



互惠共生不仅是地球上所有生命形式的重要组成部分, 而且是陆地生态系统早期演化的基本驱动力。早在植物登陆之前, 地衣(真菌和藻类共生)就已经在陆地表面繁衍, 并大大加快了岩石风化形成土壤的速度。正如现代生态系统中地衣能够促进植物在原生演替中生长一样, 这种菌藻共生关系为地球早期植物的登陆和繁荣奠定了基础。
早期植物开始在陆地上生长时(~4.5亿年前), 面临着如何从土壤基质中吸收水分以及可溶性养分的挑战。植物通过与原始的丛枝菌根真菌形成互惠关系来克服这一困难, 即通过真菌菌丝从土壤中获取水分和养分(Strullu-Derrien et al, 2014)。真菌菌丝平均直径约为5 μm, 而现代植物的细根在全球尺度上平均约为300 μm (Ma et al, 2018)。这意味着在生物量相同的情况下, 这些植物共生者的搜索长度可达到植物根的3,600倍。由于共生菌根对植物资源供应的重要作用, 植物和真菌之间的这种战略合作对陆地系统早期森林的出现至关重要。
早期森林的繁荣给陆生植物带来了另一个挑战: 由于难以分解的木质素和纤维素的堆积, 越来越多的营养物质被锁在有机碎屑中。从二叠纪开始, 某些腐生菌类群通过分解有机质并释放锁定养分解决了这个问题(Floudas et al, 2012)。松科植物的早期物种与这些腐生菌(~1.4-1.8亿年前)的后代形成了战略互惠关系, 逐渐演变成了现代外生菌根植物。这种独特的互惠关系进一步增强了植物获取养分的能力, 并促进植物向新的生态位扩张, 如温带森林(Martin et al, 2016)。
现代热带雨林在大约6,500万年前出现, 由于较高的生产力, 它们对土壤养分有较高的需求, 这对热带雨林植物构成了新的独特的挑战。维管植物, 特别是豆科植物, 开始与能够固定大气氮的菌株形成稳定的互惠关系(Werner et al, 2015)。固氮植物的出现使植物能够将新的氮带入系统, 使生态系统免受氮的限制。
植物-微生物互惠共生关系虽然在古生物学文献中常常被忽视, 但综上可以看出, 它在地球历史上许多重要的里程碑事件里都起到了重要的推动作用。通过帮助植物克服资源的限制, 植物和微生物的互惠共生关系促进了植物生命向新的、更具挑战性的生态位扩展, 最终促成了我们目前观察到的植物、动物和栖息地的丰富多样性。
1 生物学和理论生态学领域的历史偏见
最明显的原因是达尔文进化论产生的文化惯性。达尔文这位生物学中最有影响力的人物将竞争的观点置于生物学中最有影响力的理论的中心位置(Darwin, 1859)。毫不夸张地说, 达尔文确立了竞争的主导地位, 认为竞争是生物学和生态学中最重要的驱动力。达尔文在他的《物种起源》中确实也注意到了互惠共生的重要性: “一朵花和一只蜜蜂可能会慢慢地, 同时地或相继地, 进化并以最完美的方式相互适应。而这种适应是基于个体产生对彼此有利的结构变异, 而使它们更好地生存”。这是他在整本书中对互惠关系最深的也是唯一的一次阐述。随后发展起来的与进化相关的理论(Wilson, 1975; Wilson & Sober, 1994)则更强调合作, 但从未获得与《物种起源》类似的影响力。
从更深层的角度看, 人们倾向于关注竞争而忽视互惠共生的原因植根于二者辩证组合的本质, 即组内合作、组间竞争。人们倾向认可团体之间的竞争, 因为它更明显, 往往更吸引人的关注。然而, 推动每一次竞争升级的隐藏力量却是集团内的合作, 但对局外者来说这并不容易观察到。这种情况在人类行为的进化中经常出现(Bowles & Gintis, 2011), 在植物-微生物系统中更是如此(例如, 观察两棵树的竞争比观察帮助它们竞争的共生体要容易得多)。
而在理论生态学中, 人们对互惠共生缺乏关注还有另一层原因, 即与Lotka-Volterra方程(以下称为L-V方程)的数学特性有关。对于理论生态学家来说, L-V方程对理解种群和群落生态学具有重要意义。我们最为熟知的L-V方程是用于描述捕食者-猎物动力学和种间资源竞争的。然而, 从L-V方程被提出的那一刻起, 它其实还可以描述合作和互惠共生(Lotka, 1925)。那么为什么L-V方程很少应用于植物-微生物互惠共生或合作的领域呢? 我们在这里使用L-V方程的一般形式(Lotka, 1925的现代版)作一说明:
其中, aii表示每个物种对自身的影响, aij 表示物种j对i的影响。一般而言, aii由于密度依赖性而呈负值。因此, 两个物种相互作用的动态是由 aij 的符号决定。具体来说, 我们可以把两种物种的相互作用分为以下几类: 竞争(a12, a21 < 0)、互利共生(a12, a21 > 0)、捕食或寄生(a12 < 0, a21 > 0)。May (1974)在其关于生态系统稳定性和复杂性的经典著作中指出, 互惠共生在理论生态学领域被忽视是因为历史原因, 而非生物学原因。更具体地说, 他认为早期建立的Lotka-Volterra系统难以维持互惠关系的稳定性。比较图2中的a图和b图可见, 参数的唯一区别是b图(无限种群)比a图有更强的互惠交互项。换句话说, 较强的互惠作用可引起种群的无限增长, 因此互惠关系成为了“导致群落动态不稳定的因素” (May, 1974)。
图1

图1
陆地生态系统中植物-微生物互惠共生的演化。a. 在登陆后不久, 陆地植物与内生菌根真菌(AMF)的祖先形成了共生关系(Strullu-Derrien et al, 2014)。图中展示了单个根尖(左, 浅绿色的表皮根细胞和浅棕色的皮层细胞)和AMF菌丝是如何形成细胞内结构的。b. 早期森林出现于木本维管植物占优势的早泥盆纪(Willis & McElwain, 2014)。c. 陆地植物与外生菌根真菌(EMF) (Cairney, 2000; Martin et al, 2016)建立稳定共生关系。EMF是能够分解木质素的腐生真菌的后代, 帮助植物获取有机质中被锁住的养分。d. 受环境条件变化的选择, 早白垩纪出现了有花植物, 是植物适应性创新的一个里程碑(Willis & McElwain, 2014)。e. 植物创新的另一个里程碑事件发生在地下,白垩纪-古近纪界线不久之后(Werner et al, 2015), 植物与固氮菌形成了互惠共生关系。这些细菌(居住在橙色的细胞中)可以打开氮气的三键, 为植物供应可利用的氮。共生关系a、c、e用红色箭头指向地质时间, 地质史关键事件b、d则使用蓝色箭头。该图修改自Lu和Hedin (2019)的
Fig. 1
Evolution of plant-microbe mutualism in terrestrial ecosystems. a. Land plants formed associations with early ancestors of arbuscular mycorrhizal fungi (AMF) soon after the plant’s colonization of terrestrial ecosystems (Strullu-Derrien et al, 2014). The schematic illustrates how an individual root tip (left, showing epidermal root cells in light green and cortical cells in light brown) and AMF mycelial forms intracellular structure. AMF hyphae is magnitude thinner than even the thinnest plant roots, allowing them superb ability to access soil resources from the porous soil matrix. b. The early forest emerged in the early Devonian after woody vascular plants gained dominance (Willis & McElwain, 2014). c. Land plants formed associations with ectomycorrhizal fungi (EMF) (Cairney, 2000), the descendant of wood-decaying fungi, aiding plants in accessing nutrients that otherwise would be locked into organic matter. d. Selected by the changing environmental condition, flowering plants emerged during the early Cretaceous as a milestone for plants’ adaptive innovation (Willis & McElwain, 2014). e. Another milestone for plant innovation happened belowground, shortly after the Cretaceous-Paleogene boundary Werner et al (2015), with plants forming mutualistic associations with nitrogen-fixing bacteria. These bacteria (housed in these orange cells) can breakdown the triple bond of N2 gas and supply plants with plant available forms of nitrogen. The geological timings of mutualistic relationships a, c, e are indicated by red arrows, while that of geological events b, d by blue arrows. This figure is modified based on
