种子植物的花粉传递依赖不同的传粉媒介, 虫媒传粉的多样性、普遍性和高效性使其受到大量关注(黄双全和郭友好, 2000 ), 而水媒和风媒传粉往往被认为效率较低而在自然生态系统中处于劣势地位(Darwin, 1876 ; Faegri & van der Pijl, 1979 )。然而, 新的研究表明, 风媒传粉并不如想象中那么低效, 在某些环境条件或某些植物类群中, 其效率甚至高于虫媒传粉(Friedman & Barrett, 2009 )。目前, 被子植物中已经发现至少65次从动物媒向风媒的传粉转变(Linder, 1998 ; Friedman, 2011 )。传粉模式从虫媒转变为风媒, 相对普遍的原因可能在于植物只需要进化出相对简单的花部特征就可以适应风媒; 但从风媒转变为虫媒在自然界中却很少存在, 可能因为植物要重新获得与吸引、报酬和适应昆虫访花行为相关的复杂花部特征, 会经历更大的进化限制(Huang et al, 2013 )。因此, 通常认为从虫媒到风媒的传粉系统转变在某种程度上是不可逆的(Cox, 1991 ; Dodd et al, 1999 )。按照以上观点, 随着全球自然环境和动物传粉服务水平的双重变化(Hegland et al, 2009 ; Ollerton et al, 2014 ), 植物界中将极可能出现更高比例和多样化的风媒传粉现象。
传粉生物学研究中对风媒贡献的忽略往往源自对研究方法理解和运用上的一些误区。如某些植物被观察到了昆虫访问, 抑或其具有某些适应虫媒传粉的特征, 如散发气味、具有鲜艳颜色等, 都被认为是虫媒传粉的; 但最新研究表明, 具以上花部特征的如买麻藤属(Gnetum )和乌头属(Aconitum )植物, 均存在风媒传粉模式(Gong et al, 2016 ; Wang et al, 2016 )。此外, 与虫媒传粉研究相似, 短时的观察和片面的实验并不能得到准确全面的结论(龚燕兵和黄双全, 2007 ); 如静风条件下的风媒传粉实验, 往往会错误评估植物的风媒传粉效率(Niklas, 2015 )。这些研究方法上的缺陷, 极可能导致对植物风媒传粉存在性的不正确描述, 从而严重低估自然界中风媒传粉的普遍性和多样化的风媒传粉策略(Owens et al, 1998 )。基于以上背景, 本文总结并评价了风媒传粉的研究方法, 以期能对开展相关植物繁殖生态和保护研究的学者提供方法学上的参考。
1 风媒传粉植物的多样性
1.1 种类多样性
种子植物在不同选择压力下进化出多样化的传粉模式, 其中虫媒传粉植物的普遍性已受到大量关注, 而风媒传粉植物的种类多样性往往被低估。现存五大类种子植物——银杏(Ginkgo biloba )、松柏类(conifers)、苏铁类(cycads)、买麻藤目和被子植物中(Wang & Ran, 2014 ), 都已先后报道了不同比例风媒传粉模式的存在(Ackerman, 2000 ; Cully et al, 2002 )。
裸子植物被认为是风媒传粉起源的, 其中银杏和松柏类植物都是风媒传粉的(Owens et al, 1998 ; Jin et al, 2012 ); 苏铁类植物中虫媒和风媒传粉均能发挥作用(王乾等, 1997 ; Kono & Tobe, 2007 ); 买麻藤目中风媒是最主要的传粉方式, 但在各属中比例有所不同(Ickert-Bond & Renner, 2016 )。麻黄属(Ephedra )以风媒传粉为主(Bolinder et al, 2015 ), 而百岁兰(Welwitschia mirabilis )则主要为虫媒传粉(Wetschnig & Depisch, 1999 ), 早期研究认为买麻藤属是专性虫媒传粉的(Kato & Inoue, 1994 ), 但最新研究却揭示了其风媒传粉的存在(Gong et al, 2016 )。
对于虫媒传粉起源的被子植物, 风媒传粉被认为是从虫媒传粉转变而来的衍生模式, 业已发现被子植物的许多类群中风媒传粉是独立进化而来的(Friedman & Barrett, 2009 )。Ackerman (2000)的系统调查显示, 风媒传粉的被子植物分布在木兰亚纲、金缕梅亚纲、石竹亚纲、五桠果亚纲、蔷薇亚纲、菊亚纲、泽泻亚纲、棕榈亚纲、鸭跖草亚纲和百合亚纲中。在目的水平, 除鸭跖草亚纲中风媒传粉目的比例高达85%外, 其他各个亚纲都只有不到一半的目中出现了风媒传粉; 而以每目中风媒传粉科的数量做频率分布图, 则发现其符合对数正态分布规律, 即绝大部分的目中仅有0-2个科的植物是风媒传粉的, 比例最高的无患子目, 有高达5个科的植物具有风媒传粉模式。整体来讲, 被子植物中至少有18%的科和10%的种是风媒传粉的(Ackerman, 2000 )。
1.2 生境多样性
风媒传粉植物在地球上并不是随机分布的, 而是具有明显的地理和生态趋势(Whitehead, 1969 ; Regal, 1982 )。风媒花粉的成功传递不仅由风力决定, 许多生物与非生物因素都对风媒传粉有着重要影响, 如不同生境中植物群落结构的多样性、动物传粉服务的水平和稳定性、温湿度和降雨量等, 都能显著影响该生境中风媒传粉植物的比例(详见表1 和下文第3节)。
早期研究表明, 风媒传粉植物的比例往往随纬度和海拔的上升具有明显增加的趋势(Regal, 1982 )。温带地区植物居群相对聚集并且遮挡较少, 因而有利于花粉在空气中传播并成功传递到同种柱头。另外, 温带地区一般都具有严格的季节性, 相对于热带地区其植物生长季节较短, 因此很多树种通过早春开花以增加生长时间, 同时可避免与盛花期植物的传粉竞争。然而, 早春虫媒传粉服务相对缺乏且不稳定, 使得这些植物中很多依赖风媒传粉(Elzinga et al, 2007 )。正因如此, 温带的落叶森林、开阔草原、干旱荒漠及高山地区都易进化出风媒传粉物种(如彭德力等, 2012 )。Regal (1982)以北美木本植物为对象, 统计了不同生境中风媒传粉植物的比例, 发现不同群落间风媒传粉植物比例变化较大, 其中大部分温带森林中都有一半以上的种类是风媒传粉的。
物种丰富的热带地区降雨较多、湿度较大、树叶遮挡以及植物分散生长等都不利于风媒传粉, 这些因素在低地雨林中的影响尤为严重。然而, Vamosi等(2006)的大范围调查显示, 这些高生物多样性地区的植物因为虫媒传粉竞争而经历着更严重的花粉限制; 而花粉限制的选择压可能介导植物即使在这样不利的条件下也能进化出风媒传粉机制以寻求生殖保障(Cully et al, 2002 )。Bawa (1990)的早期调查显示, 热带低地雨林中也有1-4%的物种具风媒传粉模式。
2 风媒传粉的研究方法
2.1 花粉捕获法
探寻风媒传粉的可能性, 可首先使用花粉捕获装置(pollen traps)明确植株周围的空气中是否携有花粉; 通过收集目标植物的花粉作为参照, 根据花粉的大小、形状和表面纹饰等特征对捕获的花粉进行计数(如Gong et al, 2016 )。花粉捕获装置需悬挂于植株附近的适当距离, 尽量分多个方向散开, 在每个主方向上可设置距离梯度, 如此可量化风媒花粉的传播能力。根据花期、散粉和柱头可授情况, 可选择在白天或者晚上的合适时间段内悬挂装置。天气干燥有风时更易捕获风媒花粉, 但在某些不利条件如下雨或无风时使用该方法, 则可以对比评价不同环境条件下的风媒传粉效率。
捕获花粉最简单有效因而被广泛使用的装置是黏性载玻片(sticky slide), 即涂有凡士林、甘油或其他任何黏性物质的显微镜载玻片。黏性玻片可以通过多种形式安装以截获花粉, 如可平铺于培养皿、玻璃筒等器皿中, 再将器皿置于合适高度的标杆顶端, 如此便可收集由于重力作用垂直下落的风媒花粉; 也可将黏性玻片通过长尾夹和绳线垂直悬挂安置, 以捕获因风力而横向运动的花粉; 抑或模拟目标植物制作假花序, 作为底座大量安置玻片(Ackerman & Kevan, 2005 )。崔大练等(2008)在北重楼(Paris verticillata )开花植株的东南西北4个方位分别按照0.2、0.4、0.6和0.8 m的距离梯度安放涂有凡士林的玻片, 并以2小时为间隔回收玻片统计花粉捕获量, 从而得出该植物风媒花粉的传播能力和日散布规律。
使用以上方法也可能面临一些问题, 如雨天时玻片捕捉的花粉可能被雨水冲刷至黏性物质下层而难以计数, 又或直接被冲刷掉影响结果准确性。解决的办法是将玻片安放到顶端遮挡、但侧面敞开的简易装置中(如Durham sampler, 详见Ackerman & Kevan, 2005 )。此外, 野外条件下携有花粉的昆虫可能误碰玻片而影响实验结果。因此, 在显微计数时应特别注意一些有明显触碰痕迹、花粉分布极不均匀的玻片, 样本量充足时可考虑将其删除; 还可通过观察确定玻片未被昆虫触碰、对玻片进行套网或在昆虫不活跃期(如夜间)悬挂玻片(Dafni & Dukas, 1986 )等方法, 减少昆虫随机活动对风媒花粉捕获的干扰。
黏性玻片只能对空气中的花粉密度及传播距离进行相对定量测定, 精确测定需借助商业购买的空气颗粒取样器, 如英国Burkard Scientific公司的定量空气自动取样器和美国Sampling Technologies公司的Rotorod取样器。这些仪器都能在单位时间内吸取固定体积的空气从而实现花粉密度的精确测定。前者在测定10 μm以下花粉时效率较高, 而后者更适合测定大于10 μm的花粉(Frenz, 1999 )。
2.2 传粉者排除法
传粉者排除法(pollinator exclusion)是验证风媒传粉最直接有效的方法。一般来讲, 研究者需在植物开花前使用合适孔径的网袋, 套网隔绝潜在传粉者但允许风媒花粉进入, 将套网组(风媒)与自然对照组(风媒+虫媒)的坐果率和结籽率进行比较, 便能了解风媒传粉的生殖贡献情况(Kearns & Inouye, 1993 )。Culley等(2002)总结了使用该方法对风媒虫媒混合传粉系统(ambophily)植物的研究, 发现不同植物中, 风媒的传粉贡献比例变化较大(8-93%), 但大部分处于中间水平。在实验过程中, 以下三类因素的影响可能导致实验结果不理想或难以解释, 需在野外引起重视。
其一为网袋因素。一般的网袋都空间有限, 在野外使用过程中很易与植物繁殖器官接触而影响有性生殖过程。为避免这种不利影响, 需对套网处理进行“升级”。如若隔绝的传粉者为鸟类, 则仅需使用支撑较好的细铁丝网即可(如Fang et al, 2012 ); 如果使用尼龙网袋或纱袋隔绝传粉昆虫, 最好使用硬铁丝等制作支架支撑网袋, 并在封口处用绳线或细铁丝密封(见《生物多样性》2007年第15卷第6期封面)。此外, 网袋的表面积较大可能富集空气中的花粉, 使得套网组接受比自然组更多的风媒花粉; 相反的情况, 尽管网袋孔径大于花粉直径, 网袋在某种程度上仍有可能阻挡风媒花粉的自由通行。因此, 为验证套袋对花粉的富集或阻碍, 需在套袋内外设置黏性玻片比较花粉密度, 以判断是否受到该类因素的影响(Sacchi & Price, 1988 )。
其二为动物因素。套网往往无法隔绝某些小型昆虫的访问而影响实验结果。使用更小网孔的网袋或多层套网可以增加对小型昆虫访问的阻碍, 但也可能影响风媒花粉的进入。Baker和Cruden (1991)使用杀虫剂喷洒植物以驱除蓟马和蚜虫并取得了不错的效果。此外, 食花者(florivore)和食果者(frugivore)破坏花果可能影响有效数据的获取, 研究者应选择适当手段, 如增加套网时间等进行规避。
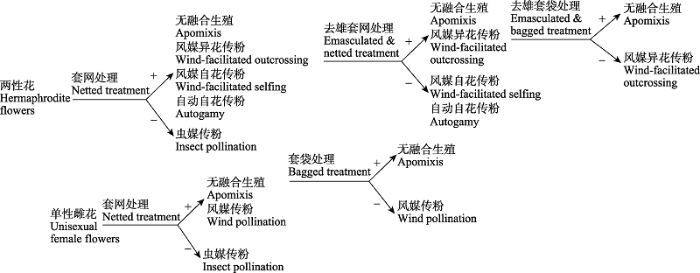
其三为植物因素。植物复杂的性系统(张大勇, 2004 )、交配系统(陈小勇, 2004 )和传粉系统(黄双全和郭友好, 2000 )可能增加实验结果解读的复杂性。研究者需在套网的基础上增加套袋(同时隔绝风媒和虫媒)和去雄处理并进行组合, 判断自动自交(autogamy)和无融合生殖(apomixis)在风媒传粉植物中发生的可能性(技术路线详见图1 )。
图1 传粉者排除法结合传粉处理检验风媒传粉的技术路线。+表示结籽率> 0, -表示结籽率 = 0。
Fig. 1 Technical routes to test wind pollination by pollinator exclusion and pollination treatments. + indicates seed set > 0, - indicates seed set = 0.
