


生态网络刻画了生态系统中生物组分之间的相互作用。食物网是生态学中研究最早、最多的网络类型, 它表征了物种之间通过捕食而形成的复杂网络关系。食物网表明生态系统的营养结构和过程具有高度的复杂性, 这一复杂性是生态系统进行物质循环和能量流动的基础(Lindeman, 1942; Bascompte, 2009; Thompson et al, 2012)。阐明食物网结构与功能之间的关系, 既是生态学的基本理论问题, 也是预测全球变化背景下生态系统响应的重要依据(Tylianakis et al, 2008; Sentis et al, 2014)。
虽然食物网是生态学研究的一个经典议题, 但早期关于食物网结构和功能的研究往往是分离的。自Elton (1927)发表Animal Ecology一书以来, 群落生态学家关于食物网的拓扑结构、不同营养级的数量特征等开展了大量研究。随后, Lindeman (1942)关于湖泊生态系统营养动态的研究发表, 引发了生态系统生态学家对食物网能量流动、营养传递效率和生产力等的广泛研究。过去几十年来, 生态学家尝试将食物网结构与功能结合起来, 但这些研究大多关注简单网络模块, 比如两个或三个物种构成的食物链或竞争模块(Pimm, 1982; Loreau, 2010; McCann, 2012)。这些简单模块只是整个食物网的一个子集, 基于此类子集的研究结果并不一定能推广至复杂食物网(Wang & Brose, 2018)。为了理解自然生态系统整体的结构和功能, 需要开展复杂食物网研究(Seibold et al, 2018)。然而, 食物网的高度复杂性为研究工作带来了很大的挑战, 尤其是经验分析。因此, 为了理解复杂食物网的结构和功能及其应对外界干扰的稳定性, 生态学家常常借助理论模型方法进行研究(May, 1973; McCann, 2012; Moore & de Ruiter, 2012)。
本文对近期关于复杂食物网结构与功能的理论进展做了综述。首先介绍复杂食物网中网络结构、多样性和功能的度量指标, 接着围绕以下三个问题对近期进展做了概述(图1): 食物网结构如何影响物种多样性维持? 食物网结构与多样性如何影响生态系统功能? 全球变化如何改变食物网结构、多样性与功能? 本文所介绍的理论研究主要基于复杂食物网模型, 在Box 1中对此类模型的基本框架做了阐述。需要说明的是, 复杂性与稳定性关系是食物网研究中的一个经典和重要问题, 关于这一问题已有多篇中文综述(陈云峰等, 2014; 徐光华等, 2019), 因此本文不作特别介绍。
图1
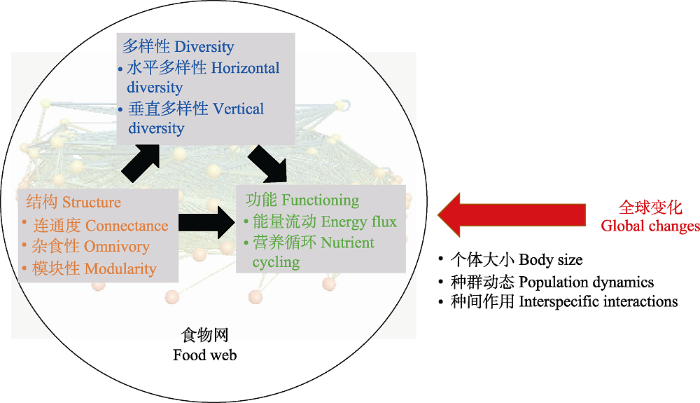
图1
复杂食物网的结构、多样性与功能间的关系。食物网结构可影响物种多样性和生态系统功能的维持, 多样性也对功能起重要作用。全球变化可通过改变个体大小、种群动态、种间作用等影响食物网结构、多样性与功能。背景食物网来自http://www.foodwebs.org/。
Fig.1
Relationships between the structure, diversity and functioning of complex food webs. Food web structure can affect species diversity and ecosystem functioning, and diversity also affects functioning. Global change can affect food webs through changing species body size, population dynamics, interspecific interaction, etc. The food web in the background was obtained from http://www.foodwebs.org/.
| Box 1 复杂食物网理论模型 |
| 食物网建模的核心是对网络结构与种群动态的理论刻画。网络结构决定了每个物种在生态系统中的位置, 即“谁吃谁” (Who eats who?)。种群动态刻画了任一物种如何受其捕食者和猎物的影响, 即“如何吃” (How does a predator eat its prey?)。这里概述食物网结构与动态模型的一般框架。在具体应用中, 针对研究问题可有不同的具体模型。比如在复杂性-稳定性研究中, 陈云峰等(2014)和徐光华等(2019)综述了多种常用的食物网模型。 |
| $\cdot$“谁吃谁”: 模拟食物网结构 |
| 生态学家提出了多种理论模型试图刻画食物网的网络结构。早期的理论模型包括随机模型(May, 1973)和级联模型(cascade model; Cohen & Newman, 1985)。随机模型假定生态系统内不同物种之间的捕食关系是随机发生的, 往往只作为一个零模型来使用。比如, May (1973)关于生态系统稳定性与复杂性关系的研究就使用了随机模型。级联模型考虑了物种间的非对称性, 假定物种可在某一性状轴上排序, 性状值高的物种可捕食性状值低的物种, 反之则不成立。该模型假定的性状轴可以是影响捕食关系的特定性状, 比如个体大小。然而有研究表明, 个体较大的物种并非取食所有比其小的物种, 而是有一个最优取食范围(Persson et al, 1998)。基于这一考虑, Williams和Martinez (2000)提出了生态位模型(niche model)。该模型在级联模型的基础上, 假定捕食者只捕食某一性状范围内的物种, 且允许自食和互相捕食。生态位模型为自然界食物网的多种结构指标给出了很好的解释, 因此成为食物网研究的一个经典模型。后续研究基于生态位模型发展了很多变种, 从不同角度对生态位模型做了扩展, 包括引入谱系关系(Cattin et al, 2004)、多维生态位(Allesina et al, 2008)、个体大小限制(Schneider et al, 2016)等。需要注意的是, 物种分布模型中同样有“生态位模型”, 用以刻画物种对环境因子的响应方式, 与此处模拟食物网结构的生态位模型不同。 |
| $\cdot$“如何吃”: 模拟食物网动态与功能 |
| 对食物网中的任一物种而言, 种群增长主要取决于其获取资源的速率, 种群下降主要取决于其被捕食的速率以及新陈代谢和自然死亡。因此, 种群动态模拟的关键是给出物种获取资源或捕食的速率。通常地, 这一速率由功能响应(functional response)函数刻画(Holling, 1959): |
| $F(N)=\frac{a{{N}^{q}}}{1+ah{{N}^{q}}}$ (B1) |
| 其中, N表示猎物密度, F(N)表示单个捕食者的捕食速率, a表示捕食者的攻击速率(attack rate), h表示消化或处理一只猎物所需的时间(handling time)。q是一个调控密度依赖形式的参数。当q = 0时, 捕食速率F(N)是与猎物密度N无关的常数, 称为“I型”功能响应; q = 1时, F(N)在密度N较低时随N线性增长, 随后增长速率减慢, 最终达到饱和水平, 称为“II型”功能响应; q = 2时, F(N)在密度N较低时随N呈二次函数增长, 随后增长减慢, 最终达到饱和水平, 称为“III型”功能响应。很多研究者通过实验分析了猎物密度和捕食速率之间的关系, 发现很多实验系统都服从“II型”功能响应(Rall et al, 2012)。但近期也有实验表明, 功能响应的类型受物种属性的影响: 当捕食者与猎物个体大小接近时, 其捕食过程服从“II型”功能响应; 而当捕食者比猎物大很多时, 服从“III型”功能响应(Kalinkat et al, 2013)。虽然以上三种功能响应函数最为常用, 但生态学家也提出了很多其他模型来刻画捕食速率和猎物密度的关系(Turchin, 2003; Yan & Zhang, 2014)。 |
| 基于功能响应函数, 食物网中任一植物种i(Pi)和动物种j(Aj)的种群大小的时间动态由以下方程决定(Schneider et al, 2016): |
| $\frac{d{{P}_{i}}}{dt}={{r}_{i}}{{G}_{i}}{{P}_{i}}-\underset{k}{\mathop \sum }\,{{A}_{k}}{{F}_{ki}}-{{x}_{i}}{{P}_{i}}$ (B2) |
| $\frac{d{{A}_{j}}}{dt}={{e}_{1}}{{A}_{j}}\cdot \sum\limits_{i:植物资源}{{{F}_{ji}}+{{e}_{2}}{{A}_{j}}}\cdot \sum\limits_{j:动物资源}{{{F}_{jk}}-}\underset{l}{\mathop \sum }\,{{A}_{l}}{{F}_{lj}}-{{x}_{j}}{{A}_{j}}~~$. (B3) |
| 其中, ri 表示植物种i的最大生长速率, Gi刻画了营养元素对植物种i生长的限制(见Schneider et al, 2016), xi表示个体代谢速率; e1和e2分别是取食植物和动物时的能量利用效率。Fji表示功能响应函数, 刻画了捕食者j对其猎物种i的取食速率。但不同于公式B1, 由于捕食者j有多种猎物, 因此功能响应函数扩展为下式(Schneider et al, 2016): |
| ${{F}_{ji}}=\frac{{{\omega }_{ji}}{{a}_{ji}}N_{i}^{q}}{1+\mathop{\sum }_{k}{{\omega }_{jk}}{{a}_{jk}}{{h}_{jk}}N_{k}^{q}}$ (B4) |
| 其中Ni表示第i个猎物种的生物量, ajk和hjk表示捕食者j对猎物k的攻击速率和处理时间, ωjk表示捕食者j对猎物k的取食偏好。类似地, q表示取食速率对猎物密度的依赖关系为II型(q = 1)或III型(q = 2)功能响应。 |
| 为了模拟食物网功能, 后续研究通过引入物种个体大小, 将基于种群的模型拓展到基于生物量的模型, 其中关注变量不再是“种群变化”, 而是“生物量变化” (Yodzis & Innes, 1992; Brose et al, 2006)。这一拓展将捕食过程直接描述为能量流动过程, 从而构建了食物网种群动态与生态系统功能之间的联系。此后基于大量食物网的数据分析表明, 个体大小对食物网结构(捕食关系)和动态(代谢速率、捕食过程)均有重要影响(Brose et al, 2006; Rall et al, 2012)。在此基础上, Schneider等(2016)发展了一个新的基于个体大小的参数化食物网模型, 其中食物网结构和动态均由个体大小决定。该模型整合了物种性状、代谢理论、能量动态等, 为研究食物网结构和功能提供了一个新的理论工具(Wang & Brose, 2018)。 |
1 食物网结构、多样性和功能的度量
1.1 食物网结构指标
生态学家提出了大量指标来刻画食物网的结构(Gauzens et al, 2013; 孙书存, 2017)。在整个食物网水平上, 三个量化生态系统复杂性的常用指标是: 总物种数(S, 即食物网中所有动物和植物的物种数)、总连接数(L, 即食物网中具有捕食关系的所有连接数)和连通度(connectance, L/S2)。此外, 模块性(modularity)度量了食物网的分室特征(compartment), 具体指食物网内由不同物种集合形成的子模块, 模块内物种有较紧密的交互关系, 模块间的交互关系相对较少或较弱(Yodzis, 1982; Krause et al, 2003)。