图2
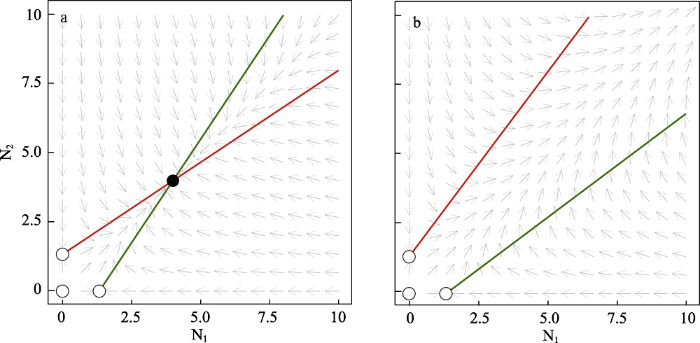
图2
互惠种群N1和N2的相平面。a. 当雅各比行列式det(J) > 0时, 相互作用导致稳定的非平凡平衡(r1 = r2 = 2, a11 = a22 = -1.5, a12 = a21 = 1)。b. 当雅各比行列式det(J) < 0时, 相互作用导致种群数量无限增长的非稳定动态(r1 = r2 = 2, a11 = a22 = -1.5, a12 = a21 = 2)。种群N1的零增长等值线用绿色, N2的零增长等值线用红色。黑色填充圆表示稳定平衡点, 白色填充圆表示非稳定和半稳定(鞍点)平衡点, 灰色箭头表示种群变化方向。绘图软件Julia 1.4.1。
Fig. 2
Phase planes of mutualistic interactions between population N1 and N2. a. The mutualistic interaction leads to stable nontrivial equilibrium when det(J) > 0 (r1 = r2 = 2, a11 = a22 = -1.5, a12 = a21 = 1). b. The mutualistic interaction leads to non-stable dynamics (infinite populations size) when det(J) < 0 (r1 = r2 = 2, a11 = a22 = -1.5, a12 = a21 = 2). The nullcline of population N1 is labeled in green and N2 in red. Black-filled circle denotes stable equilibrium while white-filled circles denote non-stable and half-stable (saddle points) equilibria. Plots are made in Julia 1.4.1.
此外, 植物的多个个体和多个共生体菌株之间的生物相互作用在自然界中几乎是普遍的(Moeller & Neubert, 2016), 而L-V方程最初被提出是来处理两两相互作用。虽然L-V框架可以很容易扩展到包含多个物种的相互作用(Goh, 1979; Neuhauser & Fargione, 2004), 但这会使它失去最初让它受欢迎的简洁性。
综上, 我们可以理解为什么在生物学文献中特别是在理论生态学(其中种群生物学是核心)中, 涉及到互惠关系的篇幅仍然相对不足。然而, 关于植物-微生物互惠的研究在两种不同视角下得到了迅速发展: 微生物学家聚焦于植物-微生物相互作用的机制, 而生态系统生态学家聚焦于植物-微生物相互作用的宏观影响。本文将概述这两种视角及其相关的数学工具(表1)。
表1 本综述中涉及的建模方法
Table 1
| 研究视角 Perspective | 建模方法 Modeling approach | 优点 Strength | 弱点 Weakness |
|---|---|---|---|
| 种群生物学 Population biology | L-V方程 Lotka-Volterra equations | 熟悉, 简洁 Familiarity and simplicity | 互惠导致种群不稳定性 Infinite population due to mutualism |
| 微生物生物学 Microbial biology | 迭代囚徒困境 Iterated Prisoner’s Dilemma | 简单, 通用性 Simplicity and generality | 缺乏种群动态, 缺乏伙伴选择, 对称设置 Lack of population dynamics, lack of partner choice, and symmetric setup |
| 生物市场理论 Biological market theory | 非对称设置, 伙伴选择 Asymmetry and partner choice | 各种数学工具的混合 Lack of simplicity, mixture of tools | |
| 生态系统生态学 Ecosystem ecology | 现象学 Phenomenology | 计算效率高 Computational efficiency | 原理机制不足 Not mechanistic |
| 优化 Optimization | 概念简单 Conceptual simplicity | 任意选择的目标函数 Arbitrary goal function | |
| 自适应动态 Adaptive dynamics | 可以模拟生物对变化的响应 Can capture biological adaptation | 计算成本高, 难扩展到大尺度模型中 Computationally costly to scale up |
2 微生物学家的观点: 关注微观机制
接下来要介绍的这个视角是一个多元化的研究领域, 关注的是植物和微生物相互作用的机理。具体来说, 来自这一视角的研究者更多地关注植物-微生物相互作用是如何出现(emergence)和维持的, 而较少关注这种相互作用对种群动态和生态系统过程的影响。以下简要介绍这一学派的建模方法, 主要关注博弈论方法(囚徒困境)和生物市场理论。
2.1 囚徒困境
自达尔文时代起, 互惠共生作用一直是一个令博物学家和科学家们困扰的难题。根据定义, 互惠共生包括相互作用的双方从它们的互惠互利者中获益(b), 同时因为提供服务而产生自身成本(c) (基于定义, b > c > 0)。进化生物学家会很自然地问一个简单的问题: 如果一个个体“决定”从互惠互利者那里获利却不付出成本去提供服务(成本 = 0), 将有何后果? 这种欺骗策略将享受更高的回报, 即(b-0) > (b-c), 并将随着时间的推移入侵互惠的种群。既然如此, 那为什么进化史保留了互惠共生呢?
我们可以用一个简单的理想化的二人博弈(表2)来表述这一悖论。在这个博弈中, 玩家可以选择两种策略: 合作和欺骗。这种特殊的设置在博弈论的文献中被称为“捐赠博弈” (Hilbe et al, 2013), 它是更为普遍的“囚徒困境” (Prisoner’s Dilemma, 简称PD)博弈的一种特殊情况。自从囚徒困境首次被引入科学文献, 它已经成为理解合作的一般动态的一个理想工具。在20世纪70年代, 它被引入生物学, 用来理解利他行为和互惠共生(Trivers, 1971)。
表2 囚徒困境博弈特殊情况下的收益矩阵。在每一轮游戏中, 每个玩家都可以选择合作或欺骗。合作的收益为b, 成本为c, 如果双方都合作, 双方都得到b-c的回报。如果双方都不合作, 回报是零。如果一方作弊, 另一方合作, 作弊者没有支付成本c就得到了效益b, 合作者支付成本c却没有获得效益b, 博弈的纳什均衡用粗体表示。
Table 2
| 玩家B (合作) Player B (Cooperate) | 玩家B (欺骗) Player B (Cheat) | |
|---|---|---|
| 玩家A (合作) Player A (Cooperate) | b-c; b-c | - c; b |
| 玩家A (欺骗) Player A (Cheat) | b; -c | 0; 0 |
基于我们对囚徒困境的理解, 上面提到的悖论具有一个纳什均衡(Nash equilibrium), 即(0, 0)。在这个均衡中, 任何一方都不能通过单方面改变策略来提高自己的适应度(fitness)。这个均衡告诉我们, 两个玩家都会欺骗, 并且只要满足b > c > 0, 欺骗就是一种进化稳定策略。很明显, 这个均衡并不是整个系统的最佳解决方案, 因为当把两个玩家的收益相加时, 2(b-c) > (b-c) > 0。从这个简单的模型中得出的定性结论显然与我们在自然界中观察到的情况不一致, 因为植物和微生物之间的互惠共生关系是数百万年来植物进化的基础, 并且在不同的群体中都有进化(图1)。这就提出了一个基本问题: 合作/互惠关系如何在囚徒困境框架内维持?