2.3 空气动力学
风媒传粉研究往往受诸多实际问题的限制, 如人力成本制约大量黏性玻片的花粉计数, 时间成本制约传粉者排除法的野外开展, 因为从套网处理到结实率数据获取往往需要等候太长时间。在这种情况下, 空气动力学则是一个很好的替代方法。研究者只需在野外快速收集研究物种的组织材料, 通过干燥或冷冻等方法保存后带回实验室, 开展后续实验即可。想在实验室内完全理解花粉在空气中的运动规律和实际效果是十分困难的, 因为太多的变量需要考虑(见Ackerman & Kevan, 2005 )。但就一般研究而言, 风媒传粉效率主要受花粉终端沉降速度(terminal settling velocity, U t )和周围气流加速场两个因素的影响(Niklas, 1985 )。U t 可用来推测植物风媒传粉的可能性, 风洞(wind tunnel)实验可以反映雌蕊周围气流加速场中花粉的真实运动情况, 而电脑模拟可以对其进行计算和评估(Niklas & Buchmann, 1987 ; Bolinder et al, 2015 )。
2.3.1 花粉沉降系数
U t 就是在静止的气流中, 花粉垂直下落达到稳定状态, 即重力和阻力平衡后的运动速度(Niklas & Spatz, 2012 ), 风媒传粉植物的U t 一般为2-6 cm/s (郭友好, 1994 )。密度低、体积小的花粉往往U t 较小, 因而具有更大的传播距离(Cresswell et al, 2010 )。以下介绍测量U t 的两种常用方法及使用中的问题和改进建议:
(1)频闪摄影法。包括黑箱子和玻璃圆筒法, 二者原理相似, 只是提供花粉在静止气流中下落的装置不同: 前者使用内壁涂有亚光材料以减少光散射的黑箱子(Niklas & Buchmann, 1987 ), 后者使用圆柱形的玻璃圆筒(Bolinder et al, 2015 )。测定技术均采用静止流体中花粉下落的频闪摄影技术。实验时需在黑暗的环境中放置好闪光灯和相机镜头, 当闪光灯和相机快门打开的同时释放花粉, 之后捕捉花粉的运动图像。使用匀速下落过程中两张连续图像的间距乘以闪光频率便可求得U t 。Bolinder等(2015)使用玻璃圆筒法测得Ephedra foeminea 的U t 在4.7- 21.44 cm/s之间, 这种大的差异是由花粉的不同聚集程度引起的。Hirose和Osada (2016)的玻璃圆筒法也大致类似, 只不过在玻璃圆筒底端安装发射波长为635 nm的激光装置, 替代闪光灯照明以协助花粉运动过程的拍摄; 他们研究的5种植物的U t 平均值分布在1.03-3.2 cm/s之间, 花粉越大的物种, U t 值越大。
(2)沉降塔法。沉降塔设备主要包括4个部分(详见Di-Giovanni et al, 1995 ; Borrell, 2012 ): 花粉释放装置、双层圆柱筒(提供花粉下落所需的静止空气环境)、转盘(其上装载径向排列的单面涂有凡士林的玻片, 并与花粉的释放同步进行)、控制面板(电动机供电, 可以控制圆盘转速)。实验中, 花粉垂直下落距离已知, 根据花粉在转盘玻片上的沉积位置和转盘转速来确定下落时间, 最终求得U t 。Borrell (2012)对Di-Giovanni等(1995)的沉降塔设备加以改进, 使用小型的振动刷代替原来的充气装置释放花粉, 对6种禾本科和4种雪松属(Cedrus )植物的U t 进行快速测定; 发现不同物种U t 差异显著, 禾本科植物的U t 平均值在3.81-5.29 cm/s之间, 显著低于雪松属植物的U t (6.72-9.18)。作者预测这种差异可以影响花粉的传播距离进而可能影响不同植物的遗传多样性和居群结构。
(3)问题和改进建议。为保证精确性, U t 的测定最好采用新鲜或者冻存保鲜的花粉(Karl Niklas, 私人联系)。但对于一些野外采集后难以保鲜的花粉, 终端沉降速度只能在野外实时测定。例如买麻藤(Gnetum montanum )的花药一旦离体或稍用力碰触便会氧化发黑, 即使迅速-20℃冻存也无法避免(朱亚如, 个人观察)。另外, 沉降塔设备在野外使用不便, Niklas和Buchmann (1987) 的黑箱子法更加便捷。但在实际使用过程中, 我们发现闪光灯的效果并不理想, 而强光手电的效果更佳。可预先在黑箱内壁制作一些刻度条背景, 便于记录花粉下落的距离。使用高清画质单反相机连续摄像, 记录花粉匀速下落的距离和时间, 即可求得U t 。
2.3.2 风洞实验
关于风洞实验, 其工作场的建立需满足以下3个条件: (1)配备变速风扇。Niklas和Buchmann (1987)使用变速范围为0.20-15.5 m/s的风扇; (2)工作场中气流的稳定性。风洞中放置充满氦气的中性漂浮球, 通过频闪照明观察球排出风洞的轨迹来判断(Niklas, 1984 )。因闪光频率已知, 该技术还可用来校准气流速度。实验中, 也可借助风速仪对气流速度进行周期性检测; (3)相对均匀的花粉浓度。实验中释放的花粉量可由体积判断, 释放点在风洞内部, 花粉浓度可通过连续监测穿过拍摄区的花粉数量判断。Niklas和Buchmann (1987)对2种麻黄(Ephedra trifurca 和E. nevadensis )的风洞实验结果显示, 某一特定的风速下, 植物在传粉过程中更易接受同种花粉。Cresswell等(2007)使用新鲜的花粉和雌球花材料, 在风速和花粉密度均稳定的风洞中, 对3种松科植物不同部位的花粉接受能力进行了比较, 结果发现3种植物的迎风面均能接受约70%的花粉, 背风面只能接受约2%的花粉, 而左右侧面接受的花粉比例在2种松属(Pinus )植物中无差异, 但在Cedrus libani 中却差异显著。
2.3.3 电脑模拟
基于风洞实验测定的不同物种花粉运动的差异, 可通过电脑模拟来评估导致这种空气动力学差异的物理基础(Niklas & Buchmann, 1987 ; Cresswell et al, 2007 )。电脑模拟前需获取的指标有: (1)花粉的密度、大小; (2)雌性生殖器官的大小、形态和普遍的朝向; (3)平均风速(Bolinder et al, 2015 )。花粉密度测定可使用密度梯度离心法(Forster & Flenley, 1993 ), 但如果U t 已知, 也可以采用以下公式:
其中r 是花粉半径, g是重力加速度, ρ 和ρ 0 分别是花粉和介质的密度, μ是空气的动力粘度系数, U t 是终端沉降速度(Niklas, 1985 ); 而雌性生殖器官的形态和大小可根据实际情况使用数码拍照、MicroCT扫描仪或扫描电镜测定(Cresswell et al, 2007 )。雌性生殖器官周围花粉行为的模拟详见Niklas和Kerchner (1986)设计的程序。Niklas和Buchmann (1987)使用电脑模拟E. trifurca 和E. nevadensis 胚珠周围气流场中花粉的运动行为, 得到了与风洞实验相似的结果; 并且预测, 不同物种花粉大小和密度的差异, 导致各物种花粉具有特定的惯性特征, 结合不同物种胚珠所形成的特异空气动力微环境, 即产生了风媒传粉过程中更易接受同种花粉的现象。Bolinder等(2015)的电脑模拟发现E. foeminea 不具备任何风媒花粉捕捉的功能特征, 这也与其虫媒传粉的早期观点一致。
3 风媒传粉影响因素的研究方法
传粉生物学的发展, 已从单纯描述传粉现象向揭示传粉机理和传粉选择作用下植物的适应性进化等方向发展。因此, 需对影响风媒传粉的因素进行研究, 以更深入理解风媒传粉发生的原因和后果。影响风媒的因素很多, 以下将其分为非生物和生物两大类, 就其部分重要因素进行论述, 更全面的因素总结见表1 。
3.1 非生物因素
研究非生物因素, 一般须在不同环境条件下, 在测量风媒传粉效率的基础上, 对某个或几个关注的非生物因素进行测量(见表1 ), 以寻找其中关联。风速、降雨量、温湿度等基础数据可从附近的气象站、野外工作台站等获取; 但如果想了解实验居群微环境中的相关数据, 则须针对性购买相关仪器进行测量。如我们购买的FODY TEMPUS Pro小型气象站, 能够连续监测大部分影响风媒传粉的非生物因素。
风速是影响风媒传粉的一个重要因素, 不但影响花粉的散播距离, 同时还影响柱头对花粉的捕捉(Whitehead, 1969 )。当风速大到可以克服花粉的U t , 但又不至于阻止柱头对花粉的接收时, 同种个体间的花粉传递效率最高(Niklas, 1985 ; Culley et al, 2002 )。鲁先文等(2008)使用风速计和黏性玻片法检验风速对沙棘(Hippophae rhamnoides )风媒传粉距离的影响, 发现风速较低时花粉传播距离一般不超过25 m; 但风速超过3.5 m/s时, 顺风花粉传播距离可高至85 m。Molina等(2001)使用Burkard定量空气自动取样器研究车前属(Plantago )植物花粉密度与环境因素的关系, 发现温度、湿度和风速对空气中的花粉密度均有显著影响。
高降雨量带来的湿度增加可影响花粉的聚集程度, 风媒花粉在潮湿空气中极易变黏而导致U t 增加; 湿度还能影响花药的脱水程度, 从且影响花粉的释放(Niklas, 1985 )。降雨最直接的影响就是“雨滴清除”现象, 即适合大小的雨滴可在较短时间内清除空气中所有的花粉(McDonald, 1962 )。Gong等(2016)在晴雨两种天气条件于小叶买麻藤(Gnetum parvifolium )雌雄株附近悬挂黏性玻片, 发现晴天时, 雄雌株附近玻片均能接收到花粉, 但雨天时雌株附近玻片上却无花粉捕获。
3.2 生物因素
花粉的大小、密度、形态和超微结构都能影响风媒传粉效率, 风媒花粉的大小一般在20-40 μm, 表面平滑干燥以减少花粉聚集(Ackerman & Kevan, 2005 ), 一些风媒物种的花粉壁内部存在大量空白区域, 有利于增加其浮力(Wodehouse, 1935 )。利用电镜技术就能清楚观察到花粉表面和内部结构差异(Tekleva, 2016 )。Bolinder等(2015)利用扫描和透射电镜技术对麻黄属植物的花粉进行研究, 发现花粉壁的超微结构与终端沉降速度是相关联的, 花粉壁顶盖下层空间内颗粒物质的相对比例和数量共同决定花粉密度, 并与整体的大小一起决定U t , 进而影响花粉散播能力。
而花柱的结构也十分重要, 花柱暴露性、柱头黏性和表面积的增加都有利于提高风媒花粉捕获效率, 如裸子植物能分泌传粉滴、禾本科植物具羽毛状或帚刷状柱头等(Faegri & van der Pijl, 1979 )。朱俊义等(2014)利用扫描电镜技术, 发现部分桦木科植物的花柱表皮细胞能形成乳突, 有利于增加花粉接受面积从而提高风媒传粉效率。
除以上花部构成因素外, 开花式样也能对风媒传粉产生重要影响(Friedman & Barrett, 2009 )。一般来讲, 风速随着高度的增加而增加, 对于风媒传粉的雌雄同株物种, 雄花处于较高位置更有利于花粉的散播(Bickel & Freeman, 1993 )。Niklas (1987)利用风洞实验, 发现紧凑和弥散型圆锥花序在花粉接受方面具二态性, 即紧凑型圆锥花序对气流的阻力较大, 所以花粉在花序的上风向处偏离, 而在下风向处通过沉降被捕捉; 而弥散型圆锥花序对气流的阻力较小, 因此大部分的花粉在上风向处与花序碰撞并被捕捉。Friedman和Harder (2004)采用花序捆绑固定法以减少花序的随风移动幅度, 发现该处理降低了紧凑型圆锥花序的花粉移出量, 但不影响柱头花粉接受量; 而在弥散型花序中, 花粉的移出量不受影响, 但柱头花粉接受量却显著降低。
4 结语
在自然界中, 仅根据风媒传粉综合征(wind pollination syndrome; 表1 ; Faegri & van der Pijl, 1979 )来判断风媒传粉的有无是不全面的, 有些植物即使不完全具备这些典型特征, 也可能具有风媒传粉模式(Gong et al, 2016 )。在全球气候变化、生境片断化、生物入侵等自然选择压力下, 植物某些细微特征的改变, 都有可能带来传粉模式的转变(Friedman, 2011 )。
相对于动物传粉研究, 目前我国风媒传粉研究所关注的植物种类仍十分有限。已发表的论文大多仅描述风媒植物的适应特征(如朱俊义等, 2014 ), 或使用野外实验方法研究风媒传粉的影响因素和生殖贡献(如鲁先文等, 2008 ; 崔大练等, 2008 ), 目前仍未见空气动力学方法在我国风媒传粉研究中的应用。此外, 仅有少量研究关注风媒与动物媒的并存现象及传粉转变的适应意义(Huang et al, 2013 ; Gong et al, 2016 ; Wang et al, 2016 )。
风媒传递的花粉往往体积较小而肉眼难辨, 相对于可直接观察的动物传粉过程, 其在野外研究中的隐蔽性更强。在野外实验中, 研究者应首选使用“花粉捕获法+传粉者排除法”的组合方法, 明确风媒传粉的可能性和生殖贡献大小。野外时间充足时, 应在各种环境条件下使用该组合方法对比测量风媒传粉效率。如野外条件受限, 研究者应快速收集相关植物组织材料, 在实验室内开展空气动力学研究。然而, 野外实验和室内方法并不能相互替代, 两类方法的综合对比运用才能由表及里地揭示自然传粉现象和发生机理(黄双全, 2012 )。同时应特别注意, 风媒和虫媒传粉可并存于一种植物, 此类研究应结合使用传粉昆虫行为的研究方法(龚燕兵和黄双全, 2007 )。本文就风媒传粉的研究方法进行了相对全面的总结和评价, 希望能对我国开展相关研究的学者有所裨益。
致谢:感谢郭友好教授和Karl J. Niklas教授提供的宝贵资料和建议!