在物种水平上, 可以定义物种的连接度(degree, 即与该物种具有捕食关系的所有捕食者和猎物的物种数)、广食性(generality, 即该物种的所有猎物种数)、脆弱性(vulnerability, 即该物种的所有捕食者种数)等。对于食物网中的物种来说, 一个核心概念是物种的营养级(trophic level)。营养级刻画了物种在生态系统中的营养位置, 通常定义为从非生物资源到该物种的能量传递路径的长度。如果该物种取食多种资源, 那么可根据其与非生物资源的不同传递路径定义其营养级: 最短链营养级、最长链营养级、平均营养级(Williams & Martinez, 2004)。当所有路径长度都相同时, 三种营养级的定义是一致的; 反之, 则意味着不同传递路径长度不同, 三种营养级的计算结果也不同。产生这种差异的原因即物种的杂食性(omnivory), 它指的是食物网中的某些物种可以取食位于两个(或多个)不同营养级的物种(Pimm, 1982)。狭义上的杂食性指某种动物可取食植物和动物, 而食物网研究中一般采用广义的杂食性定义, 即可以取食位于不同营养级的两物种。研究发现, 自然界食物网中存在广泛的杂食性(Thompson et al, 2007)。与杂食性相对应, 有学者提出了营养一致性(trophic coherence)指标(Johnson et al, 2014)。该指标度量了捕食者与猎物的营养级差值的变异性, 因此杂食性越高, 营养一致性越低。
杂食性和模块性是食物网结构复杂性的两个重要方面。杂食性导致不同营养级之间的差别变得模糊, 因而增加了食物网垂直结构的复杂性。模块性不仅增大了食物网内种间相互作用的异质性, 还增加了食物网水平和垂直结构的复杂性。如果没有杂食性和模块性, 食物网很大程度上可简化为食物链(即每个营养级由若干营养功能相似的物种组成)。因此, 阐明食物网中的杂食性和模块性特征, 是理解食物网结构复杂性的一个关键。
1.2 食物网的水平和垂直多样性
虽然物种多样性也可作为食物网的结构指标之一, 但由于很多研究专门探讨食物网中多样性的维持机制及其生态功能, 因此本文将多样性作为一类独立指标进行阐述。在食物网中, 多样性可由不同指标、不同维度来刻画。指标上, 可基于物种数和连接度来计算。特别地, 生态系统中的所有物种的总数可以表征总的物种多样性, 所有连接的总数可以表征总的种间关系多样性。维度上, 食物网一般可由二维平面图表示, 垂直高度表示物种的营养级, 即处于同一高度的物种属于同一营养级, 因而可从水平和垂直两个维度刻画食物网的多样性(图2a)。水平多样性(horizontal diversity)刻画了某一营养级内的物种多样性, 垂直多样性(vertical diversity)则刻画了跨营养级的结构复杂性。后者可通过营养级的数目来度量, 比如食物链长度、复杂食物网的平均营养级或最高营养级(Duffy et al, 2007; Wang & Brose, 2018)。
图2
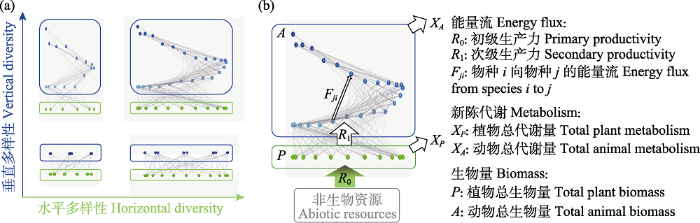
图2
食物网中的生物多样性(a)和功能(b)。在食物网中, 生物多样性可通过水平多样性(如同一营养级内的物种数)和垂直多样性(如食物网的最高营养级)度量, 功能可通过营养级或物种之间的能量流、植物或动物的总代谢量和生物量刻画。其中, 非生物资源与植物群落(绿色框)之间的能量流表征了初级生产力, 植物与动物群落(蓝色框)之间的能量流表征了次级生产力。修改自Wang & Brose (2018)和Schneider等(2016)。
Fig. 2
Food web diversity (a) and functioning (b). Biodiversity in food webs can be measured by both horizontal diversity (e.g. species richness within trophic levels) and vertical diversity (e.g. the maximum trophic level). Functioning can be measured by energy fluxes between two trophic levels or two species, total plant or animal metabolism, and total plant or animal biomass. Specifically, the energy flux from abiotic resource to plant communities (green box) represents the primary productivity, and that from plant communities to animal communities (blue box) represents the secondary productivity. Modified from Wang & Brose (2018) and Schneider et al (2016).
此外, Duffy等(2007)提出也可基于连接数来定义水平和垂直多样性, 即水平生态位幅度(horizontal niche breadth)和垂直生态位幅度(vertical niche breadth)。具体地, 水平生态位幅度由捕食者的广食性刻画, 即捕食者的总猎物种数; 垂直生态位幅度由捕食者的杂食性刻画, 即其猎物所分布的营养级范围。但在本文中, 水平和垂直多样性特指基于物种数的指标, 即分别指营养级内物种数和营养级的数目, 而将广食性和杂食性作为网络结构指标。
1.3 食物网的功能
食物网的主要生态功能是能量传递和物质循环。Elton (1927)提出了数量金字塔(numerical pyramid)的概念, 表征不同营养级间的种群大小分布。这一概念后来被发展为能量金字塔(energy pyramid), 用来量化不同营养级之间的能量传递过程(Lindeman, 1942; Trebilco et al, 2013)。其中, 能量金字塔的最底层为初级生产力, 表征了初级生产者从非生物环境中固定的能量; 第二层为次级生产力, 量化了初级生产者向初级消费者传递的能量(图2b)。近期研究对能量金字塔的概念做了扩展, 通过计算任意两物种之间的能量流(energy flux)来刻画食物网局部的功能特征, 从而对食物网能量动态做出全面量化(Barnes et al, 2018) (图2b)。具体地, 物种之间的能量流可通过能量守恒法则计算得出, 即物种的能量输入等于能量输出(de Ruiter et al, 1994)。由于需要对每一物种计算其能量平衡, 因此该算法有较大的计算复杂度。最近Gauzens等(2019)发展了一个矩阵算法, 并提供了R软件包fluxweb, 只要给出食物网中每一物种的代谢速率和取食比例, 则可推断整个网络的能量流。其中, 代谢速率可由物种个体大小推算得到(Brown et al, 2004; Barnes et al, 2018)。除了能量流, 近期研究也常用植物和动物的总生物量或总代谢量作为食物网的功能指标(Schneider et al, 2016; Wang S et al, 2019; Maureaud et al, 2020)。
在物质循环方面, 解析碳、氮、磷等元素在生态系统和大气之间的循环过程是生态系统研究的核心内容(傅声雷等, 2019)。但近期食物网理论研究大多只关注能量流动, 对物质循环过程的理论探讨仍较少, 因此本文主要关注食物网结构和多样性对能量过程的作用。整合食物网与物质循环将是未来研究的一个重要方向(详见5.2节)。
2 食物网结构如何影响物种多样性?
经典的竞争理论预测, 共存的物种数不会超过限制资源的种类(Tilman, 1982)。但如果竞争同一资源的两物种有一个共同的捕食者, 且二者在资源利用和抵御捕食上存在权衡关系, 则可实现共存(Holt et al, 1994)。这是由于两个竞争物种不仅在资源获取上形成“利用性竞争” (exploitative competition), 还由于捕食者的存在而形成“似然竞争” (apparent competition), 后者为两个物种构建了新的生态位, 从而促进物种共存。Chesson和Kuang (2008)基于现代物种共存理论进一步揭示, 物种在资源利用或抵御捕食方面的差异性可通过类似机制促进共存, 因此利用性竞争和似然竞争在影响物种共存上具有一定的对称性。
在前期基于竞争或捕食研究的基础上, 近期的理论研究探讨了复杂食物网内的物种多样性维持。Brose (2008)利用复杂食物网研究了动物群落对植物物种共存的影响, 发现即使只有一种非生物营养资源, 食物网也可以维持几十种植物的稳定共存(也见Wang & Brose, 2018)。其原因是, 动物群落形成的复杂食物网改变了植物群落的生物环境, 极大拓展了植物群落的生态位空间, 由于不同植物与食物网中的不同动物形成营养关系, 这种非对称的“营养生态位” (trophic niche)促进了植物种之间的共存。这一理论预测得到了近期实验的支持。如Wang L等(2019)在我国内蒙古草地开展的放牧实验发现, 混合放牧有利于维持更高的草地物种多样性。
对食物网整体而言, 网络结构对多样性维持有重要影响, 这里主要关注杂食性和模块性结构。Stouffer和Bascompte (2010)研究了复杂食物网中四种不同的三物种组件(比如食物链、竞争、似然竞争、杂食性)对多样性维持的作用。结果表明, 食物网中的杂食性组件越多, 物种多样性越高, 因而杂食性有利于复杂食物网中的多样性维持。Wang S等(2019)考虑了一种特定的杂食性结构, 即集团内捕食(intraguild predation), 通过模拟去除食物网中的集团内捕食, 发现该结构有利于维持更高的物种多样性。该研究结合五物种的简单网络和复杂食物网揭示了其作用机制: 集团内捕食促使能量在营养级内部停留, 减缓了能量从低营养级到高营养级的传递, 从而可降低高营养级物种的下行调控, 最终增强了整个食物网的种群稳定性和多样性维持。此外, 模块性使得食物网中不同物种的相互作用局部化, 从而减弱了物种之间的交互作用强度, 同时也减弱了干扰在食物网中的传递。因此, 模块性结构最终更有利于复杂食物网中的物种多样性维持和稳定性(Thébault & Fontaine, 2010; Stouffer & Bascompte, 2011)。这一理论结果为理解自然界食物网中相对较高的模块性(相比于互惠网络)提供了一种可能思路(Thébault & Fontaine, 2010)。
食物网中的连接度也影响物种多样性的维持。早期观点认为, 物种连接度越高, 其灭绝引起的次级灭绝就越多, 因此食物网中连接度较高的物种是多样性维持的关键(Dunne et al, 2002)。但该结论是基于静态的食物网结构模型得出的, 并未考虑种群动态以及物种间的交互作用与反馈(Box 1)。Zhao等(2016)结合食物网结构和种群动态模型发现, 对物种多样性维持起重要作用的是能量流更大的物种(也即总能量输入或输出更多的物种)、而非连接度更高的物种。此外, 物种性状也可通过改变种群动态和种间关系来影响食物网的物种多样性维持。基于个体大小的食物网模型表明, 捕食者-猎物的个体大小比例对多样性维持具有重要作用(Brose et al, 2006)。具体地, 捕食者相对于猎物的个体大小比例越大, 则营养级间的能量流相对越缓慢, 从而可增强种群稳定性和食物网的物种多样性。
3 食物网结构与多样性如何影响生态系统功能?
