditions, show statistical signatures of CSD after the onset of environmental deterioration and before the critical transition. Populations in constant environments did not have these patterns. Four statistical indicators all showed evidence of the approaching bifurcation as early as 110 days ( approximately 8 generations) before the transition occurred. Two composite indices improved predictability, and comparative analysis showed that early warning signals based solely on observations in deteriorating environments without reference populations for standardization were hampered by the presence of transient dynamics before the onset of deterioration, pointing to the importance of reliable baseline data before environmental deterioration begins. The universality of bifurcations in models of population dynamics suggests that this phenomenon should be general.
Review on alternative stable states in ecosystems
生态系统多稳态研究进展
Repeated fires trap Amazonian blackwater floodplains in an open vegetation state
Floodplains as an Achilles’ heel of Amazonian forest resilience
Historical contingency in community assembly: Integrating niches, species pools, and priority effects
Community assembly: Alternative stable states or alternative transient states?
More than the sum of its parts: Self-organized patterns and emergent properties in ecosystems
整体大于部分之和: 生态自组织斑图及其涌现属性
Rare, intense, big fires dominate the global tropics under drier conditions
Global resilience of tropical forest and savanna to critical transitions
DOI:10.1126/science.1210657
URL
PMID:21998390
[本文引用: 2]
It has been suggested that tropical forest and savanna could represent alternative stable states, implying critical transitions at tipping points in response to altered climate or other drivers. So far, evidence for this idea has remained elusive, and integrated climate models assume smooth vegetation responses. We analyzed data on the distribution of tree cover in Africa, Australia, and South America to reveal strong evidence for the existence of three distinct attractors: forest, savanna, and a treeless state. Empirical reconstruction of the basins of attraction indicates that the resilience of the states varies in a universal way with precipitation. These results allow the identification of regions where forest or savanna may most easily tip into an alternative state, and they pave the way to a new generation of coupled climate models.
El Niño as a window of opportunity for the restoration of degraded arid ecosystems
Strong facilitation in mild environments: The stress gradient hypothesis revisited
The interplay of facilitation and competition in plant communities
The effect of biomanipulation on phosphorus exchange between sediment and water in shallow, tropical Huizhou West Lake, China
Biomanipulation as a restoration tool to combat eutrophication: Recent advances and future challenges
Frost maintains forests and grasslands as alternate states in a montane tropical forest-grassland mosaic; but alien tree invasion and warming can disrupt this balance
Early warning signals also precede non-catastrophic transitions
DOI:10.1111/more.2013.122.issue-5 URL [本文引用: 2]
When can positive interactions cause alternative stable states in ecosystems?
Alternative stable states in coastal intertidal wetland ecosystems of Yangtze estuary, China
长江口滨海湿地潮间带生态系统的多稳态特征
Long-term ecosystem effects of sand-binding vegetation in the Tengger Desert, northern China
Survey on threshold detection methods of regime shift in shallow lake ecosystem
浅水湖泊生态系统稳态转换的阈值判定方法
Superorganisms or loose collections of species? A unifying theory of community patterns along environmental gradients
DOI:10.1111/ele.13289
URL
PMID:31134748
[本文引用: 2]
The question whether communities should be viewed as superorganisms or loose collections of individual species has been the subject of a long-standing debate in ecology. Each view implies different spatiotemporal community patterns. Along spatial environmental gradients, the organismic view predicts that species turnover is discontinuous, with sharp boundaries between communities, while the individualistic view predicts gradual changes in species composition. Using a spatially explicit multispecies competition model, we show that organismic and individualistic forms of community organisation are two limiting cases along a continuum of outcomes. A high variance of competition strength leads to the emergence of organism-like communities due to the presence of alternative stable states, while weak and uniform interactions induce gradual changes in species composition. Dispersal can play a confounding role in these patterns. Our work highlights the critical importance of considering species interactions to understand and predict the responses of species and communities to environmental changes.