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Ackerman JD 2000 ) Abiotic pollen and pollination: ecological, functional, and evolutionary perspectives
. Plant Systematics and Evolution , 222 , 167 -185 .
[本文引用: 2]
[2]
Ackerman JD Kevan PG 2005 ) Abiotic pollination
. In: Practical Pollination Biology (eds Dafni A, Kevan PG, Husband BC) , pp. 435 -480. Enviroquest Ltd., Cambridge.
[本文引用: 4]
[3]
Baker JD Cruden RW 1991 ) Thrips-mediated self-pollination of two facultatively xenogamous wetland species
. American Journal of Botany , 78 , 959 -963 .
[4]
Bawa KS 1990 ) Plant-pollinator interactions in tropical rain forests
. Annual Review of Ecology and Systematics , 21 , 399 -422 .
[5]
Bickel AM Freeman DC 1993 ) Effects of pollen vector and plant geometry on floral sex ratio in monoecious plants
. American Midland Naturalist , 130 , 239 -247 .
[本文引用: 1]
[6]
Bolinder K Niklas KJ Rydin C 2015 ) Aerodynamics and pollen ultrastructure in Ephedra
. American Journal of Botany , 102 , 457 -470 .
[本文引用: 4]
[7]
Borrell JS 2012 ) Rapid assessment protocol for pollen settling velocity: implications for habitat fragmentation
. Bioscience Horizons , 5 , 1 -9 .
[本文引用: 1]
[8]
Chen XY 2004 ) Mating systems
. In: Plant Life-History Evolution and Reproductive Ecology (ed. Zhang DY) , pp. 258 -279 . Science Press, Beijing. (in Chinese)
[本文引用: 1]
[陈小勇 (2004 ) 交配系统
. 见: 植物生活史进化与繁殖生态学 (张大勇主编) , 258 -279 页. 科学出版社 , 北京 .]
[本文引用: 1]
[9]
Cox PA 1991 ) Abiotic pollination: an evolutionary escape for animal-pollinated angiosperms
. Philosophical Transactions of the Royal Society B: Biological Sciences , 333 , 217 -224 .
[本文引用: 1]
[10]
Cresswell JE Henning K Pennel C Lahoubi M Patrick MA Young PG Tabor GR 2007 ) Conifer ovulate cones accumulate pollen principally by simple impaction
. Proceedings of the National Academy of Sciences, USA , 104 , 18141 -18144 .
[本文引用: 2]
[11]
Cresswell JE Krick J Patrick MA Lahoubi M 2010 ) The aerodynamics and efficiency of wind pollination in grasses
. Functional Ecology , 24 , 706 -713 .
[本文引用: 1]
[12]
Cui DL Man XL Ma YX Zhang YS 2008 ) Study on pollination ecology of Paris verticillata M.-Bieb. Acta Botanica Boreali-Occidentalia Sinica
, 28 , 298 -302 . (in Chinese with English abstract)
[本文引用: 1]
[崔大练 , 满秀玲 , 马玉心 , 张玉生 (2008 ) 北重楼传粉生态学研究
. 西北植物学报 , 28 , 298 -302 .]
[本文引用: 1]
[13]
Culley TM Weller SG Sakai AK 2002 ) The evolution of wind pollination in angiosperms
. Trends in Ecology & Evolution , 17 , 361 -369 .
[本文引用: 3]
[14]
Dafni A Dukas R 1986 ) Insect and wind pollination in Urginea maritima (Liliaceae)
. Plant Systematics and Evolution , 154 , 1 -10 .
[本文引用: 1]
[15]
Darwin C 1876 ) The Effects of Cross and Self Fertilisation in the Vegetable Kingdom
. John Murray, London .
[本文引用: 1]
[16]
Di-Giovanni F Kevan PG Nasr ME 1995 ) The variability in settling velocities of some pollen and spores
. Grana , 34 , 39 -44 .
[本文引用: 1]
[17]
Dodd ME Silvertown J Chase MW 1999 ) Phylogenetic analysis of trait evolution and species diversity variation among angiosperm families
. Evolution , 53 , 732 -744 .
[本文引用: 1]
[18]
Elzinga JA Atlan A Biere A Gigord L Weis AE Bernasconi G 2007 ) Time after time: flowering phenology and biotic interactions
. Trends in Ecology & Evolution , 22 , 432 -439 .
[本文引用: 1]
[19]
Faegri K van der Pilj L 1979 ) The Principles of Pollination Ecology, 3rd edn . Pergamon Press , Oxford .
[本文引用: 3]
[20]
Fang Q Chen YZ Huang SQ 2012 ) Generalist passerine pollination of a winter-flowering fruit tree in central China
. Annals of Botany , 109 , 379 -384 .
[本文引用: 1]
[21]
Forster M Flenley JR 1993 ) Pollen purification and fractionation by equilibrium density gradient centrifugation
. Palynology , 17 , 137 -155 .
[本文引用: 1]
[22]
Frenz DA 1999 ) Comparing pollen and spore counts collected with the Rotorod Sampler and Burkard spore trap
. Annals of Allergy, Asthma & Immunology , 83 , 341 -349 .
[本文引用: 1]
[23]
Friedman J 2011 ) Gone with the wind: understanding evolutionary transitions between wind and animal pollination in the angiosperms
. New Phytologist , 191 , 911 -913 .
[本文引用: 2]
[24]
Friedman J Barrett SCH 2009 ) Wind of change: new insights on the ecology and evolution of pollination and mating in wind-pollinated plants
. Annals of Botany , 103 , 1515 -1527 .
[本文引用: 3]
[25]
Friedman J Harder LD 2004 ) Inflorescence architecture and wind pollination in six grass species
. Functional Ecology , 18 , 851 -860 .
[26]
Gong YB Huang SQ 2007 ) On methodology of foraging behavior of pollinating insects
. Biodiversity Science , 15 , 576 -583 . (in Chinese with English abstract)
[本文引用: 2]
[龚燕兵 , 黄双全 (2007 ) 传粉昆虫行为的研究方法探讨
. 生物多样性 , 15 , 576 -583 .]
[本文引用: 2]
[27]
Gong YB Yang M Vamosi JC Yang HM Mu WX Li JK Wan T 2016 ) Wind or insect pollination? Ambophily in a subtropical gymnosperm Gnetum parvifolium (Gnetales)
. Plant Species Biology , 31 , 272 -279 .
[本文引用: 5]
[28]
Guo YH 1994 ) Pollination biology and the evolution of plants
. In: Plant Evolutionary Biology (eds Chen JK, Yang J) , pp. 232 -280 . Wuhan University Press, Wuhan. (in Chinese)
[本文引用: 1]
[郭友好 (1994 ) 传粉生物学与植物的进化
. 见: 植物进化生物学 (陈家宽, 杨继主编) , 232 -280 页. 武汉大学出版社 , 武汉 .]
[本文引用: 1]
[29]
Hegland SJ Nielsen A Lázaro A Bjerknes AL Totland Ø 2009 ) How does climate warming affect plant-pollinator interactions?
Ecology Letters , 12 , 184 -195 .
[本文引用: 1]
[30]
Hirose Y Osada K 2016 ) Terminal settling velocity and physical properties of pollen grains in still air
. Aerobiologia , 32 , 385 -394 .
[31]
Huang SQ 2012 ) Pollination biology in China in the 21st century: getting a good start
. Biodiversity Science , 20 , 239 -240 . (in Chinese)
[本文引用: 1]
[黄双全 (2012 ) 二十一世纪中国传粉生物学的研究: 良好的开端
. 生物多样性 , 20 , 239 -240 .]
[本文引用: 1]
[32]
Huang SQ Guo YH 2000 ) Advances in pollination biology
. Chinese Science Bulletin , 45 , 225 -237 . (in Chinese)
[本文引用: 2]
[黄双全 , 郭友好 (2000 ) 传粉生物学的研究进展
. 科学通报 , 45 , 225 -237 .]
[本文引用: 2]
[33]
Huang SQ Xiong YZ Barrett SCH 2013 ) Experimental evidence of insect pollination in Juncaceae, a primarily wind-pollinated family
. International Journal of Plant Sciences , 174 , 1219 -1228 .
[本文引用: 2]
[34]
Ickert-Bond SM Renner SS 2016 ) The Gnetales: recent insights on their morphology, reproductive biology, chromosome numbers, biogeography, and divergence times
. Journal of Systematics and Evolution , 54 , 1 -16 .
[本文引用: 1]
[35]
Jin B Zhang L Lu Y Wang D Jiang XX Zhang M Wang L 2012 ) The mechanism of pollination drop withdrawal in Ginkgo biloba L
. BMC Plant Biology , 12 , 59 .
[本文引用: 1]
[36]
Kato M Inoue T 1994 ) Origin of insect pollination
. Nature , 368 , 195 .
[本文引用: 1]
[37]
Kearns CA Inouye DW 1993 ) Techniques for Pollination Biologists
. University Press of Colorado, Niwot .
[本文引用: 1]
[38]
Kono M Tobe H 2007 ) Is Cycas revoluta (Cycadaceae) wind- or insect-pollinated?
American Journal of Botany , 94 , 847 -855 .
[本文引用: 1]
[39]
Linder HP 1998 ) Morphology and the evolution of wind pollination
. In: Reproductive Biology in Systematics, Conservation and Economic Botany (eds Owens SJ, Rudall PJ) , pp. 123 -135 . Royal Botanic Gardens, Kew.
[本文引用: 1]
[40]
Lu XW Ma RJ Sun K 2008 ) Determination of the wind pollination distances and flowering characteristics of Hippophae rhamnoides L. ssp. sinensis Rousi (Elaeagnaceae)
. Acta Ecologica Sinica , 28 , 2518 -2525 . (in Chinese with English abstract)
[本文引用: 1]
[鲁先文 , 马瑞君 , 孙坤 (2008 ) 中国沙棘(Hippophae rhamnoides L. ssp. sinensis Rousi)的开花特性及风媒传粉距离的检测
. 生态学报 , 28 , 2518 -2525 .]
[本文引用: 1]
[41]
McDonald JE 1962 ) Collection and washout of airborne pollens and spores by raindrops
. Science , 135 , 435 -437 .
[本文引用: 1]
[42]
Molina RT Palacios IS RodrÍguez AFM Muñoz JT Corchero AM 2001 ) Environmental factors affecting airborne pollen concentration in anemophilous species of Plantago
. Annals of Botany , 87 , 1 -8 .
[43]
Niklas KJ 1984 ) The motion of windborne pollen grains around conifer ovulate cones: implications on wind pollination
. American Journal of Botany , 71 , 356 -374 .
[本文引用: 1]
[44]
Niklas KJ 1985 ) The aerodynamics of wind pollination
. The Botanical Review , 51 , 328 -386 .