食物网的复杂结构为生态系统内的物质和能量流动提供了多样化的渠道, 从而可能影响生态系统功能。解析食物网结构和多样性对生态系统的调控作用, 是理解生态系统运行机制的关键, 也是整合群落生态学和生态系统生态学的重要途径(Thompson et al, 2012)。本节概述了有关网络结构和物种多样性如何影响食物网功能维持的最新进展。
3.1 食物网结构与生态系统功能
在生物、社会、经济等不同领域的网络中, 网络结构都对维持相应系统的功能具有重要意义(Strogatz, 2001), 食物网中亦然。早期关于营养级联(trophic cascade)的研究, 为理解食物网结构如何影响生态系统功能提供了重要认识。营养级联刻画了食物链中的顶级捕食者对低营养级物种的间接作用。特别地, 顶级捕食者通过下行级联效应可影响初级生产者, 而后者又通过上行效应(bottom-up effect)对高营养级物种产生反馈。过去几十年关于营养级联开展了大量研究, 其中涉及的几个重要问题包括: 营养级联在水生和陆生生态系统中是否都存在、其强度如何(Strong, 1992; Polis et al, 2000); 营养级联在生态系统内和不同生态系统之间的作用机制是什么(Knight et al, 2005; Trussell et al, 2006; Leroux & Loreau, 2008); 顶级捕食者灭绝可能产生怎样的级联效应(Estes et al, 2011)等。孙书存(2017)一文对营养级联理论作了深入阐述。
虽然营养级联效应在食物链模型中得到了很好的支持, 但在复杂食物网中, 杂食性可能大大减弱营养级联的作用。杂食性使得顶级捕食者与低营养级物种间存在多种连接路径, 不同路径上的级联作用可能相互抵消, 从而减弱捕食者对低营养级的调控(Polis & Strong, 1996)。特别地, 杂食性可改变生物量的分布。食物链中顶级捕食者的跨营养级取食, 可导致生物量在营养级间的分布从金字塔型变为“沙漏”型(Woodson et al, 2018)。复杂食物网中的集团内捕食(一种杂食性结构)可减弱高营养级的下行效应(top-down effect)作用, 从而促进低营养级物种的生物量积累, 而后者又通过上行效应增加高营养级的总生物量(Wang S et al, 2019)。然而, 当外界生产力或营养水平较低时, 集团内捕食不利于能量或物质向高营养级传递, 从而导致高营养级的总生物量降低。杂食性表征了捕食者在垂直方向上的生态位幅度, 而其在水平方向上的生态位幅度则由广食性刻画。捕食者的广食性可增强猎物群落内的似然竞争, 从而改变种群动态和生态系统功能。在单营养级群落中, 物种之间的利用性竞争可减弱生态位互补作用(niche complementarity; Loreau, 2010), 不利于生态系统功能的维持。Poisot等(2013)发现, 类似于利用性竞争, 多营养级系统中的似然竞争对生态系统功能有类似的负作用, 提出了营养互补假说(trophic complementarity hypothesis)。具体地, 给定一个植物群落, 食草动物的广食性越高, 则植物种之间的似然竞争越强, 最终植物群落的生物量和生产力越低。
虽然关于食物网的研究有很长的历史, 但以往研究较多关注食物网结构与稳定性的关系, 对网络结构如何影响功能的认识仍较零散。特别地, 在众多的食物网结构和功能指标(见本文第1节)中, 不同结构指标与功能指标之间有何关联, 仍缺乏系统认识(Thompson et al, 2012)。虽然如此, 近期研究关于食物网多样性对生态系统功能的作用取得了一系列新进展, 见3.2节。
3.2 食物网多样性与生态系统功能
生物多样性与生态系统功能是近三十年来生态学领域的研究热点, 为理解生物多样性丧失的生态后果提供了重要认识。虽然最早的生物多样性实验之一就研究了多营养级系统中的生物多样性对生态系统过程的影响(Naeem et al, 1994), 但后期多样性实验大多考虑单营养级系统, 尤其是植物群落。这些实验表明, 植物多样性可促进生态系统初级生产力, 作用机制可归纳为物种间的生态位互补或竞争产生的选择效应(selection effect; Loreau, 2010; Tilman et al, 2014)。然而, 为了理解自然界中的物种丧失的可能后果, 需要在食物网框架下研究生物多样性与功能的关系, 这一方面是因为自然生态系统中的物种都受到营养级间的相互作用, 另一方面是因为高营养级物种面临更大的灭绝风险。虽然早期食物网研究较少关注多样性与生态系统功能的关系, 但过去20年在这一问题上取得了重要进展, 以下从水平多样性和垂直多样性两方面阐述。
水平方向上, 近期研究主要从资源获取和抵抗能力两方面探讨了营养级内的水平多样性的作用。首先, 高营养级内的水平多样性可促进该营养级的能量获取效率和生物量, 降低资源营养级的生物量(Ives et al, 2004; Duffy et al, 2007)。然而, 相比于植物多样性, 高营养级内的水平多样性对功能的促进作用可能较弱, 这是因为不同于植物对非生物资源的获取, 高营养级对低营养级的取食会产生较强的下行调控, 从而引起种群波动, 甚至导致低营养级物种的灭绝(Rosenzweig, 1971; Duffy et al, 2007)。Cardinale等(2006)基于111项生物多样性实验的整合分析却表明, 植物、食草动物、捕食者、分解者等不同营养类群的水平多样性均具有正作用, 即促进自身营养级的生产力和生物量并降低其资源营养级的浓度或生物量; 与植物多样性相比, 其他三个营养级的水平多样性具有相同强度的正作用。这可能是由于该整合分析中的动物群落大多是下行作用相对较弱的无脊椎动物类群(Borer et al, 2005), 也可能是由于营养级内多样性的增加引起了更多的集团内捕食, 从而减弱了对低营养级的下行调控(Wang S et al, 2019)。此外, 高营养级的存在可定性或定量地改变营养级内水平多样性的作用(Thébault & Loreau, 2003; Jiang & Pu, 2009), 但以往的理论和实验结果并未得出一致性结论, 而是发现高营养级的作用受水平和垂直生态位幅度、物种在种群增长和抵抗捕食上是否存在权衡关系等因素影响(Duffy et al, 2007)。另一方面, 营养级内的水平多样性可增强该营养级对其捕食者群落的抵抗能力。比如, 多样性更高的植物群落有更大的概率包含对食草动物抵抗力强的物种, 从而降低食草动物的取食压力(Hillebrand & Cardinale, 2004)。对于寄主-寄生物系统而言, 寄主的多样性可通过降低寄生物的传播速率、抑制被感染的寄主种群等方式产生“稀释作用” (dilution effect), 降低寄主的被感染风险(Civitello et al, 2015)。总而言之, 对某一特定营养级而言, 水平多样性可促进其对低营养级的资源获取效率以及对高营养级的抵抗力。这一作用可通过级联作用传递到更高或更低的营养级, 但由于杂食性的存在和中间营养级行为方式的调整, 多样性作用的强度可能随营养级间的距离增加而大大减弱(Duffy et al, 2007; Haddad et al, 2009)。
垂直方向上, 食物网的最高营养级(即顶级捕食者)可通过直接或间接的下行作用影响整个食物网中的种群动态并最终影响初级生产者的生产力(Wang & Brose, 2018; Eisenhauer et al, 2019)。经典的营养级联理论预测, 顶级捕食者抑制其猎物营养级以及其他与其距离为奇数的营养级, 而释放与其距离为偶数的营养级。因此, 随食物链长度增加, 初级生产者的生产力和生物量呈上升与下降交替的波动变化(Loreau, 2010)。近期研究探讨了复杂食物网模型中垂直多样性的作用, 得出了不同的预测结果。Wang和Brose (2018)发现在复杂食物网中, 初级生产力随最高营养级增加呈单调的指数增长, 提出了“垂直多样性假说” (vertical diversity hypothesis)。食物链和复杂食物网模型之所以给出了不一致的预测结果, 是因为食物链中存在较强的营养级联作用, 而复杂食物网中的杂食性使得营养级联的作用大大减弱(Polis & Strong, 1996; Wang & Brose, 2018)。在复杂食物网中, 垂直多样性的作用机制可从两方面理解: 一方面, 复杂的营养结构可为植物物种提供更多样的生态位, 促进了营养互补效应(Poisot et al, 2013), 进而增强植物群落的营养吸收效率。另一方面, 动物群落的捕食作用可改变植物群落的性状组成, 从而通过选择效应影响植物多样性与初级生产力的关系(Schneider et al, 2016)。特别地, 更高的垂直多样性选择个体更大的植物种, 而由于单位质量的新陈代谢速率随个体大小增加而减小, 因此动物群落的选择作用可提高植物群落的营养利用效率(Wang & Brose, 2018)。
综上, 水平多样性有利于相应的营养级更有效地从低营养级获取资源和抵抗高营养级的捕食, 垂直多样性可通过下行调控作用促进初级生产力。类似于单营养级系统中的多样性作用(Tilman et al, 2014), 食物网中的水平和垂直多样性的作用机制也可通过互补效应和选择效应来认识, 但两种效应的具体实现机制需从营养调控角度来理解。目前整合水平和垂直多样性的研究仍较少, 但理论模型表明, 食物网的总物种多样性可提高生态系统生产力、生物量、总代谢速率等多方面的功能(Schneider et al, 2016; Maureaud et al, 2020)。
4 全球变化如何影响食物网结构、多样性与功能?