在囚徒困境框架中实现持续合作的一种方法是在游戏中进行多轮博弈, 每个玩家都能够“记住”上一轮所使用的策略并相应地改变自己的策略(迭代囚徒困境, Iterated Prisoner’s Dilemma, 简称IPD)。IPD是理解植物-微生物共生与合作的基础, 被广泛应用。下面我简要介绍IPD数学方法的要点。表2中收益矩阵的特殊情况可以推广如下: 可以定义Pk为给定个体k (k = A, B)的回报, 为收益b与成本c的差。收益b是对方j投资Ij的函数(k ≠ j), 成本c是自身(k)投资给对方Ik的函数:
在植物-微生物互惠系统中, A可以代表植物宿主, B代表与之共生的微生物, IA代表植物对共生生物的碳投资, IB代表共生体对寄主植物的养分供给。收益矩阵是由双方的投资组合(Ik, Ij)唯一决定的。IPD框架的核心是将时间维度添加到收益方程中:
其中t代表A和B之间进行的某一轮游戏(图3a)。下一轮每个玩家的投资$I_{t+1}^{k}$是前一轮游戏的函数:
图3
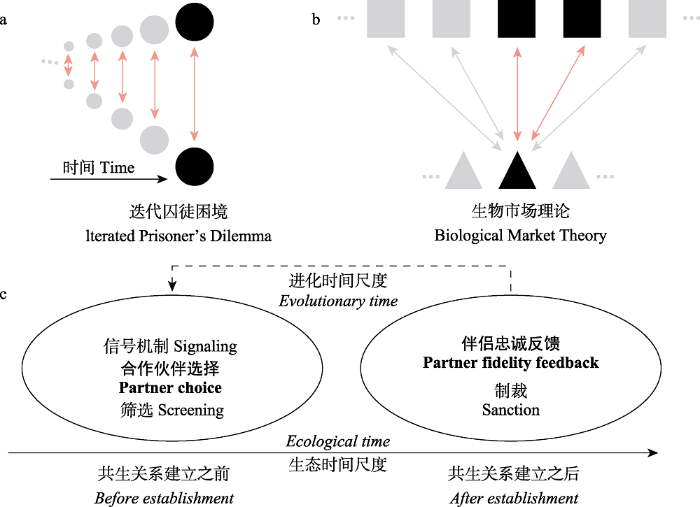
图3
迭代囚徒困境(IDP) (a)与生物市场理论(BMT) (b)的图形说明以及BMT的核心概念(c)。a. 两个玩家随着时间的推移以对称的方式重复互动, 每个玩家都能够记住最近一次的互动。玩家被标记为相同的填充点, 强调了这种方法固有的对称性(例如, 蝙蝠之间的种内合作)。b. 两组玩家通过类似于二分网络的形式进行交互作用, 其中一组中的每个玩家都可以与来自另一组的多个玩家进行交互(为了简化视觉, 这里只显示了部分交互)。这两个组在这里用不同的符号表示(正方形和三角形), 以强调这种方法内在的不对称性, 这使得它在处理特定种间的互惠互动(例如, 植物与微生物, 蜜蜂与花)时很有优势。BMT模型的最小设置用浅红色标记, 一共三个玩家, 其中一个玩家与另外两个玩家交互。c. 生物市场理论里面的最重要基本概念可以从生态时间尺度分成两类(实线时间轴): 合作伙伴选择和伴侣忠诚反馈。这两类概念所对应的生物学过程在进化时间尺度上是相连的(虚线)。
Fig. 3
Graphical illustration of Iterated Prisoner’s Dilemma (IDP) (a) vs. Biological Market Theory (BMT) (b), and core concepts used in BMT (c). a. Two players interact in a symmetric manner repeatedly over time, with each player being able to remember interactions from the immediate last time step. The players are denoted with the same filled dots, emphasizing the symmetry inherent to this approach (for example, intraspecific cooperation between individual bats). b. Two classes of players interact through a bipartite-graph alike interaction, where each player in one class can interact with multiple players from the other class (for visual simplicity, only part of the interactions are shown here). The two classes are denoted with different symbols here to emphasize the inherent asymmetry of this approach, which makes it unique in dealing with inter-specific mutualistic interaction (for example, plants vs. microbes, bees vs. flowers). The minimal setup of a BMT model is labeled in light red, where 1 player interacts with 2 other. c. The most fundamental concepts in BMT can be divided into two broad classes based on timescale (solid time arrow): partner choice vs. partner fidelity feedback. The biological processes represented by these two classes of concepts are linked over evolutionary time (dashed line).
函数f定义了玩家所采取的迭代策略, 例如著名的IPD策略“针锋相对” (tit-for-tat)可以表示为$I_{t+1}^{k}=I_{t}^{j}$(一般初始投资I0会选择合作)。Dobeli (1998)从植物-微生物互惠共生的角度提出了以下策略:
其中, 植物根据其先前的回报将其前一轮投资用回报率b来修正。如果b为正, 则意味着每一个互惠者(寄主植物和微生物共生体)只要之前的回报为正(来自交互作用的净收益), 就会增加投资。这是惩罚欺骗/寄生, 鼓励互惠, 从而维持合作的重要机制。
综上, 博弈论方法尽管构造非常简单, 却可以让我们获得强大的洞察力。然而博弈论方法尤其是应用最广泛的IPD方法, 存在一些关键的局限性:
首先, 自然界中的植物和微生物之间的碳-营养交换(投资组合) (Ik, Ij)是连续的, 形成一个从寄生到互惠的连续梯度, 但博弈论方法的默认设置是针对每个玩家的二元策略, 比如开或关。尽管后来的工作(从20世纪90年代后期开始)将二元策略扩展至连续策略, 但这种扩展常常不可避免地导致模型变得非常复杂, 而简单性恰恰是博弈论方法的优势所在。
其次, 在囚徒困境的框架下, 持续的互惠共生需要同一参与者之间的重复互动。在植物-微生物互惠共生中, 这意味着植物和微生物通过垂直传代, 以相同的基因型相互作用(后代树继承的微生物与母株相同)。但一般认为, 植物-微生物的互利共生更多地依赖水平传播, 很少通过垂直传播(Noë & Kiers, 2018)。
最后, 可能也是最重要的一点, 囚徒困境(以及其他游戏)的默认设置假设了交互类之间的对称关系。这种设置适用于解决种内合作问题, 但没有考虑到种间互惠共生固有的不对称性。例如, 豆科植物与根瘤菌的相互作用涉及一个寄主与多种菌株的相互作用(Simms & Taylor, 2002), 而植物与菌根的相互作用涉及多个寄主和共生体。
2.2 生物市场理论
由于囚徒困境模型的局限性, Noë在研究不对称求偶系统时提出了生物市场理论(Biological Market Theory, BMT) (Noë, 1990; Noë & Hammerstein, 1994)。该理论与IPD的关键区别是: 玩家可以根据服务质量的不同在多个玩家之间进行选择(因此BMT中玩家的最小数量为3, 如图3所示)。目前, BMT已经成为了解植物-微生物互惠共生及其对环境条件依赖性的最流行的工具。BMT的核心思想是, 互惠交换在本质上类似于人类社会中进行产品和服务交易的市场。BMT不是一种具体形式的数学方法, 而更多是一种基于一系列假设的概念框架。Ronald Noë和他的同事在他们的开创性论文中概述了这些假设(Noë & Hammerstein, 1994), 并在最近的一篇更具有生物学基础的综述中对这些假设做了更新(Noë & Kiers, 2018)。在具体的数学模型上, BMT可以呈现为非常不同的形式, 比如双人博弈、Tilman的资源竞争框架(de Mazancourt & Schwartz, 2010)等。
BMT的核心包括两个概念: 合作伙伴选择和伴侣忠实反馈(图3c)。合作伙伴选择指的是在寄主-共生体关系建立前每一方都可以在多个报价中进行选择, 评估预期回报并在最有利的回报中做出选择。但宿主或共生体如何在不执行合作关系之前就计算预期收益?这就需要双方通过一些信号来预测它们未来的收益, 比如使用化学混合物在根瘤菌和植物宿主之间传递信号。但这些预先建立的机制存在一个问题, 即作弊者可以侵入系统。伙伴忠实反馈是确保互惠关系稳定维持的一个重要机制, 即在合作关系开始执行后, 彼此确保对对方负责。其要点可以通过公式6来理解, 其中t+1时刻的相互投资取决于前一个时间步长的收益。最简单的情况是一个具有正反馈系数的线性响应(参见等式7)。例如, 如果根瘤菌表现良好, 宿主会通过优先的碳分配奖励它们(Bever, 2015); 而如果细菌作弊, 则宿主会通过给予较少的资源来惩罚共生体, 通常称为“制裁” (sanction)。
然而伴侣忠实反馈机制会产生试错成本。理想情况下, 伴侣会在不需要参与的情况下就预先甄别有可能作弊的一方。可以想象, 在进化过程中双方可以从它们建立合作后的表现中学习, 并将合作过程中的回报与某些信号建立相关函数, 从而进化出更加微妙的伴侣选择机制。随着时间的推移, 欺骗行为和对抗措施会不停出现, 就像一场进化的舞蹈。我们原本预期很强的伴侣选择机制会导致一个唯一赢家的出现以及随之而来的生物多样性的消失, 但正是这种进化上的“军备竞赛”维持了互惠共生关系在物种和群落层面的多样性。相比结构更简单的博弈论模型, 生物市场理论已经逐渐成为了学术界用来研究多样性如何维持以及多样性如何受到环境因素影响的有力工具。
2.3 未来的发展方向
任何学科的进步都需要两条腿走路: 经验认识的发展, 以及能够刻画经验认识的理论创新。以下三个具体方向可能会为微生物学家视角提供助益:
首先, 需要更好地理解植物-微生物相互作用的微尺度生理过程, 包括信号通路(Foo et al, 2014)、资源交换机制(Wipf et al, 2019)、相互作用网络性(Noë & Kiers, 2018)等。研究技术的发展无疑会推动对这些过程的理解。但如果能促进菌根真菌和固氮领域科研人员之间的交流合作, 将显著加快积累经验认识的速度。
其次, 人们越来越多地意识到, “囚徒困境博弈”框架在很多情形下可能并不是理解合作的最佳理论框架(Doebeli & Hauert, 2005)。例如, “雪堆博弈”是一种更加微妙的游戏, 基于该游戏无需多重迭代就可以维持合作。但目前将该模型框架应用于植物与微生物相互作用的研究还很少。
最后, 需要更强大的数学方法来整合不同类型模型的优点, 同时避免其固有的局限性。尽管还处于起步阶段, 但网络理论似乎是一个很有前途的方向, 因为它具有描述共生关系的天然复杂性的内在能力。菌根的相互作用网络和豆科植物-根瘤菌的相互作用网络都可以被理想化为类似于二分网络, 而这些交互网络所涉及的物种多样性(宿主层面和共生菌层面)是传统的数学方法无法描述的。在很长一段时间内, 理论生态学在处理生物多样性时的做法一直是把多样性分解为少量的功能属性, 然后再用能够描述这些功能属性的简单数学工具来分析(比如把成百上千种真菌统一看作一个共生菌类群, 然后在博弈模型中用一个玩家来描绘)。而网络模型的兴起让我们可以憧憬, 在不久的将来我们的理论工具可以直面物种多样性带来的复杂性。鉴于网络理论和工具的快速发展(Santos & Pacheco, 2005), 我们有理由相信在植物-微生物互惠共生的研究中发展网络分析有很好的前景(Montesinos-Navarro et al, 2012; Noe & Kiers, 2018)。
3 生态系统生态学家的视角: 关注大尺度的影响
与微生物学家的视角不同, 生态系统生态学家、生物地球化学和地球系统建模学者的关注点集中在群落、生态系统乃至整个生物圈的涌现特性上。历史上, 这些宏观尺度植物-微生物互惠共生的作用在很大程度上被忽视了。然而, 近来越来越多的证据表明植物-微生物的相互作用在塑造宏观尺度生态系统属性上具有关键作用(Averill et al, 2014; Terrer et al, 2016; Craig et al, 2018; Tedersoo et al, 2020), 因而植物-微生物互惠共生在宏观尺度上的定量分析正逐渐引起关注。
这些定量分析的核心是碳-营养元素交换的概念构架, 该构架将对共生体的碳投资视为成本, 将从共生体中获得的营养素视为收益。Vitousek和Field (1999)的工作可能是最早从生态系统角度模拟植物-微生物相互作用的尝试, 其中氮吸收途径成本被概念化并建立为数学模型。尽管它比较原始简单, 并且只关注了植物和根瘤菌的相互作用, 但它从生态系统生态学家和生物地球化学家的角度抓住了碳-营养交换的核心概念。该方案可以描述为以下一般形式:
其中g代表单个植物或整个植被的生长率(通常以净初级生产量的形式), xi是影响增长率的因素, $\theta $作为内在成本, 它表征了不同营养吸收途径j之间的成本差异, 如固氮和菌根营养吸收的成本不同。
尽管这一模型提供了刻画碳-营养元素交换过程的一般性理论构架, 但长期以来研究植物-菌根相互作用与植物-根瘤菌相互作用的生态学家一直缺乏交流。这种隔阂从美国生态学会年度会议上两个领域会议日程的重叠中就可以明显看出。我们必须把所有地下植物-微生物的互惠共生看作是一系列进化上相互关联的生物创新(图1), 并将其反映在一个整合的数学公式中。而一个整合的观点可以让我们的精力集中在碳和营养物质的通量交换上(从以植物为中心的角度), 并有利于我们探索植物-微生物互惠共生对生态系统属性的影响。下文将简要介绍植物-微生物共生的理论/计算方法, 以及在植被模式和生物地球化学循环中的应用。
3.1 植被模式
植被模式是植物在群落和生态系统尺度上所呈现出的空间和时间的规律性。很多研究表明, 植物和微生物之间的相互作用可促进群落水平植被模式的出现(Jiang et al, 2017; Bennett et al, 2017)。比如, 植物-真菌互惠共生影响群落构建(community assembly) (Taylor et al, 2019)、生态系统原生和次生演替早期阶段的群落更迭(Vitousek et al, 2002;Zheng et al, 2020), 以及在原始热带雨林持续被干扰和丢失养分的情况下雨林功能的维持(Batterman et al, 2013)。在更大尺度上, 植物-微生物共生在全球生态学研究中亦有重要作用。经验证据表明, 植物-微生物共生可影响全球植被的生物地理学模式, 比如植被分布的纬度格局与不同菌根类型的协同变化(Read, 1991; Read & Perez-Moreno, 2003)以及固氮植物在热带地区极高的丰富度(Jenny 1950; Hedin et al, 2009; Vitousek et al, 2002; Gentry, 1988)。然而, 尽管以往研究表明植物-微生物共生在理解生态系统大尺度时空格局中具重要意义, 但相关的数学建模却远远落后。一个关键限制是, 在生态系统尺度上仅仅考虑营养元素和碳的通量交换是不够的, 必须同时考虑植物-土壤反馈(养分循环)。
为什么整合植物-微生物互惠以及植物-土壤反馈如此重要呢?因为互惠共生关系本身会改变整个生态系统的营养状况(例如固氮情况下, 向系统中添加新的氮), 然后通过植物-土壤反馈传递到群落的动态, 因此最终的植被格局将取决于植物-微生物互惠共生与养分循环的相互作用。关于植物-土壤反馈的理论和概念的文献很早已出现(DeAngelis, 1992, Wilson & Agnew, 1992), 但对植物-土壤反馈和植物-微生物互惠共生的定量整合则要晚得多(Menge et al, 2009)。Menge等(2009)以生物地球化学领域的经典箱型模型(DeAngelis, 1992)为基础, 探讨了不同时间尺度上植物-根瘤菌互惠共生对生态系统营养限制的影响。在此基础上, 最新进展不仅整合了植物和根瘤菌的相互作用, 而且整合了在共生细菌和菌根真菌辅助下的整个植物营养策略(Lu & Hedin, 2019)。通过整合植物-土壤反馈和一系列营养互惠共生作用, 新模型能够解释和预测自然界中双稳态植被(见下文解释)的出现(Torti et al, 2001; Peh et al, 2011)。
丛枝菌根真菌(AMF)型植物主导的森林群落和外生菌根真菌(EMF)型植物主导的森林群落有时会同时出现在一个景观地带中, 但它们占据不同的斑块而不相互混合(图4a, b), 这导致AMF或EMF型植物丰度呈双峰分布(图4c, d)。Lu和Hedin (2019)提出的理论框架通过植物-土壤反馈和植物-微生物互惠共生的相互作用对这一看似奇怪的现象给出了直观解释。一方面, 菌根共生体的生理机制(图1)表明, AMF植物更依赖无机养分, 而EMF植物则依靠有机养分。另一方面, 最新研究对AMF植物和EMF植物如何影响植物-土壤养分反馈有了经验认识(Lin et al, 2017): EMF植物抑制矿化过程, 从而有利于有机养分的形成, 而AMF植物促进无机养分的产生。当这两个关键组成部分连接在一起时, 由于植物与菌根之间的正反馈作用, 出现了一种路径依赖的现象, 或者群落生态学中的“优先效应” (Schoener, 1976; Fukami & Nakajima, 2011)。具体地说, 当某种类型的植物群落形成后, 将进一步促进同一植物和菌根的建植, 最终产生对初始条件敏感的两种稳定状态(图4e)。
图4
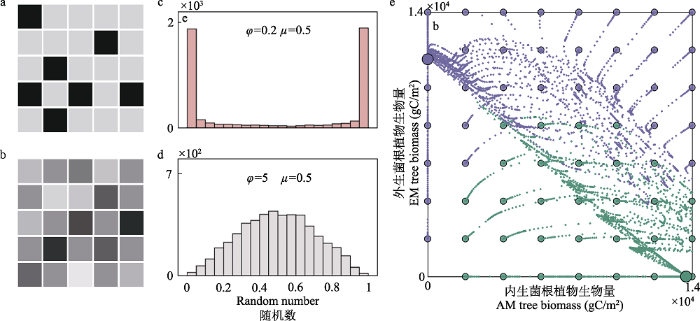
图4
双稳态植被格局及植物-微生物互惠共生的作用。a、b为植被斑块的景观图, 其中互惠共生类型A和B (在这种情况下, 丛枝菌根与外生菌根植物)的丰富度表示为色块的灰度。a所呈现的景观中, 双稳态植被状态是存在的, 一块植被要么是A主导要么是B主导, 而b则是两种互惠共生类型的随机混合。c、d互惠共生关系的分布可以用从beta分布中提取的5,000个随机数(代
Fig. 4
Bistable vegetation patterns and the role of plant-microbe mutualism. a, b. An illustration of a landscape with patches of vegetation, where the abundance of mutualistic interaction A and B (in this case, Arbuscular mycorrhizal vs. ectomycorrhizal symbioses) is denoted by the darkness of the gray hue. a presents a landscape where bistable vegetation states is found where you either find a patch of vegetation extremely high or extremely low in one type of mutualism, whereas b has a mixture of both mutualistic type in each patch. c, d. The distribution of mutualist abundance can be illustrated using the frequency distribution of 5,000 random numbers (representing 5,000 landscape patches) drawn from a beta distribution: $f(y:\mu, phi)=cy^{\mu\varphi-1} (1-y)^{(1-\mu)\varphi-1}$ (y indicates percentage of A, μ as the central tendency, and ϕ as the dispersion coefficient). e. Patches that have different founding composition (small dots) of A and B will over time diverge into two alternative stable states (larger dot, EMF indicated in purple and AMF indicated in green). Panels c, d, e are reproduced from figures published in Lu & Hedin (2019).