[本文引用: 4]
[45]
Niklas KJ 1987 ) Pollen capture and wind-induced movement of compact and diffuse grass panicles: implications for pollination efficiency
. American Journal of Botany , 74 , 74 -89 .
[本文引用: 1]
[46]
Niklas KJ 2015 ) A biophysical perspective on the pollination biology of Ephedra nevadensis and E. trifurca
. The Botanical Review , 81 , 28 -41 .
[本文引用: 1]
[47]
Niklas KJ Buchmann SL 1987 ) Aerodynamics of pollen capture in two sympatric Ephedra species
. Evolution , 41 , 104 -123 .
[本文引用: 3]
[48]
Niklas KJ Kerchner V 1986 ) Aerodynamics of Ephedra trifurca. II. Computer modelling of pollination efficiencies
. Journal of Mathematical Biology , 24 , 1 -24 .
[49]
Niklas KJ Spatz HC 2012 ) Plant Physics . University of Chicago Press , Chicago .
[本文引用: 1]
[50]
Ollerton J Erenler H Edwards M Crockett R 2014 ) Extinctions of aculeate pollinators in Britain and the role of large-scale agricultural changes
. Science , 346 , 1360 -1362 .
[本文引用: 1]
[51]
Owens JN Takaso T Runions CJ 1998 ) Pollination in conifers
. Trends in Plant Science , 3 , 479 -485 .
[本文引用: 2]
[52]
Peng DL Zhang ZQ Niu Y Yang Y Song B Sun H Li ZM 2012 ) Advances in the studies of reproductive strategies of alpine plants
. Biodiversity Science , 20 , 286 -299 . (in Chinese with English abstract)
[本文引用: 1]
[彭德力 , 张志强 , 牛洋 , 杨扬 , 宋波 , 孙航 , 李志敏 (2012 ) 高山植物繁殖策略的研究进展
. 生物多样性 , 20 , 286 -299 .]
[本文引用: 1]
[53]
Regal PJ 1982 ) Pollination by wind and animals: ecology of geographic patterns
. Annual Review of Ecology and Systematics , 13 , 497 -524 .
[本文引用: 2]
[54]
Sacchi CF Price PW 1988 ) Pollination of the arroyo willow, Salix lasiolepis: role of insects and wind
. American Journal of Botany , 75 , 1387 -1393 .
[本文引用: 1]
[55]
Tekleva M 2016 ) Pollen morphology and ultrastructure of several Gnetum species: an electron microscopic study
. Plant Systematics and Evolution , 302 , 291 -303 .
[本文引用: 1]
[56]
Vamosi JC Knight TM Steets JA Mazer SJ Burd M Ashman TL 2006 ) Pollination decays in biodiversity hotspots
. Proceedings of the National Academy of Sciences, USA , 103 , 956 -961 .
[57]
Wang LL Zhang C Yang ML Zhang GP Zhang ZQ Yang YP Duan YW 2016 ) Intensified wind pollination mediated by pollen dimorphism after range expansion in an ambophilous biennial Aconitum gymnandrum
. Ecology and Evolution , 7 , 541 -549 .
[本文引用: 2]
[58]
Wang Q Li CL Yang SY Huang R Chen FL 1997 ) Pollination biology of Cycas panzhihuaensis L. Zhou et S. Y. Yang
. Acta Botanica Sinica , 39 , 156 -163 . (in Chinese with English abstract)
[本文引用: 1]
[王乾 , 李朝銮 , 杨思源 , 黄荣 , 陈发林 (1997 ) 攀枝花苏铁传粉生物学研究
. 植物学报 , 39 , 156 -163 .]
[本文引用: 1]
[59]
Wang XQ Ran JH 2014 ) Evolution and biogeography of gymnosperms
. Molecular Phylogenetics and Evolution , 75 , 24 -40 .
[本文引用: 1]
[60]
Wetschnig W Depisch B 1999 ) Pollination biology of Welwitschia mirabilis HOOK. f. (Welwitschiaceae, Gnetopsida)
. Phyton , 39 , 167 -183 .
[本文引用: 1]
[61]
Whitehead DR 1969 ) Wind pollination in the angiosperms: evolutionary and environmental considerations
. Evolution , 23 , 28 -35 .
[本文引用: 2]
[62]
Wodehouse RP 1935 ) Pollen Grains: Their Structure, Identification and Significance in Science and Medicine
. McGraw- Hill, New York .
[本文引用: 1]
[63]
Zhang DY 2004 ) Plant Life-History Evolution and Reproductive Ecology . Science Press , Beijing . (in Chinese)
[本文引用: 1]
[张大勇 (2004 ) 植物生活史进化与繁殖生态学 . 科学出版社 , 北京 .]
[本文引用: 1]
[64]
Zhu JY Zhang LF Shen P Ren BQ Liang Y Chen ZD 2014 ) Wind pollination characteristics of styles in Betulaceae
. Chinese Bulletin of Botany , 49 , 524 -538 . (in Chinese with English abstract)
[本文引用: 1]
[朱俊义 , 张力凡 , 沈鹏 , 任保青 , 梁宇 , 陈之端 (2014 ) 桦木科植物花柱适应风媒传粉的特征
. 植物学报 , 49 , 524 -538 .]
[本文引用: 1]
Abiotic pollen and pollination: ecological, functional, and evolutionary perspectives
2
2000
... 种子植物在不同选择压力下进化出多样化的传粉模式, 其中虫媒传粉植物的普遍性已受到大量关注, 而风媒传粉植物的种类多样性往往被低估.现存五大类种子植物——银杏(Ginkgo biloba )、松柏类(conifers)、苏铁类(cycads)、买麻藤目和被子植物中(Wang & Ran, 2014 ), 都已先后报道了不同比例风媒传粉模式的存在(Ackerman, 2000 ; Cully et al, 2002 ). ...
... 对于虫媒传粉起源的被子植物, 风媒传粉被认为是从虫媒传粉转变而来的衍生模式, 业已发现被子植物的许多类群中风媒传粉是独立进化而来的(Friedman & Barrett, 2009 ).Ackerman (2000)的系统调查显示, 风媒传粉的被子植物分布在木兰亚纲、金缕梅亚纲、石竹亚纲、五桠果亚纲、蔷薇亚纲、菊亚纲、泽泻亚纲、棕榈亚纲、鸭跖草亚纲和百合亚纲中.在目的水平, 除鸭跖草亚纲中风媒传粉目的比例高达85%外, 其他各个亚纲都只有不到一半的目中出现了风媒传粉; 而以每目中风媒传粉科的数量做频率分布图, 则发现其符合对数正态分布规律, 即绝大部分的目中仅有0-2个科的植物是风媒传粉的, 比例最高的无患子目, 有高达5个科的植物具有风媒传粉模式.整体来讲, 被子植物中至少有18%的科和10%的种是风媒传粉的(Ackerman, 2000 ). ...
Abiotic pollination
4
2005
... 捕获花粉最简单有效因而被广泛使用的装置是黏性载玻片(sticky slide), 即涂有凡士林、甘油或其他任何黏性物质的显微镜载玻片.黏性玻片可以通过多种形式安装以截获花粉, 如可平铺于培养皿、玻璃筒等器皿中, 再将器皿置于合适高度的标杆顶端, 如此便可收集由于重力作用垂直下落的风媒花粉; 也可将黏性玻片通过长尾夹和绳线垂直悬挂安置, 以捕获因风力而横向运动的花粉; 抑或模拟目标植物制作假花序, 作为底座大量安置玻片(Ackerman & Kevan, 2005 ).崔大练等(2008)在北重楼(Paris verticillata )开花植株的东南西北4个方位分别按照0.2、0.4、0.6和0.8 m的距离梯度安放涂有凡士林的玻片, 并以2小时为间隔回收玻片统计花粉捕获量, 从而得出该植物风媒花粉的传播能力和日散布规律. ...
... 使用以上方法也可能面临一些问题, 如雨天时玻片捕捉的花粉可能被雨水冲刷至黏性物质下层而难以计数, 又或直接被冲刷掉影响结果准确性.解决的办法是将玻片安放到顶端遮挡、但侧面敞开的简易装置中(如Durham sampler, 详见Ackerman & Kevan, 2005 ).此外, 野外条件下携有花粉的昆虫可能误碰玻片而影响实验结果.因此, 在显微计数时应特别注意一些有明显触碰痕迹、花粉分布极不均匀的玻片, 样本量充足时可考虑将其删除; 还可通过观察确定玻片未被昆虫触碰、对玻片进行套网或在昆虫不活跃期(如夜间)悬挂玻片(Dafni & Dukas, 1986 )等方法, 减少昆虫随机活动对风媒花粉捕获的干扰. ...
... 风媒传粉研究往往受诸多实际问题的限制, 如人力成本制约大量黏性玻片的花粉计数, 时间成本制约传粉者排除法的野外开展, 因为从套网处理到结实率数据获取往往需要等候太长时间.在这种情况下, 空气动力学则是一个很好的替代方法.研究者只需在野外快速收集研究物种的组织材料, 通过干燥或冷冻等方法保存后带回实验室, 开展后续实验即可.想在实验室内完全理解花粉在空气中的运动规律和实际效果是十分困难的, 因为太多的变量需要考虑(见Ackerman & Kevan, 2005 ).但就一般研究而言, 风媒传粉效率主要受花粉终端沉降速度(terminal settling velocity, U t )和周围气流加速场两个因素的影响(Niklas, 1985 ).U t 可用来推测植物风媒传粉的可能性, 风洞(wind tunnel)实验可以反映雌蕊周围气流加速场中花粉的真实运动情况, 而电脑模拟可以对其进行计算和评估(Niklas & Buchmann, 1987 ; Bolinder et al, 2015 ). ...
... 花粉的大小、密度、形态和超微结构都能影响风媒传粉效率, 风媒花粉的大小一般在20-40 μm, 表面平滑干燥以减少花粉聚集(Ackerman & Kevan, 2005 ), 一些风媒物种的花粉壁内部存在大量空白区域, 有利于增加其浮力(Wodehouse, 1935 ).利用电镜技术就能清楚观察到花粉表面和内部结构差异(Tekleva, 2016 ).Bolinder等(2015)利用扫描和透射电镜技术对麻黄属植物的花粉进行研究, 发现花粉壁的超微结构与终端沉降速度是相关联的, 花粉壁顶盖下层空间内颗粒物质的相对比例和数量共同决定花粉密度, 并与整体的大小一起决定U t , 进而影响花粉散播能力. ...
Thrips-mediated self-pollination of two facultatively xenogamous wetland species
1991
Plant-pollinator interactions in tropical rain forests
1990
Effects of pollen vector and plant geometry on floral sex ratio in monoecious plants
1
1993
... 除以上花部构成因素外, 开花式样也能对风媒传粉产生重要影响(Friedman & Barrett, 2009 ).一般来讲, 风速随着高度的增加而增加, 对于风媒传粉的雌雄同株物种, 雄花处于较高位置更有利于花粉的散播(Bickel & Freeman, 1993 ).Niklas (1987)利用风洞实验, 发现紧凑和弥散型圆锥花序在花粉接受方面具二态性, 即紧凑型圆锥花序对气流的阻力较大, 所以花粉在花序的上风向处偏离, 而在下风向处通过沉降被捕捉; 而弥散型圆锥花序对气流的阻力较小, 因此大部分的花粉在上风向处与花序碰撞并被捕捉.Friedman和Harder (2004)采用花序捆绑固定法以减少花序的随风移动幅度, 发现该处理降低了紧凑型圆锥花序的花粉移出量, 但不影响柱头花粉接受量; 而在弥散型花序中, 花粉的移出量不受影响, 但柱头花粉接受量却显著降低. ...
Aerodynamics and pollen ultrastructure in Ephedra
4
2015
... 裸子植物被认为是风媒传粉起源的, 其中银杏和松柏类植物都是风媒传粉的(Owens et al, 1998 ; Jin et al, 2012 ); 苏铁类植物中虫媒和风媒传粉均能发挥作用(王乾等, 1997 ; Kono & Tobe, 2007 ); 买麻藤目中风媒是最主要的传粉方式, 但在各属中比例有所不同(Ickert-Bond & Renner, 2016 ).麻黄属(Ephedra )以风媒传粉为主(Bolinder et al, 2015 ), 而百岁兰(Welwitschia mirabilis )则主要为虫媒传粉(Wetschnig & Depisch, 1999 ), 早期研究认为买麻藤属是专性虫媒传粉的(Kato & Inoue, 1994 ), 但最新研究却揭示了其风媒传粉的存在(Gong et al, 2016 ). ...