全球环境变化和生物入侵等因素可能改变食物网的结构和动态(Tylianakis et al, 2008; Sentis et al, 2014)。Tylianakis等(2008)基于对688篇文献的综述分析表明, 气候和土地利用变化、富营养化(eutrophication)、生物入侵等因素会显著改变不同类型的种间作用关系, 包括植食、捕食、竞争、互惠等, 但改变的方向在不同系统中存在较大差异。深入理解全球变化对食物网的结构与功能的影响, 是预测生态系统对全球变化响应的基础。本节重点介绍近期研究关注较多的两类全球变化因子, 即气候变暖和富营养化。
4.1 气候变暖
气候变暖可通过多种方式影响食物网结构、动态和功能。很多研究表明, 气候变化可引起物种在空间分布范围和物候上的变化(Parmesan & Yohe, 2003)。由于不同物种的响应往往存在差异, 可能引起某些已有的种间关系的消失以及新的种间关系的出现, 从而引起食物网结构的重建(Song et al, 2020)。不同子群落间的响应也可能存在差异, 比如在陆地生态系统中, 增温可能引起地上食物网结构发生较大变化, 而地下食物网结构相对稳健(Thakur, 2020)。认识气候变化下的“新兴种间关系” (novel interactions)和“新兴群落” (novel community)是预测生态系统结构变化的关键(Gilman et al, 2010; Guiden et al, 2019)。
增温可引起食物网中物种性状、种群动态和种间作用强度的变化, 从而影响物种多样性维持。很多研究表明, 增温可导致生物个体变小(Daufresne et al, 2009)。由于个体大小对种群过程和种间关系有重要影响, 因此增温可间接影响食物网动态和多样性维持(Brown et al, 2004)。此外, 增温还可直接影响种群动态与种间关系。一方面, 生态学代谢理论预测, 增温可加速个体生长、新陈代谢和死亡等过程(Brown et al, 2004); 另一方面, 增温还可增大捕食者的攻击速率、降低其对猎物的处理时间, 从而增加整体捕食速率(Box 1; Rall et al, 2012; Fussmann et al, 2014)。虽然增温可同时增加捕食者的捕食速率和代谢速率, 但经验分析显示前者增加的幅度往往小于后者, 所以增温下捕食者的能量需求更难满足, 从而面临更大的灭绝风险(Fussmann et al, 2014)。因此, 增温可能导致物种多样性丧失和食物网营养级的下降(Zhang et al, 2017)。O’Gorman等(2019)利用冰岛14个地热增温的溪流食物网数据, 证实增温可降低总物种数和垂直多样性。该研究还发现, 增温降低了食物网中的连接度, 从而增大次级灭绝风险。然而, 增温并未导致该系统中个体大小的下降, 反而增大了消费者的平均个体大小, 这可能是由于在该系统中生产力也在随温度增加而增加(O’Gorman et al, 2017)。因此, 理解增温的作用需考虑其与其他全球变化因子的交互作用。Binzer等(2016)基于经验数据发展了一个参数化的食物网模型, 该模型预测, 增温在生产力较高的地区可增加物种多样性, 而在生产力较低的地区降低物种多样性。
通过改变网络结构、多样性和种间作用强度, 增温最终可影响生态系统功能。多个实验研究表明, 增温可引起生态系统初级生产力的增加, 进而影响不同营养级间的生物量分布(Petchey et al, 1999; O’Connor et al, 2009)。基于海洋和溪流食物网的研究均发现, 增温加强了消费者的下行调控作用, 促使生物量在营养级间的分布向倒金字塔型转化(O’Connor et al, 2009; O’Gorman et al, 2019)。然而, 增温可能同时改变生产者或消费者的物种组成, 从而影响营养级间的能量传递和下行调控。Petchey等(1999)的微宇宙实验发现, 增温引起高营养级物种灭绝, 因此减弱了下行调控作用, 导致初级生产者的生物量增加。Ullah等(2018)利用室内试验模拟了增温和CO2增加对海洋食物网的作用, 发现增温可引起初级生产者中的蓝细菌生物量增加。由于蓝细菌可产生毒素和化感物质, 因而降低了生产者与食草动物之间的营养传递效率(trophic efficiency), 最终导致生产者生物量增加, 而食草动物和食肉动物营养级的生物量下降。
4.2 富营养化
富营养化对多样性的作用可从两方面理解。一方面, 富营养化可增加初级生产力, 并通过上行作用传递到高营养级(Lindeman, 1942), 从而有利于维持更高的物种数和垂直多样性(Post, 2002; Moore & de Ruiter, 2012)。但另一方面, 富营养化引起的高营养级的生物量增加可增强下行调控作用, 从而降低整个系统的稳定性。特别地, 过高的营养富集可导致捕食者-猎物系统发生震荡, 从而增大物种灭绝的风险, 也即经典的“富集悖论”假说(paradox of enrichment; Rosenzweig, 1971)。基于这两方面作用, 理论模型预测: 富营养化可增加营养贫瘠地区的物种多样性, 而降低营养富集地区的多样性(Rall et al, 2008; Rip & McCann, 2011)。进一步地, 理论分析还预测富营养化与增温之间存在交互作用: 增温可缓解富营养化的负作用, 从而在高温环境下富营养化促进物种多样性(Binzer et al, 2016)。
如上所述, 富营养化可增加初级生产力, 进而促进高营养级的生产力和生物量。此外, 富营养化可增加生产者中的营养元素含量(如氮和磷), 从而提高生产者和初级消费者之间的营养传递效率(Sterner & Elser, 2002)。类似于增温, 富营养化也可能引起物种组成发生变化, 比如增加抵抗力强的物种的比例, 从而限制能量向高营养级传递。Davis等(2010)利用溪流食物网系统开展了为期5年的营养添加实验, 结果发现, 在实验初期, 初级和次级消费者的生物量和生产力都增加, 体现了溪流食物网较高的能量传递效率; 但在实验后期, 虽然初级消费者的生物量和生产力都在增加, 但次级消费者的生产力却开始下降。这是由于, 初级消费者生物量的增加主要来自个体较大的物种, 而次级消费者只能取食个体较小的物种, 从而导致初级和次级消费者之间的营养传递效率降低。
5 研究展望
食物网是生态学领域的一个经典研究议题。过去几十年中, 关于食物网的理论和经验研究都取得了重要进展, 特别是近期研究将早期的简单网络分析拓展至复杂食物网。本文围绕食物网的结构、多样性和功能, 对近期基于复杂食物网的理论进展进行了概述。为了进一步深入理解食物网结构与功能的关系及其各自的形成和维持机制, 未来研究需进一步加强食物网理论与功能性状、化学计量理论、与其他生态网络的整合、并拓展其空间和时间尺度, 从而实现对生态系统复杂性的整体认识。
5.1 物种功能性状
物种功能性状对种群动态和食物网结构与功能有重要影响。Gravel等(2016)提出在食物网研究中, 可根据物种性状的作用将其分成三类: (1)拓扑性状, 即影响网络结构或捕食关系存在与否的性状, 如昆虫口器的形状; (2)消费性状, 即影响植食或捕食速率的性状, 如资源物种中营养组分或元素的含量; (3)生活史性状, 即影响种群增长或死亡的性状, 如繁殖力和成熟时间。此分类只是基于性状作用的一个划分, 很多性状同时具有多方面作用, 因此可被归为多类。比如, 目前食物网文献中研究最多的性状是个体大小, 它不仅可以预测种间捕食关系的有无(Brose et al, 2019)、捕食速率(Rall et al, 2012)、种群增长速率(Brown et al, 2004), 还有助于理解食物网整体的结构复杂性(Laigle et al, 2018)和生态系统功能(Schneider et al, 2016; Wang & Brose, 2018)。因此加强食物网功能性状研究, 有利于进一步整合食物网结构、动态与功能。
5.2 食物网与化学计量学
目前的食物网理论大多基于种群动态模型, 因此可以较好地刻画物种在数量方面的特征, 如种群大小和生物量。但很多研究表明, 捕食关系以及种群动态都受到生物化学计量属性、营养元素含量等质量特征的影响(Sterner & Elser, 2002; Burian et al, 2020)。比如, 食草动物取食营养含量高的植物时有更高的同化效率(Sterner & Elser, 2002), 对氮和钠元素的需求可促进集团内捕食的发生(Denno & Fagan, 2003; Clay et al, 2017)。在两物种或三物种的食物链模型中考虑食物质量的影响, 可显著改变捕食者-猎物动态、物种共存条件、能量传递效率等(Elser et al, 2012; Peace, 2015)。如何在复杂食物网中同时考虑数量与质量的调控作用, 是今后食物网理论发展的一个重要方向。此类模型将促进生物地球化学循环、化学计量、种群动态、食物网等研究领域的整合, 可为理解生态系统过程与功能提供新的认识(Welti et al, 2017)。
5.3 食物网与其他生态网络的整合
自然群落中物种之间除了捕食关系, 还存在互惠(mutualism)、寄生(parasitism)、竞争等种间作用。目前越来越多的研究尝试用网络方法将不同的种间作用整合起来(Fontaine et al, 2011; Mougi & Kondoh, 2012), 比如Pocock等(2012)构建了英格兰西南部农场560种动植物之间的生态网络, 包含捕食、寄生、传粉、种子传播等种间关系。理论分析表明, 种间作用的多样性对生态系统稳定性的维持有重要作用, 比如捕食和互惠关系的同时存在可促进复杂生态系统的稳定性(Mougi & Kondoh, 2012)。另外, 当外界生物或非生物条件发生改变, 种间关系也可能改变甚至相互转化(Holland & DeAngelis, 2009)。一方面, 环境条件的变化可能改变种间关系, 比如近期有实验发现, 随着环境营养浓度的增加, 两种酵母菌株可从低营养条件下的互惠关系转化为高营养条件下的竞争或寄生关系(Hoek et al, 2016)。另一方面, 物种之间可能存在高阶作用, 即某一物种的存在可定量或定性地改变其他物种之间的关系(Goudard & Loreau, 2008; Kéfi et al, 2012)。在生态网络研究中整合不同类型的种间关系及其在生物和非生物条件下的改变与转化是一重要新兴领域。
5.4 集合食物网理论
空间过程对生物多样性和生态系统功能的维持具有重要作用。近20年来, 集合群落理论为理解生境异质性、扩散等因素的作用提供了新的研究框架, 大大增进了对生境破坏的生态效应的认识(Leibold & Chase, 2018)。然而以往集合群落模型大多关注单营养级的竞争群落, 且通常假定物种在不同斑块之间随机扩散。未来研究亟待加强对集合食物网模型的研究, 其中需特别考虑扩散方式和空间异质性(Leibold & Chase, 2018)。与竞争群落不同, 食物网中的不同营养级由于个体大小等性状上的差异, 往往具有不同的扩散速率(McCann et al, 2005; Hirt et al, 2017), 因此模型中需考虑扩散在营养级之间的差异。此外, 对食物网中的物种来说, 其扩散可能受其猎物和捕食者的影响, 即种群倾向于向猎物密度更高、捕食者密度更低的斑块扩散(Fronhofer et al, 2018)。在集合食物网模型中考虑更为现实的扩散情景, 有利于预测气候变化下食物网结构和多样性的响应(Thompson & Gonzalez, 2017)。
5.5 进化模型
虽然食物网模型可以有效地刻画自然界食物网的结构指标, 但这些模型无法解释这些结构是如何产生的。进化模型则可通过模拟物种属性(比如个体大小、捕食宽度等)的演化过程, 帮助我们理解复杂的食物网结构如何从物种进化过程中涌现出来(Loeuille & Loreau, 2005; Ingram et al, 2009)。在全球变化背景下, 生态和进化过程的反馈对于食物网结构和功能有重要作用。比如, 某一物种的适应性进化不仅对其自身的种群维持起作用(比如进化拯救效应; evolutionary rescue), 还对食物网中与其直接相关的捕食者和猎物以及间接相关的其他物种产生影响(Loeuille, 2019)。因此, 构建食物网的生态-进化模型有利于理解全球变化下生态系统的适应性响应。
参考文献
A general model for food web structure
DOI:10.1126/science.1156269 URL [本文引用: 1]
Energy flux: The link between multitrophic biodiversity and ecosystem functioning
DOI:10.1016/j.tree.2017.12.007
URL
PMID:29325921
[本文引用: 2]

Relating biodiversity to ecosystem functioning in natural communities has become a paramount challenge as links between trophic complexity and multiple ecosystem functions become increasingly apparent. Yet, there is still no generalised approach to address such complexity in biodiversity-ecosystem functioning (BEF) studies. Energy flux dynamics in ecological networks provide the theoretical underpinning of multitrophic BEF relationships. Accordingly, we propose the quantification of energy fluxes in food webs as a powerful, universal tool for understanding ecosystem functioning in multitrophic systems spanning different ecological scales. Although the concept of energy flux in food webs is not novel, its application to BEF research remains virtually untapped, providing a framework to foster new discoveries into the determinants of ecosystem functioning in complex systems.