3.2 生物地球化学循环和全球模拟
生态系统的营养限制一直是生态系统生态学和生物地球化学研究的中心议题(Vitousek, 2004)。在全球变化的背景下, 这一议题变得越来越重要。最值得注意的是, 随着大气CO2浓度的增加, 植物对养分的需求不断上升, 而越来越多的养分被锁定在有机物(活着的和死去的)中, 陆地植被可能正在耗尽养分。这种渐进性养分限制的想法最初是针对氮提出的(Luo et al, 2004), 后来也扩展到其他主要土壤养分, 如磷(Ellsworth et al, 2017)。
如上所述, 从宏观角度来看, 基于碳和营养素交易构架的数学模型具有相同的成本效益结构核心。除了这个共享的核心之外, 还可以通过一系列不同的具体算法来模拟植物的实际生长过程, 从简单算法到复杂算法构成一个梯度(表1)。
在梯度的一端是一种现象学模型, 它描述了基于观察数据的生物学过程, 并常常为特定的生态系统制定特殊的参数。例如, MySCaN (Orwin et al, 2011)、ANAFORE (Deckmyn et al, 2011)和MYCOFON (Meyer et al, 2010)都可以有效地模拟植物-菌根关系对大尺度生态系统过程的作用。这种方法的优点是, 只要提供最新的数据其计算效率就非常高。但是, 这些模型非常依赖于特定的生态系统, 其中的功能关系是从特定的生态系统总结并派生出来的, 而并不能普遍适用于其他系统。
在梯度的中间, 研究人员假设植物像人类一样是理性的(借用了经济学的概念), 暗含植物“有意识地”优化某些指标的假设。这种优化方法理论上讲可以应用于更广泛的系统。例如, 可以假设植物从碳成本最低的途径吸收养分Fisher等(2010), 类似的技术可追溯到Vitousek和Field (1999)。这个算法在操作上是简单直接的, 但是我们从经验上知道, 植物本身有一个比这个简单假设更微妙的“算法”。尽管有大量更容易获取的可溶性土壤养分, 但固氮植物在施肥后仍从固氮根瘤获取氮(代价昂贵) (Menge et al, 2015)。这条研究路线后来的发展借鉴了物理学中的电阻概念, 将每条养分吸收通道的成本视为电阻, 将通过每条通道的营养物通量视为电流(Brzostek et al, 2014; Sulman et al, 2019)。植物的多种营养获取通道(根系本身、菌根、根瘤等)可以被视为并联的电路, 将欧姆定律应用于并联电路, 使得仿真更加简单, 在某种程度上也更加真实。
优化方法理论的局限性是目标函数选择具有任意性, 比如碳的增长量、植物的高度或者生物量。随着环境条件的急剧变化, 最适应者的标准可能会发生很大的变化。最高的、最重的或者碳含量最高的可能并不是存活得最好的: 说到底最适应环境变化的才是最能够存活下来的(Darwin, 1859)。因此, 在梯度最复杂的一端是自适应动力学方法(adaptive dynamics), 它能够描述植物适应变化环境的能力(Diekmann, 2003; Dercole & Rinaldi, 2008)。由于分析和计算的复杂性, 自适应动力学方法最近才被应用到植物-微生物互惠共生的领域, 且大多数工作都局限于相对简单的理论模型。例如, Menge等(2008)分析了生态系统中固氮策略的进化局限性。最近, Lu和Hedin (2019)考虑了植物-菌根和植物-根瘤菌的互惠共生, 并利用自适应性动力学方法, 仅仅基于非常简单的外部驱动就能够预测生境尺度上的植被模式和碳、氮、磷的循环。不过由于代码复杂性和计算能力的限制, 将这些理论模型扩展为更具有预测性的全球尺度的过程模型仍比较困难。
3.3 未来的发展方向
从前面介绍的模型的复杂性梯度来看, 越是贴近现实的算法越局限于简单的小型理论模型, 而大型地球系统模型仍然使用非常具有操作性的极为简单的算法。更进一步来说, 地球系统模型使用的功能型(functional type)是固定的, 根据收集的数据预先计算每个特定生态系统的参数。虽然这种方法从计算速度的角度来看是非常优越的, 但是也带来了巨大的挑战。由于算法的刚性, 大型系统模型的预测能力被限制在用来训练模型的数据范围内。
地球系统模型的进一步发展会逐渐引入更复杂更真实的算法, 这些算法通过更小的理论模型的探索变得更成熟。近年来, 新的地球系统模型包含了各种更加复杂的优化算法(Brzostek et al, 2014; Franklin et al, 2014; Sulman et al, 2019), 以及越来越现实的生物过程(Fisher et al, 2015; Fisher & Koven, 2020; Koven et al, 2020)。尽管自适应动力学方法固有的算法复杂, 我们仍可以期待它将成为地球系统建模工具箱中的主要工具之一。事实上, 目前已发展了基于自适应动力学的地球系统模型, 只不过模拟的是地表过程(Weng et al, 2017, 2019; Franklin et al, 2020)。笔者乐观地认为, 植物-微生物互惠共生将很快被纳入采用适应性动力学方法的大规模过程模型中。
4 总结
植物-微生物的互惠共生关系在不同时空尺度上对塑造生态系统中的关键过程起着至关重要的作用。然而, 从历史上看, 生物学和生态学长期忽视了这种控制机制而更关注竞争和寄生。希望本文能吸引读者更多关注互惠共生的重要性, 认识到共生与竞争是一个动态的相互依存的矛盾体, 应该作为一个整体来研究。因为这个矛盾体的两个组成部分(就像一个硬币的两面)一起推动了陆地生态系统的进化和现代生物圈的涌现。
参考文献
Mycorrhiza-mediated competition between plants and decomposers drives soil carbon storage
Key role of symbiotic dinitrogen fixation in tropical forest secondary succession
DOI:10.1038/nature12525
URL
PMID:24037375
[本文引用: 1]

Forests contribute a significant portion of the land carbon sink, but their ability to sequester CO2 may be constrained by nitrogen, a major plant-limiting nutrient. Many tropical forests possess tree species capable of fixing atmospheric dinitrogen (N2), but it is unclear whether this functional group can supply the nitrogen needed as forests recover from disturbance or previous land use, or expand in response to rising CO2 (refs 6, 8). Here we identify a powerful feedback mechanism in which N2 fixation can overcome ecosystem-scale deficiencies in nitrogen that emerge during periods of rapid biomass accumulation in tropical forests. Over a 300-year chronosequence in Panama, N2-fixing tree species accumulated carbon up to nine times faster per individual than their non-fixing neighbours (greatest difference in youngest forests), and showed species-specific differences in the amount and timing of fixation. As a result of fast growth and high fixation, fixers provided a large fraction of the nitrogen needed to support net forest growth (50,000 kg carbon per hectare) in the first 12 years. A key element of ecosystem functional diversity was ensured by the presence of different N2-fixing tree species across the entire forest age sequence. These findings show that symbiotic N2 fixation can have a central role in nitrogen cycling during tropical forest stand development, with potentially important implications for the ability of tropical forests to sequester CO2.