Pattern formation at multiple spatial scales drives the resilience of mussel bed ecosystems
DOI:10.1038/ncomms6234
URL
PMID:25335554
[本文引用: 1]
Self-organized complexity at multiple spatial scales is a distinctive characteristic of biological systems. Yet, little is known about how different self-organizing processes operating at different spatial scales interact to determine ecosystem functioning. Here we show that the interplay between self-organizing processes at individual and ecosystem level is a key determinant of the functioning and resilience of mussel beds. In mussel beds, self-organization generates spatial patterns at two characteristic spatial scales: small-scale net-shaped patterns due to behavioural aggregation of individuals, and large-scale banded patterns due to the interplay of between-mussel facilitation and resource depletion. Model analysis reveals that the interaction between these behavioural and ecosystem-level mechanisms increases mussel bed resilience, enables persistence under deteriorating conditions and makes them less prone to catastrophic collapse. Our analysis highlights that interactions between different forms of self-organization at multiple spatial scales may enhance the intrinsic ability of ecosystems to withstand both natural and human-induced disturbances.
Potential analysis reveals changing number of climate states during the last 60 kyr
Review: Predictive ecology in a changing world
Balls, cups, and quasi-potentials: Quantifying stability in stochastic systems
DOI:10.1890/15-1047.1
URL
PMID:28792592
[本文引用: 1]
When a system has more than one stable state, how can the stability of these states be compared? This deceptively simple question has important consequences for ecosystems, because systems with alternative stable states can undergo dramatic regime shifts. The probability, frequency, duration, and dynamics of these shifts will all depend on the relative stability of the stable states. Unfortunately, the concept of
Stability of grazing systems: An application of predator-prey graphs
DOI:10.2307/2258730 URL [本文引用: 1]
Macroinvertebrate responses to regime shifts caused by eutrophication in subtropical shallow lakes
Divergent succession and implications for alternative states on rocky intertidal shores
Self-organized patchiness and catastrophic shifts in ecosystems
DOI:10.1126/science.1101867
URL
PMID:15448261
[本文引用: 2]
Unexpected sudden catastrophic shifts may occur in ecosystems, with concomitant losses or gains of ecological and economic resources. Such shifts have been theoretically attributed to positive feedback and bistability of ecosystem states. However, verifications and predictive power with respect to catastrophic responses to a changing environment are lacking for spatially extensive ecosystems. This situation impedes management and recovery strategies for such ecosystems. Here, we review recent studies on various ecosystems that link self-organized patchiness to catastrophic shifts between ecosystem states.
Cascading regime shifts within and across scales
Early-warning signals for critical transitions
DOI:10.1038/nature08227
URL
PMID:19727193
Complex dynamical systems, ranging from ecosystems to financial markets and the climate, can have tipping points at which a sudden shift to a contrasting dynamical regime may occur. Although predicting such critical points before they are reached is extremely difficult, work in different scientific fields is now suggesting the existence of generic early-warning signals that may indicate for a wide class of systems if a critical threshold is approaching.
Catastrophic regime shifts in ecosystems: Linking theory to observation
Generic indicators of ecological resilience: Inferring the chance of a critical transition
Anticipating critical transitions
DOI:10.1126/science.1225244
URL
PMID:23087241
[本文引用: 3]
Tipping points in complex systems may imply risks of unwanted collapse, but also opportunities for positive change. Our capacity to navigate such risks and opportunities can be boosted by combining emerging insights from two unconnected fields of research. One line of work is revealing fundamental architectural features that may cause ecological networks, financial markets, and other complex systems to have tipping points. Another field of research is uncovering generic empirical indicators of the proximity to such critical thresholds. Although sudden shifts in complex systems will inevitably continue to surprise us, work at the crossroads of these emerging fields offers new approaches for anticipating critical transitions.
Catastrophic shifts in ecosystems
Thresholds for boreal biome transitions
Alternative equilibria in shallow lakes
Shallow lakes theory revisited: Various alternative regimes driven by climate, nutrients, depth and lake size
Why trees and shrubs but rarely trubs?
DOI:10.1016/j.tree.2014.06.001
URL
PMID:24951396
[本文引用: 1]
An analysis of the maximum height of woody plant species across the globe reveals that an intermediate size is remarkably rare. We speculate that this may be due to intrinsic suboptimality or to ecosystem bistability with open landscapes favouring shrubs, and closed canopies propelling trees to excessive tallness.