... 风媒传粉研究往往受诸多实际问题的限制, 如人力成本制约大量黏性玻片的花粉计数, 时间成本制约传粉者排除法的野外开展, 因为从套网处理到结实率数据获取往往需要等候太长时间.在这种情况下, 空气动力学则是一个很好的替代方法.研究者只需在野外快速收集研究物种的组织材料, 通过干燥或冷冻等方法保存后带回实验室, 开展后续实验即可.想在实验室内完全理解花粉在空气中的运动规律和实际效果是十分困难的, 因为太多的变量需要考虑(见Ackerman & Kevan, 2005 ).但就一般研究而言, 风媒传粉效率主要受花粉终端沉降速度(terminal settling velocity, U t )和周围气流加速场两个因素的影响(Niklas, 1985 ).U t 可用来推测植物风媒传粉的可能性, 风洞(wind tunnel)实验可以反映雌蕊周围气流加速场中花粉的真实运动情况, 而电脑模拟可以对其进行计算和评估(Niklas & Buchmann, 1987 ; Bolinder et al, 2015 ). ...
... (1)频闪摄影法.包括黑箱子和玻璃圆筒法, 二者原理相似, 只是提供花粉在静止气流中下落的装置不同: 前者使用内壁涂有亚光材料以减少光散射的黑箱子(Niklas & Buchmann, 1987 ), 后者使用圆柱形的玻璃圆筒(Bolinder et al, 2015 ).测定技术均采用静止流体中花粉下落的频闪摄影技术.实验时需在黑暗的环境中放置好闪光灯和相机镜头, 当闪光灯和相机快门打开的同时释放花粉, 之后捕捉花粉的运动图像.使用匀速下落过程中两张连续图像的间距乘以闪光频率便可求得U t .Bolinder等(2015)使用玻璃圆筒法测得Ephedra foeminea 的U t 在4.7- 21.44 cm/s之间, 这种大的差异是由花粉的不同聚集程度引起的.Hirose和Osada (2016)的玻璃圆筒法也大致类似, 只不过在玻璃圆筒底端安装发射波长为635 nm的激光装置, 替代闪光灯照明以协助花粉运动过程的拍摄; 他们研究的5种植物的U t 平均值分布在1.03-3.2 cm/s之间, 花粉越大的物种, U t 值越大. ...
... 基于风洞实验测定的不同物种花粉运动的差异, 可通过电脑模拟来评估导致这种空气动力学差异的物理基础(Niklas & Buchmann, 1987 ; Cresswell et al, 2007 ).电脑模拟前需获取的指标有: (1)花粉的密度、大小; (2)雌性生殖器官的大小、形态和普遍的朝向; (3)平均风速(Bolinder et al, 2015 ).花粉密度测定可使用密度梯度离心法(Forster & Flenley, 1993 ), 但如果U t 已知, 也可以采用以下公式: ...
Rapid assessment protocol for pollen settling velocity: implications for habitat fragmentation
1
2012
... (2)沉降塔法.沉降塔设备主要包括4个部分(详见Di-Giovanni et al, 1995 ; Borrell, 2012 ): 花粉释放装置、双层圆柱筒(提供花粉下落所需的静止空气环境)、转盘(其上装载径向排列的单面涂有凡士林的玻片, 并与花粉的释放同步进行)、控制面板(电动机供电, 可以控制圆盘转速).实验中, 花粉垂直下落距离已知, 根据花粉在转盘玻片上的沉积位置和转盘转速来确定下落时间, 最终求得U t .Borrell (2012)对Di-Giovanni等(1995)的沉降塔设备加以改进, 使用小型的振动刷代替原来的充气装置释放花粉, 对6种禾本科和4种雪松属(Cedrus )植物的U t 进行快速测定; 发现不同物种U t 差异显著, 禾本科植物的U t 平均值在3.81-5.29 cm/s之间, 显著低于雪松属植物的U t (6.72-9.18).作者预测这种差异可以影响花粉的传播距离进而可能影响不同植物的遗传多样性和居群结构. ...
交配系统
1
2004
... 其三为植物因素.植物复杂的性系统(张大勇, 2004 )、交配系统(陈小勇, 2004 )和传粉系统(黄双全和郭友好, 2000 )可能增加实验结果解读的复杂性.研究者需在套网的基础上增加套袋(同时隔绝风媒和虫媒)和去雄处理并进行组合, 判断自动自交(autogamy)和无融合生殖(apomixis)在风媒传粉植物中发生的可能性(技术路线详见图1 ). ...
交配系统
1
2004
... 其三为植物因素.植物复杂的性系统(张大勇, 2004 )、交配系统(陈小勇, 2004 )和传粉系统(黄双全和郭友好, 2000 )可能增加实验结果解读的复杂性.研究者需在套网的基础上增加套袋(同时隔绝风媒和虫媒)和去雄处理并进行组合, 判断自动自交(autogamy)和无融合生殖(apomixis)在风媒传粉植物中发生的可能性(技术路线详见图1 ). ...
Abiotic pollination: an evolutionary escape for animal-pollinated angiosperms
1
1991
... 种子植物的花粉传递依赖不同的传粉媒介, 虫媒传粉的多样性、普遍性和高效性使其受到大量关注(黄双全和郭友好, 2000 ), 而水媒和风媒传粉往往被认为效率较低而在自然生态系统中处于劣势地位(Darwin, 1876 ; Faegri & van der Pijl, 1979 ).然而, 新的研究表明, 风媒传粉并不如想象中那么低效, 在某些环境条件或某些植物类群中, 其效率甚至高于虫媒传粉(Friedman & Barrett, 2009 ).目前, 被子植物中已经发现至少65次从动物媒向风媒的传粉转变(Linder, 1998 ; Friedman, 2011 ).传粉模式从虫媒转变为风媒, 相对普遍的原因可能在于植物只需要进化出相对简单的花部特征就可以适应风媒; 但从风媒转变为虫媒在自然界中却很少存在, 可能因为植物要重新获得与吸引、报酬和适应昆虫访花行为相关的复杂花部特征, 会经历更大的进化限制(Huang et al, 2013 ).因此, 通常认为从虫媒到风媒的传粉系统转变在某种程度上是不可逆的(Cox, 1991 ; Dodd et al, 1999 ).按照以上观点, 随着全球自然环境和动物传粉服务水平的双重变化(Hegland et al, 2009 ; Ollerton et al, 2014 ), 植物界中将极可能出现更高比例和多样化的风媒传粉现象. ...
Conifer ovulate cones accumulate pollen principally by simple impaction
2
2007
... 基于风洞实验测定的不同物种花粉运动的差异, 可通过电脑模拟来评估导致这种空气动力学差异的物理基础(Niklas & Buchmann, 1987 ; Cresswell et al, 2007 ).电脑模拟前需获取的指标有: (1)花粉的密度、大小; (2)雌性生殖器官的大小、形态和普遍的朝向; (3)平均风速(Bolinder et al, 2015 ).花粉密度测定可使用密度梯度离心法(Forster & Flenley, 1993 ), 但如果U t 已知, 也可以采用以下公式: ...
... 其中r 是花粉半径, g是重力加速度, ρ 和ρ 0 分别是花粉和介质的密度, μ是空气的动力粘度系数, U t 是终端沉降速度(Niklas, 1985 ); 而雌性生殖器官的形态和大小可根据实际情况使用数码拍照、MicroCT扫描仪或扫描电镜测定(Cresswell et al, 2007 ).雌性生殖器官周围花粉行为的模拟详见Niklas和Kerchner (1986)设计的程序.Niklas和Buchmann (1987)使用电脑模拟E. trifurca 和E. nevadensis 胚珠周围气流场中花粉的运动行为, 得到了与风洞实验相似的结果; 并且预测, 不同物种花粉大小和密度的差异, 导致各物种花粉具有特定的惯性特征, 结合不同物种胚珠所形成的特异空气动力微环境, 即产生了风媒传粉过程中更易接受同种花粉的现象.Bolinder等(2015)的电脑模拟发现E. foeminea 不具备任何风媒花粉捕捉的功能特征, 这也与其虫媒传粉的早期观点一致. ...
The aerodynamics and efficiency of wind pollination in grasses
1
2010
... U t 就是在静止的气流中, 花粉垂直下落达到稳定状态, 即重力和阻力平衡后的运动速度(Niklas & Spatz, 2012 ), 风媒传粉植物的U t 一般为2-6 cm/s (郭友好, 1994 ).密度低、体积小的花粉往往U t 较小, 因而具有更大的传播距离(Cresswell et al, 2010 ).以下介绍测量U t 的两种常用方法及使用中的问题和改进建议: ...
北重楼传粉生态学研究
1
2008
... 相对于动物传粉研究, 目前我国风媒传粉研究所关注的植物种类仍十分有限.已发表的论文大多仅描述风媒植物的适应特征(如朱俊义等, 2014 ), 或使用野外实验方法研究风媒传粉的影响因素和生殖贡献(如鲁先文等, 2008 ; 崔大练等, 2008 ), 目前仍未见空气动力学方法在我国风媒传粉研究中的应用.此外, 仅有少量研究关注风媒与动物媒的并存现象及传粉转变的适应意义(Huang et al, 2013 ; Gong et al, 2016 ; Wang et al, 2016 ). ...
北重楼传粉生态学研究
1
2008
... 相对于动物传粉研究, 目前我国风媒传粉研究所关注的植物种类仍十分有限.已发表的论文大多仅描述风媒植物的适应特征(如朱俊义等, 2014 ), 或使用野外实验方法研究风媒传粉的影响因素和生殖贡献(如鲁先文等, 2008 ; 崔大练等, 2008 ), 目前仍未见空气动力学方法在我国风媒传粉研究中的应用.此外, 仅有少量研究关注风媒与动物媒的并存现象及传粉转变的适应意义(Huang et al, 2013 ; Gong et al, 2016 ; Wang et al, 2016 ). ...
The evolution of wind pollination in angiosperms
3
2002
... 种子植物在不同选择压力下进化出多样化的传粉模式, 其中虫媒传粉植物的普遍性已受到大量关注, 而风媒传粉植物的种类多样性往往被低估.现存五大类种子植物——银杏(Ginkgo biloba )、松柏类(conifers)、苏铁类(cycads)、买麻藤目和被子植物中(Wang & Ran, 2014 ), 都已先后报道了不同比例风媒传粉模式的存在(Ackerman, 2000 ; Cully et al, 2002 ). ...
... 物种丰富的热带地区降雨较多、湿度较大、树叶遮挡以及植物分散生长等都不利于风媒传粉, 这些因素在低地雨林中的影响尤为严重.然而, Vamosi等(2006)的大范围调查显示, 这些高生物多样性地区的植物因为虫媒传粉竞争而经历着更严重的花粉限制; 而花粉限制的选择压可能介导植物即使在这样不利的条件下也能进化出风媒传粉机制以寻求生殖保障(Cully et al, 2002 ).Bawa (1990)的早期调查显示, 热带低地雨林中也有1-4%的物种具风媒传粉模式. ...
... 风速是影响风媒传粉的一个重要因素, 不但影响花粉的散播距离, 同时还影响柱头对花粉的捕捉(Whitehead, 1969 ).当风速大到可以克服花粉的U t , 但又不至于阻止柱头对花粉的接收时, 同种个体间的花粉传递效率最高(Niklas, 1985 ; Culley et al, 2002 ).鲁先文等(2008)使用风速计和黏性玻片法检验风速对沙棘(Hippophae rhamnoides )风媒传粉距离的影响, 发现风速较低时花粉传播距离一般不超过25 m; 但风速超过3.5 m/s时, 顺风花粉传播距离可高至85 m.Molina等(2001)使用Burkard定量空气自动取样器研究车前属(Plantago )植物花粉密度与环境因素的关系, 发现温度、湿度和风速对空气中的花粉密度均有显著影响. ...
Insect and wind pollination in Urginea maritima (Liliaceae)
1
1986
... 使用以上方法也可能面临一些问题, 如雨天时玻片捕捉的花粉可能被雨水冲刷至黏性物质下层而难以计数, 又或直接被冲刷掉影响结果准确性.解决的办法是将玻片安放到顶端遮挡、但侧面敞开的简易装置中(如Durham sampler, 详见Ackerman & Kevan, 2005 ).此外, 野外条件下携有花粉的昆虫可能误碰玻片而影响实验结果.因此, 在显微计数时应特别注意一些有明显触碰痕迹、花粉分布极不均匀的玻片, 样本量充足时可考虑将其删除; 还可通过观察确定玻片未被昆虫触碰、对玻片进行套网或在昆虫不活跃期(如夜间)悬挂玻片(Dafni & Dukas, 1986 )等方法, 减少昆虫随机活动对风媒花粉捕获的干扰. ...