Plant-soil feedbacks and mycorrhizal type influence temperate forest population dynamics
DOI:10.1126/science.aai8212
URL
PMID:28082590
[本文引用: 1]
Feedback with soil biota is an important determinant of terrestrial plant diversity. However, the factors regulating plant-soil feedback, which varies from positive to negative among plant species, remain uncertain. In a large-scale study involving 55 species and 550 populations of North American trees, the type of mycorrhizal association explained much of the variation in plant-soil feedbacks. In soil collected beneath conspecifics, arbuscular mycorrhizal trees experienced negative feedback, whereas ectomycorrhizal trees displayed positive feedback. Additionally, arbuscular mycorrhizal trees exhibited strong conspecific inhibition at multiple spatial scales, whereas ectomycorrhizal trees exhibited conspecific facilitation locally and less severe conspecific inhibition regionally. These results suggest that mycorrhizal type, through effects on plant-soil feedbacks, could be an important contributor to population regulation and community structure in temperate forests.
Preferential allocation, physio-evolutionary feedbacks, and the stability and environmental patterns of mutualism between plants and their root symbionts
Modeling the carbon cost of plant nitrogen acquisition: Mycorrhizal trade-offs and multipath resistance uptake improve predictions of retranslocation
Evolution of mycorrhiza systems
DOI:10.1007/s001140050762
URL
PMID:11151665
[本文引用: 2]
Most terrestrial plants live in mutualistic symbiosis with root-infecting mycorrhizal fungi. Fossil records and molecular clock dating suggest that all extant land plants have arisen from an ancestral arbuscular mycorrhizal condition. Arbuscular mycorrhizas evolved concurrently with the first colonisation of land by plants some 450-500 million years ago and persist in most extant plant taxa. Ectomycorrhizas (about 200 million years ago) and ericoid mycorrhizas (about 100 million years ago) evolved subsequently as the organic matter content of some ancient soils increased and sclerophyllous vegetation arose as a response to nutrient-poor soils respectively. Mycorrhizal associations appear to be the result of relatively diffuse coevolutionary processes. While early events in the evolution of mycorrhizal symbioses may have involved reciprocal genetic changes in ancestral plants and free-living fungi, available evidence points largely to ongoing parallel evolution of the partners in response to environmental change.
Tree mycorrhizal type predicts within-site variability in the storage and distribution of soil organic matter
DOI:10.1111/gcb.14132
URL
PMID:29573504
[本文引用: 1]
Forest soils store large amounts of carbon (C) and nitrogen (N), yet how predicted shifts in forest composition will impact long-term C and N persistence remains poorly understood. A recent hypothesis predicts that soils under trees associated with arbuscular mycorrhizas (AM) store less C than soils dominated by trees associated with ectomycorrhizas (ECM), due to slower decomposition in ECM-dominated forests. However, an incipient hypothesis predicts that systems with rapid decomposition-e.g. most AM-dominated forests-enhance soil organic matter (SOM) stabilization by accelerating the production of microbial residues. To address these contrasting predictions, we quantified soil C and N to 1 m depth across gradients of ECM-dominance in three temperate forests. By focusing on sites where AM- and ECM-plants co-occur, our analysis controls for climatic factors that covary with mycorrhizal dominance across broad scales. We found that while ECM stands contain more SOM in topsoil, AM stands contain more SOM when subsoil to 1 m depth is included. Biomarkers and soil fractionations reveal that these patterns are driven by an accumulation of microbial residues in AM-dominated soils. Collectively, our results support emerging theory on SOM formation, demonstrate the importance of subsurface soils in mediating plant effects on soil C and N, and indicate that shifts in the mycorrhizal composition of temperate forests may alter the stabilization of SOM.