Disentangling the web of life
DOI:10.1126/science.1170749
URL
PMID:19628856
[本文引用: 1]
Biodiversity research typically focuses on species richness and has often neglected interactions, either by assuming that such interactions are homogeneously distributed or by addressing only the interactions between a pair of species or a few species at a time. In contrast, a network approach provides a powerful representation of the ecological interactions among species and highlights their global interdependence. Understanding how the responses of pairwise interactions scale to entire assemblages remains one of the great challenges that must be met as society faces global ecosystem change.
Interactive effects of warming, eutrophication and size structure: Impacts on biodiversity and food-web structure
DOI:10.1111/gcb.13086
URL
PMID:26365694
[本文引用: 2]
Warming and eutrophication are two of the most important global change stressors for natural ecosystems, but their interaction is poorly understood. We used a dynamic model of complex, size-structured food webs to assess interactive effects on diversity and network structure. We found antagonistic impacts: Warming increases diversity in eutrophic systems and decreases it in oligotrophic systems. These effects interact with the community size structure: Communities of similarly sized species such as parasitoid-host systems are stabilized by warming and destabilized by eutrophication, whereas the diversity of size-structured predator-prey networks decreases strongly with warming, but decreases only weakly with eutrophication. Nonrandom extinction risks for generalists and specialists lead to higher connectance in networks without size structure and lower connectance in size-structured communities. Overall, our results unravel interactive impacts of warming and eutrophication and suggest that size structure may serve as an important proxy for predicting the community sensitivity to these global change stressors.
What determines the strength of a trophic cascade?
Allometric scaling enhances stability in complex food webs
Complex food webs prevent competitive exclusion among producer species
Predator traits determine food-web architecture across ecosystems
DOI:10.1038/s41559-019-0899-x
URL
PMID:31110252
[本文引用: 1]
Predator-prey interactions in natural ecosystems generate complex food webs that have a simple universal body-size architecture where predators are systematically larger than their prey. Food-web theory shows that the highest predator-prey body-mass ratios found in natural food webs may be especially important because they create weak interactions with slow dynamics that stabilize communities against perturbations and maintain ecosystem functioning. Identifying these vital interactions in real communities typically requires arduous identification of interactions in complex food webs. Here, we overcome this obstacle by developing predator-trait models to predict average body-mass ratios based on a database comprising 290 food webs from freshwater, marine and terrestrial ecosystems across all continents. We analysed how species traits constrain body-size architecture by changing the slope of the predator-prey body-mass scaling. Across ecosystems, we found high body-mass ratios for predator groups with specific trait combinations including (1) small vertebrates and (2) large swimming or flying predators. Including the metabolic and movement types of predators increased the accuracy of predicting which species are engaged in high body-mass ratio interactions. We demonstrate that species traits explain striking patterns in the body-size architecture of natural food webs that underpin the stability and functioning of ecosystems, paving the way for community-level management of the most complex natural ecosystems.
Toward a metabolic theory of ecology
Food quantity-quality interactions and their impact on consumer behavior and trophic transfer
Effects of biodiversity on the functioning of trophic groups and ecosystems
DOI:10.1038/nature05202
URL
PMID:17066035
[本文引用: 1]
Over the past decade, accelerating rates of species extinction have prompted an increasing number of studies to reduce species diversity experimentally and examine how this alters the efficiency by which communities capture resources and convert those into biomass. So far, the generality of patterns and processes observed in individual studies have been the subjects of considerable debate. Here we present a formal meta-analysis of studies that have experimentally manipulated species diversity to examine how it affects the functioning of numerous trophic groups in multiple types of ecosystem. We show that the average effect of decreasing species richness is to decrease the abundance or biomass of the focal trophic group, leading to less complete depletion of resources used by that group. At the same time, analyses reveal that the standing stock of, and resource depletion by, the most species-rich polyculture tends to be no different from that of the single most productive species used in an experiment. Of the known mechanisms that might explain these trends, results are most consistent with what is called the 'sampling effect', which occurs when diverse communities are more likely to contain and become dominated by the most productive species. Whether this mechanism is widespread in natural communities is currently controversial. Patterns we report are remarkably consistent for four different trophic groups (producers, herbivores, detritivores and predators) and two major ecosystem types (aquatic and terrestrial). Collectively, our analyses suggest that the average species loss does indeed affect the functioning of a wide variety of organisms and ecosystems, but the magnitude of these effects is ultimately determined by the identity of species that are going extinct.
Phylogenetic constraints and adaptation explain food-web structure
DOI:10.1038/nature02327
URL
PMID:14985761
[本文引用: 1]
Food webs are descriptions of who eats whom in an ecosystem. Although extremely complex and variable, their structure possesses basic regularities. A fascinating question is to find a simple model capturing the underlying processes behind these repeatable patterns. Until now, two models have been devised for the description of trophic interactions within a natural community. Both are essentially based on the concept of ecological niche, with the consumers organized along a single niche dimension; for example, prey size. Unfortunately, they fail to describe adequately recent and high-quality data. Here, we propose a new model built on the hypothesis that any species' diet is the consequence of phylogenetic constraints and adaptation. Simple rules incorporating both concepts yield food webs whose structure is very close to real data. Consumers are organized in groups forming a nested hierarchy, which better reflects the complexity and multidimensionality of most natural systems.
Research progress on ecosystem complexity-stability relationships based on soil food web
基于土壤食物网的生态系统复杂性-稳定性关系研究进展
The interaction between predation and competition
Biodiversity inhibits parasites: Broad evidence for the dilution effect
Towards a geography of omnivory: Omnivores increase carnivory when sodium is limiting
A stochastic theory of community food webs. I. Models and aggregated data
Global warming benefits the small in aquatic ecosystems
Long-term nutrient enrichment decouples predator and prey production
Modelling food webs and nutrient cycling in agro-ecosystems
DOI:10.1016/0169-5347(94)90059-0
URL
PMID:21236897
[本文引用: 1]
Agricultural practices affect the spatial patterns and dynamics of the decomposition of soil organic matter and the availability of plant-limiting nutrients. The biological processes underlying these patterns and dynamics are the trophic interactions among the organisms in the soil community food web. Food web models simulate nutrient flow rates close to observed rates and clarify the role of the various groups of organisms in the cycling of nutrients. Several large interdisciplinary programs are currently focusing on these interactions, with a view to developing and managing sustainable forms of agriculture.
Might nitrogen limitation promote omnivory among carnivorous arthropods?
The functional role of biodiversity in ecosystems: Incorporating trophic complexity
DOI:10.1111/j.1461-0248.2007.01037.x
URL
PMID:17498151
[本文引用: 6]
Understanding how biodiversity affects functioning of ecosystems requires integrating diversity within trophic levels (horizontal diversity) and across trophic levels (vertical diversity, including food chain length and omnivory). We review theoretical and experimental progress toward this goal. Generally, experiments show that biomass and resource use increase similarly with horizontal diversity of either producers or consumers. Among prey, higher diversity often increases resistance to predation, due to increased probability of including inedible species and reduced efficiency of specialist predators confronted with diverse prey. Among predators, changing diversity can cascade to affect plant biomass, but the strength and sign of this effect depend on the degree of omnivory and prey behaviour. Horizontal and vertical diversity also interact: adding a trophic level can qualitatively change diversity effects at adjacent levels. Multitrophic interactions produce a richer variety of diversity-functioning relationships than the monotonic changes predicted for single trophic levels. This complexity depends on the degree of consumer dietary generalism, trade-offs between competitive ability and resistance to predation, intraguild predation and openness to migration. Although complementarity and selection effects occur in both animals and plants, few studies have conclusively documented the mechanisms mediating diversity effects. Understanding how biodiversity affects functioning of complex ecosystems will benefit from integrating theory and experiments with simulations and network-based approaches.
Network structure and biodiversity loss in food webs: Robustness increases with connectance
A multitrophic perspective on biodiversity-ecosystem functioning research
Lotka re-loaded: Modeling trophic interactions under stoichiometric constraints
Trophic downgrading of planet Earth
DOI:10.1126/science.1205106
URL
[本文引用: 1]
Until recently, large apex consumers were ubiquitous across the globe and had been for millions of years. The loss of these animals may be humankind's most pervasive influence on nature. Although such losses are widely viewed as an ethical and aesthetic problem, recent research reveals extensive cascading effects of their disappearance in marine, terrestrial, and freshwater ecosystems worldwide. This empirical work supports long-standing theory about the role of top-down forcing in ecosystems but also highlights the unanticipated impacts of trophic cascades on processes as diverse as the dynamics of disease, wildfire, carbon sequestration, invasive species, and biogeochemical cycles. These findings emphasize the urgent need for interdisciplinary research to forecast the effects of trophic downgrading on process, function, and resilience in global ecosystems.