The Effects of Cross and Self Fertilisation in the Vegetable Kingdom
1
1876
... 种子植物的花粉传递依赖不同的传粉媒介, 虫媒传粉的多样性、普遍性和高效性使其受到大量关注(黄双全和郭友好, 2000 ), 而水媒和风媒传粉往往被认为效率较低而在自然生态系统中处于劣势地位(Darwin, 1876 ; Faegri & van der Pijl, 1979 ).然而, 新的研究表明, 风媒传粉并不如想象中那么低效, 在某些环境条件或某些植物类群中, 其效率甚至高于虫媒传粉(Friedman & Barrett, 2009 ).目前, 被子植物中已经发现至少65次从动物媒向风媒的传粉转变(Linder, 1998 ; Friedman, 2011 ).传粉模式从虫媒转变为风媒, 相对普遍的原因可能在于植物只需要进化出相对简单的花部特征就可以适应风媒; 但从风媒转变为虫媒在自然界中却很少存在, 可能因为植物要重新获得与吸引、报酬和适应昆虫访花行为相关的复杂花部特征, 会经历更大的进化限制(Huang et al, 2013 ).因此, 通常认为从虫媒到风媒的传粉系统转变在某种程度上是不可逆的(Cox, 1991 ; Dodd et al, 1999 ).按照以上观点, 随着全球自然环境和动物传粉服务水平的双重变化(Hegland et al, 2009 ; Ollerton et al, 2014 ), 植物界中将极可能出现更高比例和多样化的风媒传粉现象. ...
The variability in settling velocities of some pollen and spores
1
1995
... (2)沉降塔法.沉降塔设备主要包括4个部分(详见Di-Giovanni et al, 1995 ; Borrell, 2012 ): 花粉释放装置、双层圆柱筒(提供花粉下落所需的静止空气环境)、转盘(其上装载径向排列的单面涂有凡士林的玻片, 并与花粉的释放同步进行)、控制面板(电动机供电, 可以控制圆盘转速).实验中, 花粉垂直下落距离已知, 根据花粉在转盘玻片上的沉积位置和转盘转速来确定下落时间, 最终求得U t .Borrell (2012)对Di-Giovanni等(1995)的沉降塔设备加以改进, 使用小型的振动刷代替原来的充气装置释放花粉, 对6种禾本科和4种雪松属(Cedrus )植物的U t 进行快速测定; 发现不同物种U t 差异显著, 禾本科植物的U t 平均值在3.81-5.29 cm/s之间, 显著低于雪松属植物的U t (6.72-9.18).作者预测这种差异可以影响花粉的传播距离进而可能影响不同植物的遗传多样性和居群结构. ...
Phylogenetic analysis of trait evolution and species diversity variation among angiosperm families
1
1999
... 种子植物的花粉传递依赖不同的传粉媒介, 虫媒传粉的多样性、普遍性和高效性使其受到大量关注(黄双全和郭友好, 2000 ), 而水媒和风媒传粉往往被认为效率较低而在自然生态系统中处于劣势地位(Darwin, 1876 ; Faegri & van der Pijl, 1979 ).然而, 新的研究表明, 风媒传粉并不如想象中那么低效, 在某些环境条件或某些植物类群中, 其效率甚至高于虫媒传粉(Friedman & Barrett, 2009 ).目前, 被子植物中已经发现至少65次从动物媒向风媒的传粉转变(Linder, 1998 ; Friedman, 2011 ).传粉模式从虫媒转变为风媒, 相对普遍的原因可能在于植物只需要进化出相对简单的花部特征就可以适应风媒; 但从风媒转变为虫媒在自然界中却很少存在, 可能因为植物要重新获得与吸引、报酬和适应昆虫访花行为相关的复杂花部特征, 会经历更大的进化限制(Huang et al, 2013 ).因此, 通常认为从虫媒到风媒的传粉系统转变在某种程度上是不可逆的(Cox, 1991 ; Dodd et al, 1999 ).按照以上观点, 随着全球自然环境和动物传粉服务水平的双重变化(Hegland et al, 2009 ; Ollerton et al, 2014 ), 植物界中将极可能出现更高比例和多样化的风媒传粉现象. ...
Time after time: flowering phenology and biotic interactions
1
2007
... 早期研究表明, 风媒传粉植物的比例往往随纬度和海拔的上升具有明显增加的趋势(Regal, 1982 ).温带地区植物居群相对聚集并且遮挡较少, 因而有利于花粉在空气中传播并成功传递到同种柱头.另外, 温带地区一般都具有严格的季节性, 相对于热带地区其植物生长季节较短, 因此很多树种通过早春开花以增加生长时间, 同时可避免与盛花期植物的传粉竞争.然而, 早春虫媒传粉服务相对缺乏且不稳定, 使得这些植物中很多依赖风媒传粉(Elzinga et al, 2007 ).正因如此, 温带的落叶森林、开阔草原、干旱荒漠及高山地区都易进化出风媒传粉物种(如彭德力等, 2012 ).Regal (1982)以北美木本植物为对象, 统计了不同生境中风媒传粉植物的比例, 发现不同群落间风媒传粉植物比例变化较大, 其中大部分温带森林中都有一半以上的种类是风媒传粉的. ...
3
1979
... 种子植物的花粉传递依赖不同的传粉媒介, 虫媒传粉的多样性、普遍性和高效性使其受到大量关注(黄双全和郭友好, 2000 ), 而水媒和风媒传粉往往被认为效率较低而在自然生态系统中处于劣势地位(Darwin, 1876 ; Faegri & van der Pijl, 1979 ).然而, 新的研究表明, 风媒传粉并不如想象中那么低效, 在某些环境条件或某些植物类群中, 其效率甚至高于虫媒传粉(Friedman & Barrett, 2009 ).目前, 被子植物中已经发现至少65次从动物媒向风媒的传粉转变(Linder, 1998 ; Friedman, 2011 ).传粉模式从虫媒转变为风媒, 相对普遍的原因可能在于植物只需要进化出相对简单的花部特征就可以适应风媒; 但从风媒转变为虫媒在自然界中却很少存在, 可能因为植物要重新获得与吸引、报酬和适应昆虫访花行为相关的复杂花部特征, 会经历更大的进化限制(Huang et al, 2013 ).因此, 通常认为从虫媒到风媒的传粉系统转变在某种程度上是不可逆的(Cox, 1991 ; Dodd et al, 1999 ).按照以上观点, 随着全球自然环境和动物传粉服务水平的双重变化(Hegland et al, 2009 ; Ollerton et al, 2014 ), 植物界中将极可能出现更高比例和多样化的风媒传粉现象. ...
... 而花柱的结构也十分重要, 花柱暴露性、柱头黏性和表面积的增加都有利于提高风媒花粉捕获效率, 如裸子植物能分泌传粉滴、禾本科植物具羽毛状或帚刷状柱头等(Faegri & van der Pijl, 1979 ).朱俊义等(2014)利用扫描电镜技术, 发现部分桦木科植物的花柱表皮细胞能形成乳突, 有利于增加花粉接受面积从而提高风媒传粉效率. ...
... 在自然界中, 仅根据风媒传粉综合征(wind pollination syndrome; 表1 ; Faegri & van der Pijl, 1979 )来判断风媒传粉的有无是不全面的, 有些植物即使不完全具备这些典型特征, 也可能具有风媒传粉模式(Gong et al, 2016 ).在全球气候变化、生境片断化、生物入侵等自然选择压力下, 植物某些细微特征的改变, 都有可能带来传粉模式的转变(Friedman, 2011 ). ...
Generalist passerine pollination of a winter-flowering fruit tree in central China
1
2012
... 其一为网袋因素.一般的网袋都空间有限, 在野外使用过程中很易与植物繁殖器官接触而影响有性生殖过程.为避免这种不利影响, 需对套网处理进行“升级”.如若隔绝的传粉者为鸟类, 则仅需使用支撑较好的细铁丝网即可(如Fang et al, 2012 ); 如果使用尼龙网袋或纱袋隔绝传粉昆虫, 最好使用硬铁丝等制作支架支撑网袋, 并在封口处用绳线或细铁丝密封(见《生物多样性》2007年第15卷第6期封面).此外, 网袋的表面积较大可能富集空气中的花粉, 使得套网组接受比自然组更多的风媒花粉; 相反的情况, 尽管网袋孔径大于花粉直径, 网袋在某种程度上仍有可能阻挡风媒花粉的自由通行.因此, 为验证套袋对花粉的富集或阻碍, 需在套袋内外设置黏性玻片比较花粉密度, 以判断是否受到该类因素的影响(Sacchi & Price, 1988 ). ...
Pollen purification and fractionation by equilibrium density gradient centrifugation
1
1993
... 基于风洞实验测定的不同物种花粉运动的差异, 可通过电脑模拟来评估导致这种空气动力学差异的物理基础(Niklas & Buchmann, 1987 ; Cresswell et al, 2007 ).电脑模拟前需获取的指标有: (1)花粉的密度、大小; (2)雌性生殖器官的大小、形态和普遍的朝向; (3)平均风速(Bolinder et al, 2015 ).花粉密度测定可使用密度梯度离心法(Forster & Flenley, 1993 ), 但如果U t 已知, 也可以采用以下公式: ...
Comparing pollen and spore counts collected with the Rotorod Sampler and Burkard spore trap
1
1999
... 黏性玻片只能对空气中的花粉密度及传播距离进行相对定量测定, 精确测定需借助商业购买的空气颗粒取样器, 如英国Burkard Scientific公司的定量空气自动取样器和美国Sampling Technologies公司的Rotorod取样器.这些仪器都能在单位时间内吸取固定体积的空气从而实现花粉密度的精确测定.前者在测定10 μm以下花粉时效率较高, 而后者更适合测定大于10 μm的花粉(Frenz, 1999 ). ...
Gone with the wind: understanding evolutionary transitions between wind and animal pollination in the angiosperms
2
2011
... 种子植物的花粉传递依赖不同的传粉媒介, 虫媒传粉的多样性、普遍性和高效性使其受到大量关注(黄双全和郭友好, 2000 ), 而水媒和风媒传粉往往被认为效率较低而在自然生态系统中处于劣势地位(Darwin, 1876 ; Faegri & van der Pijl, 1979 ).然而, 新的研究表明, 风媒传粉并不如想象中那么低效, 在某些环境条件或某些植物类群中, 其效率甚至高于虫媒传粉(Friedman & Barrett, 2009 ).目前, 被子植物中已经发现至少65次从动物媒向风媒的传粉转变(Linder, 1998 ; Friedman, 2011 ).传粉模式从虫媒转变为风媒, 相对普遍的原因可能在于植物只需要进化出相对简单的花部特征就可以适应风媒; 但从风媒转变为虫媒在自然界中却很少存在, 可能因为植物要重新获得与吸引、报酬和适应昆虫访花行为相关的复杂花部特征, 会经历更大的进化限制(Huang et al, 2013 ).因此, 通常认为从虫媒到风媒的传粉系统转变在某种程度上是不可逆的(Cox, 1991 ; Dodd et al, 1999 ).按照以上观点, 随着全球自然环境和动物传粉服务水平的双重变化(Hegland et al, 2009 ; Ollerton et al, 2014 ), 植物界中将极可能出现更高比例和多样化的风媒传粉现象. ...
... 在自然界中, 仅根据风媒传粉综合征(wind pollination syndrome; 表1 ; Faegri & van der Pijl, 1979 )来判断风媒传粉的有无是不全面的, 有些植物即使不完全具备这些典型特征, 也可能具有风媒传粉模式(Gong et al, 2016 ).在全球气候变化、生境片断化、生物入侵等自然选择压力下, 植物某些细微特征的改变, 都有可能带来传粉模式的转变(Friedman, 2011 ). ...
Wind of change: new insights on the ecology and evolution of pollination and mating in wind-pollinated plants
3
2009
... 种子植物的花粉传递依赖不同的传粉媒介, 虫媒传粉的多样性、普遍性和高效性使其受到大量关注(黄双全和郭友好, 2000 ), 而水媒和风媒传粉往往被认为效率较低而在自然生态系统中处于劣势地位(Darwin, 1876 ; Faegri & van der Pijl, 1979 ).然而, 新的研究表明, 风媒传粉并不如想象中那么低效, 在某些环境条件或某些植物类群中, 其效率甚至高于虫媒传粉(Friedman & Barrett, 2009 ).目前, 被子植物中已经发现至少65次从动物媒向风媒的传粉转变(Linder, 1998 ; Friedman, 2011 ).传粉模式从虫媒转变为风媒, 相对普遍的原因可能在于植物只需要进化出相对简单的花部特征就可以适应风媒; 但从风媒转变为虫媒在自然界中却很少存在, 可能因为植物要重新获得与吸引、报酬和适应昆虫访花行为相关的复杂花部特征, 会经历更大的进化限制(Huang et al, 2013 ).因此, 通常认为从虫媒到风媒的传粉系统转变在某种程度上是不可逆的(Cox, 1991 ; Dodd et al, 1999 ).按照以上观点, 随着全球自然环境和动物传粉服务水平的双重变化(Hegland et al, 2009 ; Ollerton et al, 2014 ), 植物界中将极可能出现更高比例和多样化的风媒传粉现象. ...