A resource ratio theory of cooperation
Dynamics of Nutrient Cycling and Food Webs
Simulating C cycles in forest soils: Including the active role of micro-organisms in the ANAFORE forest model
A beginner’s guide to adaptive dynamics
Models of cooperation based on the Prisoner’s Dilemma and the Snowdrift game
Elevated CO2 does not increase eucalypt forest productivity on a low-phosphorus soil
Taking off the training wheels: The properties of a dynamic vegetation model without climate envelopes, CLM4.5(ED)
Perspectives on the future of land surface models and the challenges of representing complex terrestrial systems
The Paleozoic origin of enzymatic lignin decomposition reconstructed from 31 fungal genomes
DOI:10.1126/science.1221748
URL
PMID:22745431
[本文引用: 1]
Wood is a major pool of organic carbon that is highly resistant to decay, owing largely to the presence of lignin. The only organisms capable of substantial lignin decay are white rot fungi in the Agaricomycetes, which also contains non-lignin-degrading brown rot and ectomycorrhizal species. Comparative analyses of 31 fungal genomes (12 generated for this study) suggest that lignin-degrading peroxidases expanded in the lineage leading to the ancestor of the Agaricomycetes, which is reconstructed as a white rot species, and then contracted in parallel lineages leading to brown rot and mycorrhizal species. Molecular clock analyses suggest that the origin of lignin degradation might have coincided with the sharp decrease in the rate of organic carbon burial around the end of the Carboniferous period.
Common and divergent roles of plant hormones in nodulation and arbuscular mycorrhizal symbioses
Organizing principles for vegetation dynamics
DOI:10.1038/s41477-020-0655-x
URL
PMID:32393882
[本文引用: 1]
Plants and vegetation play a critical-but largely unpredictable-role in global environmental changes due to the multitude of contributing processes at widely different spatial and temporal scales. In this Perspective, we explore approaches to master this complexity and improve our ability to predict vegetation dynamics by explicitly taking account of principles that constrain plant and ecosystem behaviour: natural selection, self-organization and entropy maximization. These ideas are increasingly being used in vegetation models, but we argue that their full potential has yet to be realized. We demonstrate the power of natural selection-based optimality principles to predict photosynthetic and carbon allocation responses to multiple environmental drivers, as well as how individual plasticity leads to the predictable self-organization of forest canopies. We show how models of natural selection acting on a few key traits can generate realistic plant communities and how entropy maximization can identify the most probable outcomes of community dynamics in space- and time-varying environments. Finally, we present a roadmap indicating how these principles could be combined in a new generation of models with stronger theoretical foundations and an improved capacity to predict complex vegetation responses to environmental change.
Forests trapped in nitrogen limitation―An ecological market perspective on ectomycorrhizal symbiosis
Community assembly: Alternative stable states or alternative transient states?
DOI:10.1111/j.1461-0248.2011.01663.x
URL
PMID:21790934
[本文引用: 1]
The concept of alternative stable states has long been a dominant framework for studying the influence of historical contingency in community assembly. This concept focuses on stable states, yet many real communities are kept in a transient state by disturbance, and the utility of predictions for stable states in explaining transient states remains unclear. Using a simple model of plant community assembly, we show that the conditions under which historical contingency affects community assembly can differ greatly for stable versus transient states. Differences arise because the contribution of such factors as mortality rate, environmental heterogeneity and plant-soil feedback to historical contingency changes as community assembly proceeds. We also show that transient states can last for a long time relative to immigration rate and generation time. These results argue for a conceptual shift of focus from alternative stable states to alternative transient states for understanding historical contingency in community assembly.
Changes in plant community diversity and floristic composition on environmental and geographical gradients
The nitrogen paradox in tropical forest ecosystems
Evolution of extortion in iterated prisoner’s dilemma games
Causes of the high nitrogen and organic matter content of certain tropical forest soils
Plant-mycorrhizal interactions mediate plant community coexistence by altering resource demand
Benchmarking and parameter sensitivity of physiological and vegetation dynamics using the Functionally Assembled Terrestrial Ecosystem Simulator (FATES) at Barro Colorado Island, Panama
Similar below-ground carbon cycling dynamics but contrasting modes of nitrogen cycling between arbuscular mycorrhizal and ectomycorrhizal forests
Global plant-symbiont organization and emergence of biogeochemical cycles resolved by evolution-based trait modelling
Progressive nitrogen limitation of ecosystem responses to rising atmospheric carbon dioxide
Evolutionary history resolves global organization of root functional traits
DOI:10.1038/nature25783
URL
PMID:29466331
[本文引用: 1]
Plant roots have greatly diversified in form and function since the emergence of the first land plants, but the global organization of functional traits in roots remains poorly understood. Here we analyse a global dataset of 10 functionally important root traits in metabolically active first-order roots, collected from 369 species distributed across the natural plant communities of 7 biomes. Our results identify a high degree of organization of root traits across species and biomes, and reveal a pattern that differs from expectations based on previous studies of leaf traits. Root diameter exerts the strongest influence on root trait variation across plant species, growth forms and biomes. Our analysis suggests that plants have evolved thinner roots since they first emerged in land ecosystems, which has enabled them to markedly improve their efficiency of soil exploration per unit of carbon invested and to reduce their dependence on symbiotic mycorrhizal fungi. We also found that diversity in root morphological traits is greatest in the tropics, where plant diversity is highest and many ancestral phylogenetic groups are preserved. Diversity in root morphology declines sharply across the sequence of tropical, temperate and desert biomes, presumably owing to changes in resource supply caused by seasonally inhospitable abiotic conditions. Our results suggest that root traits have evolved along a spectrum bounded by two contrasting strategies of root life: an ancestral 'conservative' strategy in which plants with thick roots depend on symbiosis with mycorrhizal fungi for soil resources and a more-derived 'opportunistic' strategy in which thin roots enable plants to more efficiently leverage photosynthetic carbon for soil exploration. These findings imply that innovations of belowground traits have had an important role in preparing plants to colonize new habitats, and in generating biodiversity within and across biomes.
Unearthing the roots of ectomycorrhizal symbioses
DOI:10.1038/nrmicro.2016.149
URL
PMID:27795567
[本文引用: 2]
During the diversification of Fungi and the rise of conifer-dominated and angiosperm- dominated forests, mutualistic symbioses developed between certain trees and ectomycorrhizal fungi that enabled these trees to colonize boreal and temperate regions. The evolutionary success of these symbioses is evident from phylogenomic analyses that suggest that ectomycorrhizal fungi have arisen in approximately 60 independent saprotrophic lineages, which has led to the wide range of ectomycorrhizal associations that exist today. In this Review, we discuss recent genomic studies that have revealed the adaptations that seem to be fundamental to the convergent evolution of ectomycorrhizal fungi, including the loss of some metabolic functions and the acquisition of effectors that facilitate mutualistic interactions with host plants. Finally, we consider how these insights can be integrated into a model of the development of ectomycorrhizal symbioses.
Evolutionary tradeoffs can select against nitrogen fixation and thereby maintain nitrogen limitation
Emergence and maintenance of nutrient limitation over multiple timescales in terrestrial ecosystems
Diversity of nitrogen fixation strategies in Mediterranean legumes
Simulating mycorrhiza contribution to forest C- and N cycling―The MYCOFON model
Multiple friends with benefits: An optimal mutualist management strategy?