The ecological and evolutionary implications of merging different types of networks
DOI:10.1111/j.1461-0248.2011.01688.x
URL
[本文引用: 1]
Interactions among species drive the ecological and evolutionary processes in ecological communities. These interactions are effectively key components of biodiversity. Studies that use a network approach to study the structure and dynamics of communities of interacting species have revealed many patterns and associated processes. Historically these studies were restricted to trophic interactions, although network approaches are now used to study a wide range of interactions, including for example the reproductive mutualisms. However, each interaction type remains studied largely in isolation from others. Merging the various interaction types within a single integrative framework is necessary if we want to further our understanding of the ecological and evolutionary dynamics of communities. Dividing the networks up is a methodological convenience as in the field the networks occur together in space and time and will be linked by shared species. Herein, we outline a conceptual framework for studying networks composed of more than one type of interaction, highlighting key questions and research areas that would benefit from their study.
Bottom-up and top-down control of dispersal across major organismal groups
DOI:10.1038/s41559-018-0686-0
URL
PMID:30397298
[本文引用: 1]
Ecology and evolution unfold in spatially structured communities, where dispersal links dynamics across scales. Because dispersal is multicausal, identifying general drivers remains challenging. In a coordinated distributed experiment spanning organisms from protozoa to vertebrates, we tested whether two fundamental determinants of local dynamics, top-down and bottom-up control, generally explain active dispersal. We show that both factors consistently increased emigration rates and use metacommunity modelling to highlight consequences on local and regional dynamics.
Ecological stability in response to warming
Food-web aggregation, methodological and functional issues
Fluxweb: An R package to easily estimate energy fluxes in food webs
A framework for community interactions under climate change
Nontrophic interactions, biodiversity, and ecosystem functioning: An interaction web model
DOI:10.1086/523945
URL
PMID:18171154
[本文引用: 1]
Research into the relationship between biodiversity and ecosystem functioning has mainly focused on the effects of species diversity on ecosystem properties in plant communities and, more recently, in food webs. Although there is growing recognition of the significance of nontrophic interactions in ecology, these interactions are still poorly studied theoretically, and their impact on biodiversity and ecosystem functioning is largely unknown. Existing models of mutualism usually consider only one type of species interaction and do not satisfy mass balance constraints. Here, we present a model of an interaction web that includes both trophic and nontrophic interactions and that respects the principle of mass conservation. Nontrophic interactions are represented in the form of interaction modifications. We use this model to study the relationship between biodiversity and ecosystem properties that emerges from the assembly of entire interaction webs. We show that ecosystem properties such as biomass and production depend not only on species diversity but also on species interactions, in particular on the connectance and magnitude of nontrophic interactions, and that the nature, prevalence, and strength of species interactions in turn depend on species diversity. Nontrophic interactions alter the shape of the relationship between biodiversity and biomass and can profoundly influence ecosystem processes.
The meaning of functional trait composition of food webs for ecosystem functioning
Predator-prey interactions in the anthropocene: Reconciling multiple aspects of novelty
Plant species loss decreases arthropod diversity and shifts trophic structure
Consumer effects decline with prey diversity
A general scaling law reveals why the largest animals are not the fastest
Resource availability modulates the cooperative and competitive nature of a microbial cross-feeding mutualism
Consumer-resource theory predicts dynamic transitions between outcomes of interspecific interactions
DOI:10.1111/j.1461-0248.2009.01390.x
URL
PMID:19807773
[本文引用: 1]
Interactions between two populations are often defined by their interaction outcomes; that is, the positive, neutral, or negative effects of species on one another. Yet, signs of outcomes are not absolute, but vary with the biotic and abiotic contexts of interactions. Here, we develop a general theory for transitions between outcomes based on consumer-resource (C-R) interactions in which one or both species exploit the other as a resource. Simple models of C-R interactions revealed multiple equilibria, including one for species coexistence and others for extinction of one or both species, indicating that species' densities alone could determine the fate of interactions. All possible outcomes [(+ +), (+ -), (--), (+ 0), (- 0), (0 0)] of species coexistence emerged merely through changes in parameter values of C-R interactions, indicating that variation in C-R interactions resulting from biotic and abiotic conditions could determine shifts in outcomes. These results suggest that C-R interactions can provide a broad mechanism for understanding context- and density-dependent transitions between interaction outcomes.
Some characteristics of simple types of predation and parasitism
Simple rules for interspecific dominance in systems with exploitative and apparent competition
Niche evolution, trophic structure, and species turnover in model food webs
DOI:10.1086/599301
URL
PMID:19459779
[本文引用: 1]
The features that govern the stability and persistence of species interaction networks, such as food webs, remain elusive, but recent work suggests that the distribution and strength of trophic links play an important role. Potential omnivory-stability relationships have been investigated and debated extensively, but we still have a relatively poor understanding of how levels of omnivory relate to the stability of diverse food webs. Here, we use an evolutionary assembly model to investigate how different trade-offs in resource use influence both food web structure and dynamic stability during the assembly process. We build on a previous model by allowing speciation along with the evolution of two traits: body size and feeding-niche width. Across a wide range of conditions, the level of omnivory in a food web is positively related to its dynamic instability (variability and species turnover). Parameter values favoring omnivory also allow a wider range of phenotypes to invade, often displacing existing species. This high species turnover leaves signatures in reconstructed phylogenies, with shorter branches connecting extant species in more omnivorous food webs. Our findings suggest that features of the environment may influence both trophic structure and dynamic stability, leading to emergent omnivory-stability relationships.
A synthesis of subdisciplines: Predator-prey interactions, and biodiversity and ecosystem functioning
Different effects of species diversity on temporal stability in single-trophic and multitrophic communities
DOI:10.1086/605961
URL
PMID:19775227
[本文引用: 1]
The question of how species diversity affects ecological stability has long interested ecologists and yet remains largely unresolved. Historically, attempts to answer this question have been hampered by the presence of multiple potentially confounding stability concepts, confusion over responses at different levels of ecological organization, discrepancy between theoretical predictions, and, particularly, the paucity of empirical studies. Here we used meta-analyses to synthesize results of empirical studies published primarily in the past 2 decades on the relationship between species diversity and temporal stability. We show that the overall effect of increasing diversity was positive for community-level temporal stability but neutral for population-level temporal stability. There were, however, striking differences in the diversity-stability relationship between single- and multitrophic systems, with diversity stabilizing both population and community dynamics in multitrophic but not single-trophic communities. These patterns were broadly equivalent across experimental and observational studies as well as across terrestrial and aquatic studies. We discuss possible mechanisms for population stability to increase with diversity in multitrophic systems and for diversity to influence community-level stability in general. Overall, our results indicate that diversity can affect temporal stability, but the effects may critically depend on trophic complexity.
Trophic coherence determines food-web stability
Body masses, functional responses and predator-prey stability
DOI:10.1111/ele.12147
URL
PMID:23819684
[本文引用: 1]
The stability of ecological communities depends strongly on quantitative characteristics of population interactions (type-II vs. type-III functional responses) and the distribution of body masses across species. Until now, these two aspects have almost exclusively been treated separately leaving a substantial gap in our general understanding of food webs. We analysed a large data set of arthropod feeding rates and found that all functional-response parameters depend on the body masses of predator and prey. Thus, we propose generalised functional responses which predict gradual shifts from type-II predation of small predators on equally sized prey to type-III functional-responses of large predators on small prey. Models including these generalised functional responses predict population dynamics and persistence only depending on predator and prey body masses, and we show that these predictions are strongly supported by empirical data on forest soil food webs. These results help unravelling systematic relationships between quantitative population interactions and large-scale community patterns.
More than a meal… integrating non-feeding interactions into food webs
DOI:10.1111/j.1461-0248.2011.01732.x
URL
PMID:22313549
[本文引用: 1]
Trophic cascades across ecosystems
DOI:10.1038/nature03962
URL
PMID:16208370
[本文引用: 1]
Predation can be intense, creating strong direct and indirect effects throughout food webs. In addition, ecologists increasingly recognize that fluxes of organisms across ecosystem boundaries can have major consequences for community dynamics. Species with complex life histories often shift habitats during their life cycles and provide potent conduits coupling ecosystems. Thus, local interactions that affect predator abundance in one ecosystem (for example a larval habitat) may have reverberating effects in another (for example an adult habitat). Here we show that fish indirectly facilitate terrestrial plant reproduction through cascading trophic interactions across ecosystem boundaries. Fish reduce larval dragonfly abundances in ponds, leading to fewer adult dragonflies nearby. Adult dragonflies consume insect pollinators and alter their foraging behaviour. As a result, plants near ponds with fish receive more pollinator visits and are less pollen limited than plants near fish-free ponds. Our results confirm that strong species interactions can reverberate across ecosystems, and emphasize the importance of landscape-level processes in driving local species interactions.
Compartments revealed in food-web structure
Species traits as drivers of food web structure
Subsidy hypothesis and strength of trophic cascades across ecosystems
Evolutionary emergence of size-structured food webs
Eco-evolutionary dynamics in a disturbed world: Implications for the maintenance of ecological networks
Trait-based food web model reveals the underlying mechanisms of biodiversity-ecosystem functioning relationships
The dynamics of spatially coupled food webs
DOI:10.1111/j.1461-0248.2005.00742.x
URL
PMID:21352455
[本文引用: 1]
The dynamics of ecological systems include a bewildering number of biotic interactions that unfold over a vast range of spatial scales. Here, employing simple and general empirical arguments concerning the nature of movement, trophic position and behaviour we outline a general theory concerning the role of space and food web structure on food web stability. We argue that consumers link food webs in space and that this spatial structure combined with relatively rapid behavioural responses by consumers can strongly influence the dynamics of food webs. Employing simple spatially implicit food web models, we show that large mobile consumers are inordinately important in determining the stability, or lack of it, in ecosystems. More specifically, this theory suggests that mobile higher order organisms are potent stabilizers when embedded in a variable, and expansive spatial structure. However, when space is compressed and higher order consumers strongly couple local habitats then mobile consumers can have an inordinate destabilizing effect. Preliminary empirical arguments show consistency with this general theory.
Diversity of interaction types and ecological community stability
DOI:10.1126/science.1220529
URL
PMID:22822151
[本文引用: 2]
Ecological theory predicts that a complex community formed by a number of species is inherently unstable, guiding ecologists to identify what maintains species diversity in nature. Earlier studies often assumed a community with only one interaction type, either an antagonistic, competitive, or mutualistic interaction, leaving open the question of what the diversity of interaction types contributes to the community maintenance. We show theoretically that the multiple interaction types might hold the key to understanding community dynamics. A moderate mixture of antagonistic and mutualistic interactions can stabilize population dynamics. Furthermore, increasing complexity leads to increased stability in a
Declining biodiversity can alter the performance of ecosystems
Warming and resource availability shift food web structure and metabolism
DOI:10.1371/journal.pbio.1000178
URL
PMID:19707271
[本文引用: 2]
Climate change disrupts ecological systems in many ways. Many documented responses depend on species' life histories, contributing to the view that climate change effects are important but difficult to characterize generally. However, systematic variation in metabolic effects of temperature across trophic levels suggests that warming may lead to predictable shifts in food web structure and productivity. We experimentally tested the effects of warming on food web structure and productivity under two resource supply scenarios. Consistent with predictions based on universal metabolic responses to temperature, we found that warming strengthened consumer control of primary production when resources were augmented. Warming shifted food web structure and reduced total biomass despite increases in primary productivity in a marine food web. In contrast, at lower resource levels, food web production was constrained at all temperatures. These results demonstrate that small temperature changes could dramatically shift food web dynamics and provide a general, species-independent mechanism for ecological response to environmental temperature change.