The mystery of missing trubs revisited: A response to McGlone et al and Qian and Ricklefs
DOI:10.1016/j.tree.2014.11.001 URL PMID:25468358 [本文引用: 1]
A global climate niche for giant trees
DOI:10.1111/gcb.14167
URL
PMID:29658194
[本文引用: 2]
Rainforests are among the most charismatic as well as the most endangered ecosystems of the world. However, although the effects of climate change on tropical forests resilience is a focus of intense research, the conditions for their equally impressive temperate counterparts remain poorly understood, and it remains unclear whether tropical and temperate rainforests have fundamental similarities or not. Here we use new global data from high precision laser altimetry equipment on satellites to reveal for the first time that across climate zones 'giant forests' are a distinct and universal phenomenon, reflected in a separate mode of canopy height (~40 m) worldwide. Occurrence of these giant forests (cutoff height > 25 m) is negatively correlated with variability in rainfall and temperature. We also demonstrate that their distribution is sharply limited to situations with a mean annual precipitation above a threshold of 1,500 mm that is surprisingly universal across tropical and temperate climates. The total area with such precipitation levels is projected to increase by ~4 million km(2) globally. Our results thus imply that strategic management could in principle facilitate the expansion of giant forests, securing critically endangered biodiversity as well as carbon storage in selected regions.
Direct experimental evidence for alternative stable states: A review
Sharp ecotones spark sharp ideas: comment on “Structural, physiognomic and above-ground biomass variation in savanna-forest transition zones on three continents-how different are co-occurring savanna and forest formations?” by Veenendaal et al (2015)
Feedback between drought and deforestation in the Amazon
Bistability, spatial interaction, and the distribution of tropical forests and savannas
Forest-rainfall cascades buffer against drought across the Amazon
Resilience of tropical tree cover: The roles of climate, fire, and herbivory
The global extent and determinants of savanna and forest as alternative biome states
The Anthropocene: are humans now overwhelming the great forces of nature
DOI:10.1579/0044-7447(2007)36[614:taahno]2.0.co;2
URL
PMID:18240674
[本文引用: 1]
We explore the development of the Anthropocene, the current epoch in which humans and our societies have become a global geophysical force. The Anthropocene began around 1800 with the onset of industrialization, the central feature of which was the enormous expansion in the use of fossil fuels. We use atmospheric carbon dioxide concentration as a single, simple indicator to track the progression of the Anthropocene. From a preindustrial value of 270-275 ppm, atmospheric carbon dioxide had risen to about 310 ppm by 1950. Since then the human enterprise has experienced a remarkable explosion, the Great Acceleration, with significant consequences for Earth System functioning. Atmospheric CO2 concentration has risen from 310 to 380 ppm since 1950, with about half of the total rise since the preindustrial era occurring in just the last 30 years. The Great Acceleration is reaching criticality. Whatever unfolds, the next few decades will surely be a tipping point in the evolution of the Anthropocene.
Morphological traits of submerged macrophytes reveal specific positive feedbacks to water clarity in freshwater ecosystems
Stoichiometric mechanisms of regime shifts in freshwater ecosystem
Alternative states and positive feedbacks in restoration ecology
DOI:10.1016/j.tree.2003.10.005
URL
PMID:16701225
[本文引用: 1]
There is increasing interest in developing better predictive tools and a broader conceptual framework to guide the restoration of degraded land. Traditionally, restoration efforts have focused on re-establishing historical disturbance regimes or abiotic conditions, relying on successional processes to guide the recovery of biotic communities. However, strong feedbacks between biotic factors and the physical environment can alter the efficacy of these successional-based management efforts. Recent experimental work indicates that some degraded systems are resilient to traditional restoration efforts owing to constraints such as changes in landscape connectivity and organization, loss of native species pools, shifts in species dominance, trophic interactions and/or invasion by exotics, and concomitant effects on biogeochemical processes. Models of alternative ecosystem states that incorporate system thresholds and feedbacks are now being applied to the dynamics of recovery in degraded systems and are suggesting ways in which restoration can identify, prioritize and address these constraints.