The network structure of plant-arbuscular mycorrhizal fungi
A mutualism-parasitism continuum model and its application to plant-mycorrhizae interactions
Mycorrhizal markets, firms, and co-ops
A veto game played by baboons: A challenge to the use of the Prisoner’s Dilemma as a paradigm for reciprocity and cooperation
Biological markets: Supply and demand determine the effect of partner choice in cooperation, mutualism and mating
Organic nutrient uptake by mycorrhizal fungi enhances ecosystem carbon storage: A model-based assessment
DOI:10.1111/j.1461-0248.2011.01611.x
URL
PMID:21395963
[本文引用: 1]
Understanding the factors that drive soil carbon (C) accumulation is of fundamental importance given their potential to mitigate climate change. Much research has focused on the relationship between plant traits and C sequestration, but no studies to date have quantitatively considered traits of their mycorrhizal symbionts. Here, we use a modelling approach to assess the contribution of an important mycorrhizal fungal trait, organic nutrient uptake, to soil C accumulation. We show that organic nutrient uptake can significantly increase soil C storage, and that it has a greater effect under nutrient-limited conditions. The main mechanism behind this was an increase in plant C fixation and subsequent increased C inputs to soil through mycorrhizal fungi. Reduced decomposition due to increased nutrient limitation of saprotrophs also played a role. Our results indicate that direct uptake of nutrients from organic pools by mycorrhizal fungi could have a significant effect on ecosystem C cycling and storage.
Mechanisms of monodominance in diverse tropical tree-dominated systems
Mycorrhizas and nutrient cycling in ecosystems―A journey towards relevance?
Scale-free networks provide a unifying framework for the emergence of cooperation
Alternatives to Lotka-Volterra competition: Models of intermediate complexity
Partner choice in nitrogen-fixation mutualisms of legumes and rhizobia
DOI:10.1093/icb/42.2.369
URL
PMID:21708730
[本文引用: 1]
Mutualistic interactions are widespread and obligatory for many organisms, yet their evolutionary persistence in the face of cheating is theoretically puzzling. Nutrient-acquisition symbioses between plants and soil microbes are critically important to plant evolution and ecosystem function, yet we know almost nothing about the evolutionary dynamics and mechanisms of persistence of these ancient mutualisms. Partner-choice and partner-fidelity are mechanisms for dealing with cheaters, and can theoretically allow mutualisms to persist despite cheaters.Many models of cooperative behavior assume pairwise interactions, while most plant-microbe nutrient-acquisition symbioses involve a single plant interacting with numerous microbes. Market models, in contrast, are well suited to mutualisms in which single plants attempt to conduct mutually beneficial resource exchange with multiple individuals. Market models assume that one partner chooses to trade with a subset of individuals selected from a market of potential partners. Hence, determining whether partner-choice occurs in plant-microbe mutualisms is critical to understanding the evolutionary persistence and dynamics of these symbioses. The nitrogen-fixation/carbon-fixation mutualism between leguminous plants and rhizobial bacteria is widespread, ancient, and important for ecosystem function and human nutrition. It also involves single plants interacting simultaneously with several to many bacterial partners, including ineffective (
Fungal associations in Horneophyton ligneri from the Rhynie Chert (c. 407 million year old) closely resemble those in extant lower land plants: Novel insights into ancestral plant-fungus symbioses
Diverse mycorrhizal associations enhance terrestrial C storage in a global model
Successional dynamics of nitrogen fixation and forest growth in regenerating Costa Rican rainforests
DOI:10.1002/ecy.2637
URL
PMID:30698284
[本文引用: 1]
Regenerating tropical forests have an immense capacity to capture carbon and harbor biodiversity. The recuperation of the nitrogen cycle following disturbance can fuel biomass regeneration, but few studies have evaluated the successional dynamics of nitrogen and nitrogen inputs in tropical forests. We assessed symbiotic and asymbiotic nitrogen fixation, soil inorganic nitrogen concentrations, and tree growth in a well-studied series of five tropical forest plots ranging from 19 yr in age to old-growth forests. Wet-season soil inorganic nitrogen concentrations were high in all plots, peaking in the 29-yr-old plot. Inputs from symbiotic nitrogen fixation declined through succession, while asymbiotic nitrogen fixation peaked in the 37-yr-old plot. Consequently, the dominant nitrogen fixation input switched from symbiotic fixation in the younger plots to asymbiotic fixation in the older plots. Tree growth was highest in the youngest plots and declined through succession. Interestingly, symbiotic nitrogen fixation was negatively correlated with the basal area of nitrogen-fixing trees across our study plots, highlighting the danger in using nitrogen-fixing trees as a proxy for rates of symbiotic nitrogen fixation. Our results demonstrate that the nitrogen cycle has largely recuperated by 19 yr following disturbance, allowing for rapid biomass regeneration at our site. This work provides important insight into the sources and dynamics of nitrogen that support growth and carbon capture in regenerating Neotropical forests.
How mycorrhizal associations drive plant population and community biology
Mycorrhizal association as a primary control of the CO2 fertilization effect
Causes and consequences of monodominance in tropical lowland forests
DOI:10.1086/318629
URL
PMID:18707268
[本文引用: 1]
Tropical canopy dominance in lowland, well-drained forests by one plant species is a long-standing conundrum in tropical biology. Research now shows that dominance is not the result of one trait or mechanism. We suggest that the striking dominance of Gilbertiodendron dewevrei in the Ituri Forest of northeastern Congo is the result of a number of traits in adult trees that significantly modify the understory environment, making it difficult for other species to regenerate there. Adults cast deep shade that reduces light levels in the understory of the Gilbertiodendron forest to levels significantly lower than in the mixed-species forest. Moreover, the monodominant forest has deep leaf litter that could inhibit the establishment of small-seeded species, and the leaf litter is slow to decompose, potentially causing the low availability of nitrogen. We expect that juveniles of Gilbertiodendron may have an advantage in this environment over other species. In general, it appears that all tropical monodominant species share a similar suite of traits.
The Evolution of Reciprocal Altruism
Towards an ecological understanding of biological nitrogen fixation
Ecosystem constraints to symbiotic nitrogen fixers: A simple model and its implications
Predicting vegetation type through physiological and environmental interactions with leaf traits: Evergreen and deciduous forests in an earth system modeling framework
DOI:10.1111/gcb.13542
URL
PMID:27782353
[本文引用: 1]
Earth system models are incorporating plant trait diversity into their land components to better predict vegetation dynamics in a changing climate. However, extant plant trait distributions will not allow extrapolations to novel community assemblages in future climates, which will require a mechanistic understanding of the trade-offs that determine trait diversity. In this study, we show how physiological trade-offs involving leaf mass per unit area (LMA), leaf lifespan, leaf nitrogen, and leaf respiration may explain the distribution patterns of evergreen and deciduous trees in the temperate and boreal zones based on (1) an evolutionary analysis of a simple mathematical model and (2) simulation experiments of an individual-based dynamic vegetation model (i.e., LM3-PPA). The evolutionary analysis shows that these leaf traits set up a trade-off between carbon- and nitrogen-use efficiency at the scale of individual trees and therefore determine competitively dominant leaf strategies. As soil nitrogen availability increases, the dominant leaf strategy switches from one that is high in nitrogen-use efficiency to one that is high in carbon-use efficiency or, equivalently, from high-LMA/long-lived leaves (i.e., evergreen) to low-LMA/short-lived leaves (i.e., deciduous). In a region of intermediate soil nitrogen availability, the dominant leaf strategy may be either deciduous or evergreen depending on the initial conditions of plant trait abundance (i.e., founder controlled) due to feedbacks of leaf traits on soil nitrogen mineralization through litter quality. Simulated successional patterns by LM3-PPA from the leaf physiological trade-offs are consistent with observed successional dynamics of evergreen and deciduous forests at three sites spanning the temperate to boreal zones.
Competition alters predicted forest carbon cycle responses to nitrogen availability and elevated CO2: Simulations using an explicitly competitive, game-theoretic vegetation demographic model
DOI:10.5194/bg-16-4577-2019 URL [本文引用: 1]
Evolutionary signals of symbiotic persistence in the legume-rhizobia mutualism
A theory of group selection
Reintroducing group selection to the human behavioral sciences
DOI:10.1017/S0140525X00036104 URL [本文引用: 1]
Positive-feedback switches in plant communities
Trading on the arbuscular mycorrhiza market: From arbuscules to common mycorrhizal networks

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}