Unexpected changes in community size structure in a natural warming experiment
A simple model predicts how warming simplifies wild food webs
A globally coherent fingerprint of climate change impacts across natural systems
DOI:10.1038/nature01286
URL
PMID:12511946
[本文引用: 1]
Causal attribution of recent biological trends to climate change is complicated because non-climatic influences dominate local, short-term biological changes. Any underlying signal from climate change is likely to be revealed by analyses that seek systematic trends across diverse species and geographic regions; however, debates within the Intergovernmental Panel on Climate Change (IPCC) reveal several definitions of a 'systematic trend'. Here, we explore these differences, apply diverse analyses to more than 1,700 species, and show that recent biological trends match climate change predictions. Global meta-analyses documented significant range shifts averaging 6.1 km per decade towards the poles (or metres per decade upward), and significant mean advancement of spring events by 2.3 days per decade. We define a diagnostic fingerprint of temporal and spatial 'sign-switching' responses uniquely predicted by twentieth century climate trends. Among appropriate long-term/large-scale/multi-species data sets, this diagnostic fingerprint was found for 279 species. This suite of analyses generates 'very high confidence' (as laid down by the IPCC) that climate change is already affecting living systems.
Effects of light, nutrients, and food chain length on trophic efficiencies in simple stoichiometric aquatic food chain models
Ontogenetic scaling of foraging rates and the dynamics of a size-structured consumer-resource model
DOI:10.1006/tpbi.1998.1380
URL
PMID:9878605
[本文引用: 1]
The ontogenetic scaling of foraging capacity strongly influences the competitive ability of differently sized individuals within a species. We develop a physiologically structured model to investigate the effect of different ontogenetic size scalings of the attack rate on the population dynamics of a consumer-resource system. The resource is assumed to reproduce continuously whereas the consumer only reproduces at discrete time instants. Depending on the ontogenetic size scaling, the model exhibited recruit-driven cycles, stable fixed point dynamics, non-recruit juvenile-driven cycles, quasiperiodic orbits, or chaotic dynamics. The kind of dynamics observed was related to the maintenance resource levels required of differently sized individuals. Stable fixed point dynamics was, besides at the persistence boundary, only observed when the minimum resource levels were similar for newborns and mature individuals. The tendency for large population fluctuations over a wide range of the parameter space was due to the consumer's pulsed reproduction. Background mortality and length of season were major determinants of cycle length. Model dynamics strongly resembled empirically observed dynamics from fish and Daphnia populations with respect to both patterns and mechanisms. The non-recruit juvenile-driven dynamics is suggested to occur in populations with size-dependent interference or preemptive competition like cicada populations.
Environmental warming alters food-web structure and ecosystem function
The robustness and restoration of a network of ecological networks
DOI:10.1126/science.1214915
URL
PMID:22363009
[本文引用: 1]
Understanding species' interactions and the robustness of interaction networks to species loss is essential to understand the effects of species' declines and extinctions. In most studies, different types of networks (such as food webs, parasitoid webs, seed dispersal networks, and pollination networks) have been studied separately. We sampled such multiple networks simultaneously in an agroecosystem. We show that the networks varied in their robustness; networks including pollinators appeared to be particularly fragile. We show that, overall, networks did not strongly covary in their robustness, which suggests that ecological restoration (for example, through agri-environment schemes) benefitting one functional group will not inevitably benefit others. Some individual plant species were disproportionately well linked to many other species. This type of information can be used in restoration management, because it identifies the plant taxa that can potentially lead to disproportionate gains in biodiversity.
Trophic complementarity drives the biodiversity-ecosystem functioning relationship in food webs
DOI:10.1111/ele.12118
URL
PMID:23692591
[本文引用: 2]
The biodiversity-ecosystem functioning (BEF) relationship is central in community ecology. Its drivers in competitive systems (sampling effect and functional complementarity) are intuitive and elegant, but we lack an integrative understanding of these drivers in complex ecosystems. Because networks encompass two key components of the BEF relationship (species richness and biomass flow), they provide a key to identify these drivers, assuming that we have a meaningful measure of functional complementarity. In a network, diversity can be defined by species richness, the number of trophic levels, but perhaps more importantly, the diversity of interactions. In this paper, we define the concept of trophic complementarity (TC), which emerges through exploitative and apparent competition processes, and study its contribution to ecosystem functioning. Using a model of trophic community dynamics, we show that TC predicts various measures of ecosystem functioning, and generate a range of testable predictions. We find that, in addition to the number of species, the structure of their interactions needs to be accounted for to predict ecosystem productivity.
Food web complexity and community dynamics
When is a trophic cascade a trophic cascade?
DOI:10.1016/s0169-5347(00)01971-6 URL PMID:11050351 [本文引用: 1]
The long and short of food-chain length
Food-web connectance and predator interference dampen the paradox of enrichment
Universal temperature and body-mass scaling of feeding rates
Cross-ecosystem differences in stability and the principle of energy flux
Paradox of enrichment: Destabilization of exploitation ecosystems in ecological times
Animal diversity and ecosystem functioning in dynamic food webs
The necessity of multitrophic approaches in community ecology
DOI:10.1016/j.tree.2018.07.001
URL
PMID:30146326
[本文引用: 1]
Trophic interactions are a fundamental part of ecosystems; yet, most ecological studies focus on single trophic levels and this hampers our ability to detect the underlying mechanisms structuring communities as well as the effects of environmental change. Here, we argue that the historical dominance of studying competition within trophic levels, and the focus on taxonomic groups without differentiating the trophic level, has led to the under-representation of multitrophic research in community ecology. There are many hurdles that challenge multitrophic approaches and we discuss solutions to overcome these. To advance our understanding of the fundamental drivers of community assembly and to provide the necessary guidance for managing and mitigating the effects of environmental change, we argue that ecologists should better align research with a trophically inclusive definition of a community.
Towards a mechanistic understanding of temperature and enrichment effects on species interaction strength, omnivory and food-web structure
DOI:10.1111/ele.12281
URL
PMID:24751223
[本文引用: 2]
Revealing the links between species functional traits, interaction strength and food-web structure is of paramount importance for understanding and predicting the relationships between food-web diversity and stability in a rapidly changing world. However, little is known about the interactive effects of environmental perturbations on individual species, trophic interactions and ecosystem functioning. Here, we combined modelling and laboratory experiments to investigate the effects of warming and enrichment on a terrestrial tritrophic system. We found that the food-web structure is highly variable and switches between exploitative competition and omnivory depending on the effects of temperature and enrichment on foraging behaviour and species interaction strength. Our model contributes to identifying the mechanisms that explain how environmental effects cascade through the food web and influence its topology. We conclude that considering environmental factors and flexible food-web structure is crucial to improve our ability to predict the impacts of global changes on ecosystem diversity and stability.
Towards a probabilistic understanding about the context-dependency of species interactions
DOI:10.1016/j.tree.2019.12.011
URL
PMID:32007296
[本文引用: 1]
Observational and experimental studies have shown that an interaction class between two species (be it mutualistic, competitive, antagonistic, or neutral) may switch to a different class, depending on the biotic and abiotic factors within which species are observed. This complexity arising from the evidence of context-dependencies has underscored a difficulty in establishing a systematic analysis about the extent to which species interactions are expected to switch in nature and experiments. Here, we propose an overarching theoretical framework, by integrating probabilistic and structural approaches, to establish null expectations about switches of interaction classes across environmental contexts. This integration provides a systematic platform upon which it is possible to establish new hypotheses, clear predictions, and quantifiable expectations about the context-dependency of species interactions.
Understanding food-web persistence from local to global scales
DOI:10.1111/j.1461-0248.2009.01407.x
URL
PMID:19968697
[本文引用: 1]
Understanding food-web persistence is an important long-term objective of ecology because of its relevance in maintaining biodiversity. To date, many dynamic studies of food-web behaviour--both empirical and theoretical--have focused on smaller sub-webs, called trophic modules, because these modules are more tractable experimentally and analytically than whole food webs. The question remains to what degree studies of trophic modules are relevant to infer the persistence of entire food webs. Four trophic modules have received particular attention in the literature: tri-trophic food chains, omnivory, exploitative competition, and apparent competition. Here, we integrate analysis of these modules' dynamics in isolation with those of whole food webs to directly assess the appropriateness of scaling from modules to food webs. We find that there is not a direct, one-to-one, relationship between the relative persistence of modules in isolation and their effect on persistence of an entire food web. Nevertheless, we observe that those modules which are most commonly found in empirical food webs are those that confer the greatest community persistence. As a consequence, we demonstrate that there may be significant dynamic justifications for empirically-observed food-web structure.
Compartmentalization increases food-web persistence
Are trophic cascades all wet? Differentiation and donor-control in speciose ecosystems
食物网结构与生态功能. 见: 生态学的现状与发展趋势 (于振良, 葛剑平, 于贵瑞, 张大勇, 方精云, 康乐, 傅伯杰, 焦念志编)
Climate warming and trophic mismatches in terrestrial ecosystems: The green-brown imbalance hypothesis
Food-web constraints on biodiversity-ecosystem functioning relationships
Stability of ecological communities and the architecture of mutualistic and trophic networks
DOI:10.1126/science.1188321
URL
PMID:20705861
[本文引用: 2]
Research on the relationship between the architecture of ecological networks and community stability has mainly focused on one type of interaction at a time, making difficult any comparison between different network types. We used a theoretical approach to show that the network architecture favoring stability fundamentally differs between trophic and mutualistic networks. A highly connected and nested architecture promotes community stability in mutualistic networks, whereas the stability of trophic networks is enhanced in compartmented and weakly connected architectures. These theoretical predictions are supported by a meta-analysis on the architecture of a large series of real pollination (mutualistic) and herbivory (trophic) networks. We conclude that strong variations in the stability of architectural patterns constrain ecological networks toward different architectures, depending on the type of interaction.