Identification of 100 fundamental ecological questions
Scale-dependent inhibition drives regular tussock spacing in a freshwater marsh
DOI:10.1086/508671
URL
PMID:17080356
[本文引用: 3]
Regular spatial patterning is common in nature, and various mechanisms of self-organization have been proposed to explain regular patterning. We report on regular spatial patterning in Carex stricta in a freshwater wetland and investigate the applicability of theoretical models that explain regular patterning based on inhibition, facilitation, or interaction between the two. Spectral analysis of aerial photographs revealed that tussocks were regularly spaced at an average distance of 60 cm. Photosynthetically active radiation varied significantly with distance from the tussock and was lowest at intermediate distance from the tussock center (15-40 cm). Using transplants to assay growth conditions, we found that C. stricta grew well in all distance classes with and without natural C. stricta biomass, except at intermediate distances when buried in C. stricta wrack. Our experimental results reveal that C. stricta inhibits its growth in a scale-dependent manner: inhibition was found to peak at intermediate distance from the tussock. We compared three alternative models to examine potential mechanisms driving regularity and found that, similar to our experimental results, scale-dependent inhibition provides the best explanation for the observed regular tussock spacing. Our study underlines the importance of scale-dependent feedback in the formation of regular spatial patterning in ecosystems.
Catastrophic vegetation shifts and soil degradation in terrestrial grazing systems
DOI:10.1016/s0169-5347(97)01133-6
URL
PMID:21238102
[本文引用: 1]
It has long been recognized that alternative vegetation states may occur in terrestrial grazing systems. This phenomenon may be of great importance as small environmental fluctuations may lead to relatively sudden and irreversible jumps between vegetation states. Early theoretical studies emphasized saturation of herbivore feeding to explain multiple stable states and catastrophic behaviour. Recent studies on semi-arid grasslands and arctic salt marshes, however, relate catastrophic events in these systems to plant-soil interactions.
Effects of fire and herbivory on the stability of savanna ecosystems
What do you mean, ‘tipping point’?
DOI:10.1016/j.tree.2016.09.011
URL
PMID:27793466
[本文引用: 1]
Over the past 10 years the use of the term 'tipping point' in the scientific literature has exploded. It was originally used loosely as a metaphor for the phenomenon that, beyond a certain threshold, runaway change propels a system to a new state. Although several specific mathematical definitions have since been proposed, we argue that these are too narrow and that it is better to retain the original definition.
Fire forbids fifty-fifty forest
Recovery rates reflect distance to a tipping point in a living system
DOI:10.1038/nature10723
URL
PMID:22198671
[本文引用: 1]
Tipping points, at which complex systems can shift abruptly from one state to another, are notoriously difficult to predict. Theory proposes that early warning signals may be based on the phenomenon that recovery rates from small perturbations should tend to zero when approaching a tipping point; however, evidence that this happens in living systems is lacking. Here we test such 'critical slowing down' using a microcosm in which photo-inhibition drives a cyanobacterial population to a classical tipping point when a critical light level is exceeded. We show that over a large range of conditions, recovery from small perturbations becomes slower as the system comes closer to the critical point. In addition, autocorrelation in the subtle fluctuations of the system's state rose towards the tipping point, supporting the idea that this metric can be used as an indirect indicator of slowing down. Although stochasticity prohibits prediction of the timing of critical transitions, our results suggest that indicators of slowing down may be used to rank complex systems on a broad scale from resilient to fragile.
Predictive Limnological Researches on Small- to Medium-sized Lakes Along the Mid-lower Yangtze River
长江中下游中小型湖泊预测湖沼学研究
Total phosphorus thresholds for regime shifts are nearly equal in subtropical and temperate shallow lakes with moderate depths and areas
Impacts of regime shift between phytoplankton and macrophyte on the microbial community structure and its carbon cycling in lakes
草藻型稳态转换对湖泊微生物结构及其碳循环功能的影响
Local facilitation may cause tipping points on a landscape level preceded by early-warning indicators
Remotely sensed canopy height reveals three pantropical ecosystem states
A changing number of alternative states in the boreal biome: Reproducibility risks of replacing remote sensing products
DOI:10.1371/journal.pone.0143014
URL
PMID:26571014
[本文引用: 1]
Publicly available remote sensing products have boosted science in many ways. The openness of these data sources suggests high reproducibility. However, as we show here, results may be specific to versions of the data products that can become unavailable as new versions are posted. We focus on remotely-sensed tree cover. Recent studies have used this public resource to detect multi-modality in tree cover in the tropical and boreal biomes. Such patterns suggest alternative stable states separated by critical tipping points. This has important implications for the potential response of these ecosystems to global climate change. For the boreal region, four distinct ecosystem states (i.e., treeless, sparse and dense woodland, and boreal forest) were previously identified by using the Collection 3 data of MODIS Vegetation Continuous Fields (VCF). Since then, the MODIS VCF product has been updated to Collection 5; and a Landsat VCF product of global tree cover at a fine spatial resolution of 30 meters has been developed. Here we compare these different remote-sensing products of tree cover to show that identification of alternative stable states in the boreal biome partly depends on the data source used. The updated MODIS data and the newer Landsat data consistently demonstrate three distinct modes around similar tree-cover values. Our analysis suggests that the boreal region has three modes: one sparsely vegetated state (treeless), one distinct 'savanna-like' state and one forest state, which could be alternative stable states. Our analysis illustrates that qualitative outcomes of studies may change fundamentally as new versions of remote sensing products are used. Scientific reproducibility thus requires that old versions remain publicly available.