... 对于虫媒传粉起源的被子植物, 风媒传粉被认为是从虫媒传粉转变而来的衍生模式, 业已发现被子植物的许多类群中风媒传粉是独立进化而来的(Friedman & Barrett, 2009 ).Ackerman (2000)的系统调查显示, 风媒传粉的被子植物分布在木兰亚纲、金缕梅亚纲、石竹亚纲、五桠果亚纲、蔷薇亚纲、菊亚纲、泽泻亚纲、棕榈亚纲、鸭跖草亚纲和百合亚纲中.在目的水平, 除鸭跖草亚纲中风媒传粉目的比例高达85%外, 其他各个亚纲都只有不到一半的目中出现了风媒传粉; 而以每目中风媒传粉科的数量做频率分布图, 则发现其符合对数正态分布规律, 即绝大部分的目中仅有0-2个科的植物是风媒传粉的, 比例最高的无患子目, 有高达5个科的植物具有风媒传粉模式.整体来讲, 被子植物中至少有18%的科和10%的种是风媒传粉的(Ackerman, 2000 ). ...
... 除以上花部构成因素外, 开花式样也能对风媒传粉产生重要影响(Friedman & Barrett, 2009 ).一般来讲, 风速随着高度的增加而增加, 对于风媒传粉的雌雄同株物种, 雄花处于较高位置更有利于花粉的散播(Bickel & Freeman, 1993 ).Niklas (1987)利用风洞实验, 发现紧凑和弥散型圆锥花序在花粉接受方面具二态性, 即紧凑型圆锥花序对气流的阻力较大, 所以花粉在花序的上风向处偏离, 而在下风向处通过沉降被捕捉; 而弥散型圆锥花序对气流的阻力较小, 因此大部分的花粉在上风向处与花序碰撞并被捕捉.Friedman和Harder (2004)采用花序捆绑固定法以减少花序的随风移动幅度, 发现该处理降低了紧凑型圆锥花序的花粉移出量, 但不影响柱头花粉接受量; 而在弥散型花序中, 花粉的移出量不受影响, 但柱头花粉接受量却显著降低. ...
Inflorescence architecture and wind pollination in six grass species
2004
传粉昆虫行为的研究方法探讨
2
2007
... 传粉生物学研究中对风媒贡献的忽略往往源自对研究方法理解和运用上的一些误区.如某些植物被观察到了昆虫访问, 抑或其具有某些适应虫媒传粉的特征, 如散发气味、具有鲜艳颜色等, 都被认为是虫媒传粉的; 但最新研究表明, 具以上花部特征的如买麻藤属(Gnetum )和乌头属(Aconitum )植物, 均存在风媒传粉模式(Gong et al, 2016 ; Wang et al, 2016 ).此外, 与虫媒传粉研究相似, 短时的观察和片面的实验并不能得到准确全面的结论(龚燕兵和黄双全, 2007 ); 如静风条件下的风媒传粉实验, 往往会错误评估植物的风媒传粉效率(Niklas, 2015 ).这些研究方法上的缺陷, 极可能导致对植物风媒传粉存在性的不正确描述, 从而严重低估自然界中风媒传粉的普遍性和多样化的风媒传粉策略(Owens et al, 1998 ).基于以上背景, 本文总结并评价了风媒传粉的研究方法, 以期能对开展相关植物繁殖生态和保护研究的学者提供方法学上的参考. ...
... 风媒传递的花粉往往体积较小而肉眼难辨, 相对于可直接观察的动物传粉过程, 其在野外研究中的隐蔽性更强.在野外实验中, 研究者应首选使用“花粉捕获法+传粉者排除法”的组合方法, 明确风媒传粉的可能性和生殖贡献大小.野外时间充足时, 应在各种环境条件下使用该组合方法对比测量风媒传粉效率.如野外条件受限, 研究者应快速收集相关植物组织材料, 在实验室内开展空气动力学研究.然而, 野外实验和室内方法并不能相互替代, 两类方法的综合对比运用才能由表及里地揭示自然传粉现象和发生机理(黄双全, 2012 ).同时应特别注意, 风媒和虫媒传粉可并存于一种植物, 此类研究应结合使用传粉昆虫行为的研究方法(龚燕兵和黄双全, 2007 ).本文就风媒传粉的研究方法进行了相对全面的总结和评价, 希望能对我国开展相关研究的学者有所裨益. ...
传粉昆虫行为的研究方法探讨
2
2007
... 传粉生物学研究中对风媒贡献的忽略往往源自对研究方法理解和运用上的一些误区.如某些植物被观察到了昆虫访问, 抑或其具有某些适应虫媒传粉的特征, 如散发气味、具有鲜艳颜色等, 都被认为是虫媒传粉的; 但最新研究表明, 具以上花部特征的如买麻藤属(Gnetum )和乌头属(Aconitum )植物, 均存在风媒传粉模式(Gong et al, 2016 ; Wang et al, 2016 ).此外, 与虫媒传粉研究相似, 短时的观察和片面的实验并不能得到准确全面的结论(龚燕兵和黄双全, 2007 ); 如静风条件下的风媒传粉实验, 往往会错误评估植物的风媒传粉效率(Niklas, 2015 ).这些研究方法上的缺陷, 极可能导致对植物风媒传粉存在性的不正确描述, 从而严重低估自然界中风媒传粉的普遍性和多样化的风媒传粉策略(Owens et al, 1998 ).基于以上背景, 本文总结并评价了风媒传粉的研究方法, 以期能对开展相关植物繁殖生态和保护研究的学者提供方法学上的参考. ...
... 风媒传递的花粉往往体积较小而肉眼难辨, 相对于可直接观察的动物传粉过程, 其在野外研究中的隐蔽性更强.在野外实验中, 研究者应首选使用“花粉捕获法+传粉者排除法”的组合方法, 明确风媒传粉的可能性和生殖贡献大小.野外时间充足时, 应在各种环境条件下使用该组合方法对比测量风媒传粉效率.如野外条件受限, 研究者应快速收集相关植物组织材料, 在实验室内开展空气动力学研究.然而, 野外实验和室内方法并不能相互替代, 两类方法的综合对比运用才能由表及里地揭示自然传粉现象和发生机理(黄双全, 2012 ).同时应特别注意, 风媒和虫媒传粉可并存于一种植物, 此类研究应结合使用传粉昆虫行为的研究方法(龚燕兵和黄双全, 2007 ).本文就风媒传粉的研究方法进行了相对全面的总结和评价, 希望能对我国开展相关研究的学者有所裨益. ...
Wind or insect pollination? Ambophily in a subtropical gymnosperm Gnetum parvifolium (Gnetales)
5
2016
... 传粉生物学研究中对风媒贡献的忽略往往源自对研究方法理解和运用上的一些误区.如某些植物被观察到了昆虫访问, 抑或其具有某些适应虫媒传粉的特征, 如散发气味、具有鲜艳颜色等, 都被认为是虫媒传粉的; 但最新研究表明, 具以上花部特征的如买麻藤属(Gnetum )和乌头属(Aconitum )植物, 均存在风媒传粉模式(Gong et al, 2016 ; Wang et al, 2016 ).此外, 与虫媒传粉研究相似, 短时的观察和片面的实验并不能得到准确全面的结论(龚燕兵和黄双全, 2007 ); 如静风条件下的风媒传粉实验, 往往会错误评估植物的风媒传粉效率(Niklas, 2015 ).这些研究方法上的缺陷, 极可能导致对植物风媒传粉存在性的不正确描述, 从而严重低估自然界中风媒传粉的普遍性和多样化的风媒传粉策略(Owens et al, 1998 ).基于以上背景, 本文总结并评价了风媒传粉的研究方法, 以期能对开展相关植物繁殖生态和保护研究的学者提供方法学上的参考. ...
... 裸子植物被认为是风媒传粉起源的, 其中银杏和松柏类植物都是风媒传粉的(Owens et al, 1998 ; Jin et al, 2012 ); 苏铁类植物中虫媒和风媒传粉均能发挥作用(王乾等, 1997 ; Kono & Tobe, 2007 ); 买麻藤目中风媒是最主要的传粉方式, 但在各属中比例有所不同(Ickert-Bond & Renner, 2016 ).麻黄属(Ephedra )以风媒传粉为主(Bolinder et al, 2015 ), 而百岁兰(Welwitschia mirabilis )则主要为虫媒传粉(Wetschnig & Depisch, 1999 ), 早期研究认为买麻藤属是专性虫媒传粉的(Kato & Inoue, 1994 ), 但最新研究却揭示了其风媒传粉的存在(Gong et al, 2016 ). ...
... 探寻风媒传粉的可能性, 可首先使用花粉捕获装置(pollen traps)明确植株周围的空气中是否携有花粉; 通过收集目标植物的花粉作为参照, 根据花粉的大小、形状和表面纹饰等特征对捕获的花粉进行计数(如Gong et al, 2016 ).花粉捕获装置需悬挂于植株附近的适当距离, 尽量分多个方向散开, 在每个主方向上可设置距离梯度, 如此可量化风媒花粉的传播能力.根据花期、散粉和柱头可授情况, 可选择在白天或者晚上的合适时间段内悬挂装置.天气干燥有风时更易捕获风媒花粉, 但在某些不利条件如下雨或无风时使用该方法, 则可以对比评价不同环境条件下的风媒传粉效率. ...
... 在自然界中, 仅根据风媒传粉综合征(wind pollination syndrome; 表1 ; Faegri & van der Pijl, 1979 )来判断风媒传粉的有无是不全面的, 有些植物即使不完全具备这些典型特征, 也可能具有风媒传粉模式(Gong et al, 2016 ).在全球气候变化、生境片断化、生物入侵等自然选择压力下, 植物某些细微特征的改变, 都有可能带来传粉模式的转变(Friedman, 2011 ). ...
... 相对于动物传粉研究, 目前我国风媒传粉研究所关注的植物种类仍十分有限.已发表的论文大多仅描述风媒植物的适应特征(如朱俊义等, 2014 ), 或使用野外实验方法研究风媒传粉的影响因素和生殖贡献(如鲁先文等, 2008 ; 崔大练等, 2008 ), 目前仍未见空气动力学方法在我国风媒传粉研究中的应用.此外, 仅有少量研究关注风媒与动物媒的并存现象及传粉转变的适应意义(Huang et al, 2013 ; Gong et al, 2016 ; Wang et al, 2016 ). ...
传粉生物学与植物的进化
1
1994
... U t 就是在静止的气流中, 花粉垂直下落达到稳定状态, 即重力和阻力平衡后的运动速度(Niklas & Spatz, 2012 ), 风媒传粉植物的U t 一般为2-6 cm/s (郭友好, 1994 ).密度低、体积小的花粉往往U t 较小, 因而具有更大的传播距离(Cresswell et al, 2010 ).以下介绍测量U t 的两种常用方法及使用中的问题和改进建议: ...
传粉生物学与植物的进化
1
1994
... U t 就是在静止的气流中, 花粉垂直下落达到稳定状态, 即重力和阻力平衡后的运动速度(Niklas & Spatz, 2012 ), 风媒传粉植物的U t 一般为2-6 cm/s (郭友好, 1994 ).密度低、体积小的花粉往往U t 较小, 因而具有更大的传播距离(Cresswell et al, 2010 ).以下介绍测量U t 的两种常用方法及使用中的问题和改进建议: ...
How does climate warming affect plant-pollinator interactions?
1
2009
... 种子植物的花粉传递依赖不同的传粉媒介, 虫媒传粉的多样性、普遍性和高效性使其受到大量关注(黄双全和郭友好, 2000 ), 而水媒和风媒传粉往往被认为效率较低而在自然生态系统中处于劣势地位(Darwin, 1876 ; Faegri & van der Pijl, 1979 ).然而, 新的研究表明, 风媒传粉并不如想象中那么低效, 在某些环境条件或某些植物类群中, 其效率甚至高于虫媒传粉(Friedman & Barrett, 2009 ).目前, 被子植物中已经发现至少65次从动物媒向风媒的传粉转变(Linder, 1998 ; Friedman, 2011 ).传粉模式从虫媒转变为风媒, 相对普遍的原因可能在于植物只需要进化出相对简单的花部特征就可以适应风媒; 但从风媒转变为虫媒在自然界中却很少存在, 可能因为植物要重新获得与吸引、报酬和适应昆虫访花行为相关的复杂花部特征, 会经历更大的进化限制(Huang et al, 2013 ).因此, 通常认为从虫媒到风媒的传粉系统转变在某种程度上是不可逆的(Cox, 1991 ; Dodd et al, 1999 ).按照以上观点, 随着全球自然环境和动物传粉服务水平的双重变化(Hegland et al, 2009 ; Ollerton et al, 2014 ), 植物界中将极可能出现更高比例和多样化的风媒传粉现象. ...