Trophic levels and trophic tangles: The prevalence of omnivory in real food webs
DOI:10.1890/05-1454
URL
PMID:17503589
[本文引用: 1]
The concept of trophic levels is one of the oldest in ecology and informs our understanding of energy flow and top-down control within food webs, but it has been criticized for ignoring omnivory. We tested whether trophic levels were apparent in 58 real food webs in four habitat types by examining patterns of trophic position. A large proportion of taxa (64.4%) occupied integer trophic positions, suggesting that discrete trophic levels do exist. Importantly however, the majority of those trophic positions were aggregated around integer values of 0 and 1, representing plants and herbivores. For the majority of the real food webs considered here, secondary consumers were no more likely to occupy an integer trophic position than in randomized food webs. This means that, above the herbivore trophic level, food webs are better characterized as a tangled web of omnivores. Omnivory was most common in marine systems, rarest in streams, and intermediate in lakes and terrestrial food webs. Trophic-level-based concepts such as trophic cascades may apply to systems with short food chains, but they become less valid as food chains lengthen.
Food webs: Reconciling the structure and function of biodiversity
DOI:10.1016/j.tree.2012.08.005
URL
PMID:22959162
[本文引用: 3]
The global biodiversity crisis concerns not only unprecedented loss of species within communities, but also related consequences for ecosystem function. Community ecology focuses on patterns of species richness and community composition, whereas ecosystem ecology focuses on fluxes of energy and materials. Food webs provide a quantitative framework to combine these approaches and unify the study of biodiversity and ecosystem function. We summarise the progression of food-web ecology and the challenges in using the food-web approach. We identify five areas of research where these advances can continue, and be applied to global challenges. Finally, we describe what data are needed in the next generation of food-web studies to reconcile the structure and function of biodiversity.
Dispersal governs the reorganization of ecological networks under environmental change
Biodiversity and ecosystem functioning
Ecosystem ecology: Size-based constraints on the pyramids of life
DOI:10.1016/j.tree.2013.03.008
URL
PMID:23623003
[本文引用: 1]
Biomass distribution and energy flow in ecosystems are traditionally described with trophic pyramids, and increasingly with size spectra, particularly in aquatic ecosystems. Here, we show that these methods are equivalent and interchangeable representations of the same information. Although pyramids are visually intuitive, explicitly linking them to size spectra connects pyramids to metabolic and size-based theory, and illuminates size-based constraints on pyramid shape. We show that bottom-heavy pyramids should predominate in the real world, whereas top-heavy pyramids indicate overestimation of predator abundance or energy subsidies. Making the link to ecological pyramids establishes size spectra as a central concept in ecosystem ecology, and provides a powerful framework both for understanding baseline expectations of community structure and for evaluating future scenarios under climate change and exploitation.
Habitat effects on the relative importance of trait- and density-mediated indirect interactions
DOI:10.1111/j.1461-0248.2006.00981.x
URL
PMID:17040327
[本文引用: 1]
Classical views of trophic cascades emphasize the primacy of consumptive predator effects on prey populations to the transmission of indirect effects [density-mediated indirect interactions (DMIIs)]. However, trophic cascades can also emerge without changes in the density of interacting species because of non-consumptive predator effects on prey traits such as foraging behaviour [trait-mediated indirect interactions (TMIIs)]. Although ecologists appreciate this point, measurements of the relative importance of each indirect predator effect are rare. Experiments with a three-level, rocky shore food chain containing an invasive predatory crab (Carcinus maenas), an intermediate consumer (the snail, Nucella lapillus) and a basal resource (the barnacle, Semibalanus balanoides) revealed that the strength of TMIIs is comparable with, or exceeds, that of DMIIs. Moreover, the sign and strength of each indirect predator effect depends on whether it is measured in risky or refuge habitats. Because habitat shifts are often responsible for the emergence of TMIIs, attention to the sign and strength of these interactions in both habitats will improve our understanding of the link between individual behaviour and community dynamics.
Global change and species interactions in terrestrial ecosystems
DOI:10.1111/j.1461-0248.2008.01250.x
URL
PMID:19062363
[本文引用: 3]
The main drivers of global environmental change (CO2 enrichment, nitrogen deposition, climate, biotic invasions and land use) cause extinctions and alter species distributions, and recent evidence shows that they exert pervasive impacts on various antagonistic and mutualistic interactions among species. In this review, we synthesize data from 688 published studies to show that these drivers often alter competitive interactions among plants and animals, exert multitrophic effects on the decomposer food web, increase intensity of pathogen infection, weaken mutualisms involving plants, and enhance herbivory while having variable effects on predation. A recurrent finding is that there is substantial variability among studies in both the magnitude and direction of effects of any given GEC driver on any given type of biotic interaction. Further, we show that higher order effects among multiple drivers acting simultaneously create challenges in predicting future responses to global environmental change, and that extrapolating these complex impacts across entire networks of species interactions yields unanticipated effects on ecosystems. Finally, we conclude that in order to reliably predict the effects of GEC on community and ecosystem processes, the greatest single challenge will be to determine how biotic and abiotic context alters the direction and magnitude of GEC effects on biotic interactions.
Climate change could drive marine food web collapse through altered trophic flows and cyanobacterial proliferation
Diversifying livestock promotes multidiversity and multifunctionality in managed grasslands
Biodiversity and ecosystem functioning in food webs: The vertical diversity hypothesis
DOI:10.1111/ele.12865
URL
PMID:29057554
[本文引用: 11]
One challenge in merging community and ecosystem ecology is to integrate the complexity of natural multitrophic communities into concepts of ecosystem functioning. Here, we combine food-web and allometry theories to demonstrate that primary production, as measured by the total nutrient uptake of the multitrophic community, is determined by vertical diversity (i.e. food web's maximum trophic level) and structure (i.e. distributions of species and their abundances and metabolic rates across trophic levels). In natural ecosystems, the community size distribution determines all these vertical patterns and thus the total nutrient uptake. Our model suggests a vertical diversity hypothesis (VDH) for ecosystem functioning in complex food webs. It predicts that, under a given nutrient supply, the total nutrient uptake increases exponentially with the maximum trophic level in the food web and it increases with its maximum body size according to a power law. The VDH highlights the effect of top-down regulation on plant nutrient uptake, which complements traditional paradigms that emphasised the bottom-up effect of nutrient supply on vertical diversity. We conclude that the VDH contributes to a synthetic framework for understanding the relationship between vertical diversity and ecosystem functioning in food webs and predicting the impacts of global changes on multitrophic ecosystems.
Intraguild predation enhances biodiversity and functioning in complex food webs
Bridging food webs, ecosystem metabolism, and biogeochemistry using ecological stoichiometry theory
DOI:10.3389/fmicb.2017.01298
URL
PMID:28747904
[本文引用: 1]
Although aquatic ecologists and biogeochemists are well aware of the crucial importance of ecosystem functions, i.e., how biota drive biogeochemical processes and vice-versa, linking these fields in conceptual models is still uncommon. Attempts to explain the variability in elemental cycling consequently miss an important biological component and thereby impede a comprehensive understanding of the underlying processes governing energy and matter flow and transformation. The fate of multiple chemical elements in ecosystems is strongly linked by biotic demand and uptake; thus, considering elemental stoichiometry is important for both biogeochemical and ecological research. Nonetheless, assessments of ecological stoichiometry (ES) often focus on the elemental content of biota rather than taking a more holistic view by examining both elemental pools and fluxes (e.g., organismal stoichiometry and ecosystem process rates). ES theory holds the promise to be a unifying concept to link across hierarchical scales of patterns and processes in ecology, but this has not been fully achieved. Therefore, we propose connecting the expertise of aquatic ecologists and biogeochemists with ES theory as a common currency to connect food webs, ecosystem metabolism, and biogeochemistry, as they are inherently concatenated by the transfer of carbon, nitrogen, and phosphorous through biotic and abiotic nutrient transformation and fluxes. Several new studies exist that demonstrate the connections between food web ecology, biogeochemistry, and ecosystem metabolism. In addition to a general introduction into the topic, this paper presents examples of how these fields can be combined with a focus on ES. In this review, a series of concepts have guided the discussion: (1) changing biogeochemistry affects trophic interactions and ecosystem processes by altering the elemental ratios of key species and assemblages; (2) changing trophic dynamics influences the transformation and fluxes of matter across environmental boundaries; (3) changing ecosystem metabolism will alter the chemical diversity of the non-living environment. Finally, we propose that using ES to link nutrient cycling, trophic dynamics, and ecosystem metabolism would allow for a more holistic understanding of ecosystem functions in a changing environment.
Limits to trophic levels and omnivory in complex food webs: Theory and data
DOI:10.1086/381964
URL
PMID:15026980
[本文引用: 1]
While trophic levels have found broad application throughout ecology, they are also in much contention on analytical and empirical grounds. Here, we use a new generation of data and theory to examine long-standing questions about trophic-level limits and degrees of omnivory. The data include food webs of the Chesapeake Bay, U.S.A., the island of Saint Martin, a U.K. grassland, and a Florida seagrass community, which appear to be the most trophically complete food webs available in the primary literature due to their inclusion of autotrophs and empirically derived estimates of the relative energetic contributions of each trophic link. We show that most (54%) of the 212 species in the four food webs can be unambiguously assigned to a discrete trophic level. Omnivory among the remaining species appears to be quite limited, as judged by the standard deviation of omnivores' energy-weighted food-chain lengths. This allows simple algorithms based on binary food webs without energetic details to yield surprisingly accurate estimates of species' trophic and omnivory levels. While maximum trophic levels may plausibly exceed historically asserted limits, our analyses contradict both recent empirical claims that these limits are exceeded and recent theoretical claims that rampant omnivory eliminates the scientific utility of the trophic-level concept.
A unifying theory for top-heavy ecosystem structure in the ocean
DOI:10.1038/s41467-017-02450-y
URL
PMID:29295998
[本文引用: 1]
Size generally dictates metabolic requirements, trophic level, and consequently, ecosystem structure, where inefficient energy transfer leads to bottom-heavy ecosystem structure and biomass decreases as individual size (or trophic level) increases. However, many animals deviate from simple size-based predictions by either adopting generalist predatory behavior, or feeding lower in the trophic web than predicted from their size. Here we show that generalist predatory behavior and lower trophic feeding at large body size increase overall biomass and shift ecosystems from a bottom-heavy pyramid to a top-heavy hourglass shape, with the most biomass accounted for by the largest animals. These effects could be especially dramatic in the ocean, where primary producers are the smallest components of the ecosystem. This approach makes it possible to explore and predict, in the past and in the future, the structure of ocean ecosystems without biomass extraction and other impacts.
The complexity-stability relationship: Progress in mathematical models
复杂性-稳定性研究: 数学模型的进展
Specific non-monotonous interactions increase persistence of ecological networks
The compartmentation of real and assembled ecosystems
Body size and consumer-resource dynamics
Food-web dynamics under climate change

{kind=link}
{kind=link}
{kind=link}
{kind=link}