Can we infer plant facilitation from remote sensing? A test across global drylands
DOI:10.1890/14-2358.1
URL
PMID:26552256
[本文引用: 1]
Facilitation is a major force shaping the structure and diversity of plant communities in terrestrial ecosystems. Detecting positive plant-plant interactions relies on the combination of field experimentation and the demonstration of spatial association between neighboring plants. This has often restricted the study of facilitation to particular sites, limiting the development of systematic assessments of facilitation over regional and global scales. Here we explore whether the frequency of plant spatial associations detected from high-resolution remotely sensed images can be used to infer plant facilitation at the community level in drylands around the globe. We correlated the information from remotely sensed images freely available through Google Earth with detailed field assessments, and used a simple individual-based model to generate patch-size distributions using different assumptions about the type and strength of plant-plant interactions. Most of the patterns found from the remotely sensed images were more right skewed than the patterns from the null model simulating a random distribution. This suggests that the plants in the studied drylands show stronger spatial clustering than expected by chance. We found that positive plant co-occurrence, as measured in the field, was significantly related to the skewness of vegetation patch-size distribution measured using Google Earth images. Our findings suggest that the relative frequency of facilitation may be inferred from spatial pattern signals measured from remotely sensed images, since facilitation often determines positive co-occurrence among neighboring plants. They pave the road for a systematic global assessment of the role of facilitation in terrestrial ecosystems.
Integrating underwater light condition and seed bank to indicate submersed macrophyte restoration zone: Lake Jinhu as a case
基于水下光照条件和种子库分布指示沉水植物恢复区: 以金湖为例
Remotely sensed canopy height reveals three pantropical ecosystem states: Reply
Temperate forest and open landscapes are distinct alternative states as reflected in canopy height and tree cover
DOI:10.1016/j.tree.2015.07.002 URL PMID:26209871 [本文引用: 1]
Critical transitions in Chinese dunes during the past 12,000 years
DOI:10.1126/sciadv.aay8020
URL
PMID:32133406
Dune systems can have alternative stable states that coexist under certain environmental conditions: a vegetated, stabilized state and a bare active state. This behavior implies the possibility of abrupt transitions from one state to another in response to gradual environmental change. Here, we synthesize stratigraphic records covering 12,000 years of dynamics of this system at 144 localities across three dune fields in northern China. We find side-by-side coexistence of active and stabilized states, and occasional sharp shifts in time between those contrasting states. Those shifts occur asynchronously despite the fact that the entire landscape has been subject to the same gradual changes in monsoon rainfall and other conditions. At larger scale, the spatial heterogeneity in dune dynamics averages out to produce relatively smooth change. However, our results do show different paths of recovery and collapse of vegetation at system-wide scales, implying that hysteretic behavior occurs in spatially extended systems.
Survey on theory and driving factors of regime shifts on lake ecosystems
湖泊生态系统稳态转换理论与驱动因子研究进展
The shaping role of self-organization: Linking vegetation patterning, plant traits and ecosystem functioning
Regime shifts revealed by paleoecological records in Lake Taibai’s ecosystem in the middle and lower Yangtze River Basin during the last century
古生态记录揭示的长江中下游太白湖生态系统稳态转换过程

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}