Terminal settling velocity and physical properties of pollen grains in still air
2016
二十一世纪中国传粉生物学的研究: 良好的开端
1
2012
... 风媒传递的花粉往往体积较小而肉眼难辨, 相对于可直接观察的动物传粉过程, 其在野外研究中的隐蔽性更强.在野外实验中, 研究者应首选使用“花粉捕获法+传粉者排除法”的组合方法, 明确风媒传粉的可能性和生殖贡献大小.野外时间充足时, 应在各种环境条件下使用该组合方法对比测量风媒传粉效率.如野外条件受限, 研究者应快速收集相关植物组织材料, 在实验室内开展空气动力学研究.然而, 野外实验和室内方法并不能相互替代, 两类方法的综合对比运用才能由表及里地揭示自然传粉现象和发生机理(黄双全, 2012 ).同时应特别注意, 风媒和虫媒传粉可并存于一种植物, 此类研究应结合使用传粉昆虫行为的研究方法(龚燕兵和黄双全, 2007 ).本文就风媒传粉的研究方法进行了相对全面的总结和评价, 希望能对我国开展相关研究的学者有所裨益. ...
二十一世纪中国传粉生物学的研究: 良好的开端
1
2012
... 风媒传递的花粉往往体积较小而肉眼难辨, 相对于可直接观察的动物传粉过程, 其在野外研究中的隐蔽性更强.在野外实验中, 研究者应首选使用“花粉捕获法+传粉者排除法”的组合方法, 明确风媒传粉的可能性和生殖贡献大小.野外时间充足时, 应在各种环境条件下使用该组合方法对比测量风媒传粉效率.如野外条件受限, 研究者应快速收集相关植物组织材料, 在实验室内开展空气动力学研究.然而, 野外实验和室内方法并不能相互替代, 两类方法的综合对比运用才能由表及里地揭示自然传粉现象和发生机理(黄双全, 2012 ).同时应特别注意, 风媒和虫媒传粉可并存于一种植物, 此类研究应结合使用传粉昆虫行为的研究方法(龚燕兵和黄双全, 2007 ).本文就风媒传粉的研究方法进行了相对全面的总结和评价, 希望能对我国开展相关研究的学者有所裨益. ...
传粉生物学的研究进展
2
2000
... 种子植物的花粉传递依赖不同的传粉媒介, 虫媒传粉的多样性、普遍性和高效性使其受到大量关注(黄双全和郭友好, 2000 ), 而水媒和风媒传粉往往被认为效率较低而在自然生态系统中处于劣势地位(Darwin, 1876 ; Faegri & van der Pijl, 1979 ).然而, 新的研究表明, 风媒传粉并不如想象中那么低效, 在某些环境条件或某些植物类群中, 其效率甚至高于虫媒传粉(Friedman & Barrett, 2009 ).目前, 被子植物中已经发现至少65次从动物媒向风媒的传粉转变(Linder, 1998 ; Friedman, 2011 ).传粉模式从虫媒转变为风媒, 相对普遍的原因可能在于植物只需要进化出相对简单的花部特征就可以适应风媒; 但从风媒转变为虫媒在自然界中却很少存在, 可能因为植物要重新获得与吸引、报酬和适应昆虫访花行为相关的复杂花部特征, 会经历更大的进化限制(Huang et al, 2013 ).因此, 通常认为从虫媒到风媒的传粉系统转变在某种程度上是不可逆的(Cox, 1991 ; Dodd et al, 1999 ).按照以上观点, 随着全球自然环境和动物传粉服务水平的双重变化(Hegland et al, 2009 ; Ollerton et al, 2014 ), 植物界中将极可能出现更高比例和多样化的风媒传粉现象. ...
... 其三为植物因素.植物复杂的性系统(张大勇, 2004 )、交配系统(陈小勇, 2004 )和传粉系统(黄双全和郭友好, 2000 )可能增加实验结果解读的复杂性.研究者需在套网的基础上增加套袋(同时隔绝风媒和虫媒)和去雄处理并进行组合, 判断自动自交(autogamy)和无融合生殖(apomixis)在风媒传粉植物中发生的可能性(技术路线详见图1 ). ...
传粉生物学的研究进展
2
2000
... 种子植物的花粉传递依赖不同的传粉媒介, 虫媒传粉的多样性、普遍性和高效性使其受到大量关注(黄双全和郭友好, 2000 ), 而水媒和风媒传粉往往被认为效率较低而在自然生态系统中处于劣势地位(Darwin, 1876 ; Faegri & van der Pijl, 1979 ).然而, 新的研究表明, 风媒传粉并不如想象中那么低效, 在某些环境条件或某些植物类群中, 其效率甚至高于虫媒传粉(Friedman & Barrett, 2009 ).目前, 被子植物中已经发现至少65次从动物媒向风媒的传粉转变(Linder, 1998 ; Friedman, 2011 ).传粉模式从虫媒转变为风媒, 相对普遍的原因可能在于植物只需要进化出相对简单的花部特征就可以适应风媒; 但从风媒转变为虫媒在自然界中却很少存在, 可能因为植物要重新获得与吸引、报酬和适应昆虫访花行为相关的复杂花部特征, 会经历更大的进化限制(Huang et al, 2013 ).因此, 通常认为从虫媒到风媒的传粉系统转变在某种程度上是不可逆的(Cox, 1991 ; Dodd et al, 1999 ).按照以上观点, 随着全球自然环境和动物传粉服务水平的双重变化(Hegland et al, 2009 ; Ollerton et al, 2014 ), 植物界中将极可能出现更高比例和多样化的风媒传粉现象. ...
... 其三为植物因素.植物复杂的性系统(张大勇, 2004 )、交配系统(陈小勇, 2004 )和传粉系统(黄双全和郭友好, 2000 )可能增加实验结果解读的复杂性.研究者需在套网的基础上增加套袋(同时隔绝风媒和虫媒)和去雄处理并进行组合, 判断自动自交(autogamy)和无融合生殖(apomixis)在风媒传粉植物中发生的可能性(技术路线详见图1 ). ...
Experimental evidence of insect pollination in Juncaceae, a primarily wind-pollinated family
2
2013
... 种子植物的花粉传递依赖不同的传粉媒介, 虫媒传粉的多样性、普遍性和高效性使其受到大量关注(黄双全和郭友好, 2000 ), 而水媒和风媒传粉往往被认为效率较低而在自然生态系统中处于劣势地位(Darwin, 1876 ; Faegri & van der Pijl, 1979 ).然而, 新的研究表明, 风媒传粉并不如想象中那么低效, 在某些环境条件或某些植物类群中, 其效率甚至高于虫媒传粉(Friedman & Barrett, 2009 ).目前, 被子植物中已经发现至少65次从动物媒向风媒的传粉转变(Linder, 1998 ; Friedman, 2011 ).传粉模式从虫媒转变为风媒, 相对普遍的原因可能在于植物只需要进化出相对简单的花部特征就可以适应风媒; 但从风媒转变为虫媒在自然界中却很少存在, 可能因为植物要重新获得与吸引、报酬和适应昆虫访花行为相关的复杂花部特征, 会经历更大的进化限制(Huang et al, 2013 ).因此, 通常认为从虫媒到风媒的传粉系统转变在某种程度上是不可逆的(Cox, 1991 ; Dodd et al, 1999 ).按照以上观点, 随着全球自然环境和动物传粉服务水平的双重变化(Hegland et al, 2009 ; Ollerton et al, 2014 ), 植物界中将极可能出现更高比例和多样化的风媒传粉现象. ...
... 相对于动物传粉研究, 目前我国风媒传粉研究所关注的植物种类仍十分有限.已发表的论文大多仅描述风媒植物的适应特征(如朱俊义等, 2014 ), 或使用野外实验方法研究风媒传粉的影响因素和生殖贡献(如鲁先文等, 2008 ; 崔大练等, 2008 ), 目前仍未见空气动力学方法在我国风媒传粉研究中的应用.此外, 仅有少量研究关注风媒与动物媒的并存现象及传粉转变的适应意义(Huang et al, 2013 ; Gong et al, 2016 ; Wang et al, 2016 ). ...
The Gnetales: recent insights on their morphology, reproductive biology, chromosome numbers, biogeography, and divergence times
1
2016
... 裸子植物被认为是风媒传粉起源的, 其中银杏和松柏类植物都是风媒传粉的(Owens et al, 1998 ; Jin et al, 2012 ); 苏铁类植物中虫媒和风媒传粉均能发挥作用(王乾等, 1997 ; Kono & Tobe, 2007 ); 买麻藤目中风媒是最主要的传粉方式, 但在各属中比例有所不同(Ickert-Bond & Renner, 2016 ).麻黄属(Ephedra )以风媒传粉为主(Bolinder et al, 2015 ), 而百岁兰(Welwitschia mirabilis )则主要为虫媒传粉(Wetschnig & Depisch, 1999 ), 早期研究认为买麻藤属是专性虫媒传粉的(Kato & Inoue, 1994 ), 但最新研究却揭示了其风媒传粉的存在(Gong et al, 2016 ). ...
The mechanism of pollination drop withdrawal in Ginkgo biloba L
1
2012
... 裸子植物被认为是风媒传粉起源的, 其中银杏和松柏类植物都是风媒传粉的(Owens et al, 1998 ; Jin et al, 2012 ); 苏铁类植物中虫媒和风媒传粉均能发挥作用(王乾等, 1997 ; Kono & Tobe, 2007 ); 买麻藤目中风媒是最主要的传粉方式, 但在各属中比例有所不同(Ickert-Bond & Renner, 2016 ).麻黄属(Ephedra )以风媒传粉为主(Bolinder et al, 2015 ), 而百岁兰(Welwitschia mirabilis )则主要为虫媒传粉(Wetschnig & Depisch, 1999 ), 早期研究认为买麻藤属是专性虫媒传粉的(Kato & Inoue, 1994 ), 但最新研究却揭示了其风媒传粉的存在(Gong et al, 2016 ). ...
Origin of insect pollination
1
1994
... 裸子植物被认为是风媒传粉起源的, 其中银杏和松柏类植物都是风媒传粉的(Owens et al, 1998 ; Jin et al, 2012 ); 苏铁类植物中虫媒和风媒传粉均能发挥作用(王乾等, 1997 ; Kono & Tobe, 2007 ); 买麻藤目中风媒是最主要的传粉方式, 但在各属中比例有所不同(Ickert-Bond & Renner, 2016 ).麻黄属(Ephedra )以风媒传粉为主(Bolinder et al, 2015 ), 而百岁兰(Welwitschia mirabilis )则主要为虫媒传粉(Wetschnig & Depisch, 1999 ), 早期研究认为买麻藤属是专性虫媒传粉的(Kato & Inoue, 1994 ), 但最新研究却揭示了其风媒传粉的存在(Gong et al, 2016 ). ...
Techniques for Pollination Biologists
1
1993
... 传粉者排除法(pollinator exclusion)是验证风媒传粉最直接有效的方法.一般来讲, 研究者需在植物开花前使用合适孔径的网袋, 套网隔绝潜在传粉者但允许风媒花粉进入, 将套网组(风媒)与自然对照组(风媒+虫媒)的坐果率和结籽率进行比较, 便能了解风媒传粉的生殖贡献情况(Kearns & Inouye, 1993 ).Culley等(2002)总结了使用该方法对风媒虫媒混合传粉系统(ambophily)植物的研究, 发现不同植物中, 风媒的传粉贡献比例变化较大(8-93%), 但大部分处于中间水平.在实验过程中, 以下三类因素的影响可能导致实验结果不理想或难以解释, 需在野外引起重视. ...
Is Cycas revoluta (Cycadaceae) wind- or insect-pollinated?
1
2007
... 裸子植物被认为是风媒传粉起源的, 其中银杏和松柏类植物都是风媒传粉的(Owens et al, 1998 ; Jin et al, 2012 ); 苏铁类植物中虫媒和风媒传粉均能发挥作用(王乾等, 1997 ; Kono & Tobe, 2007 ); 买麻藤目中风媒是最主要的传粉方式, 但在各属中比例有所不同(Ickert-Bond & Renner, 2016 ).麻黄属(Ephedra )以风媒传粉为主(Bolinder et al, 2015 ), 而百岁兰(Welwitschia mirabilis )则主要为虫媒传粉(Wetschnig & Depisch, 1999 ), 早期研究认为买麻藤属是专性虫媒传粉的(Kato & Inoue, 1994 ), 但最新研究却揭示了其风媒传粉的存在(Gong et al, 2016 ). ...
Morphology and the evolution of wind pollination
1
1998
... 种子植物的花粉传递依赖不同的传粉媒介, 虫媒传粉的多样性、普遍性和高效性使其受到大量关注(黄双全和郭友好, 2000 ), 而水媒和风媒传粉往往被认为效率较低而在自然生态系统中处于劣势地位(Darwin, 1876 ; Faegri & van der Pijl, 1979 ).然而, 新的研究表明, 风媒传粉并不如想象中那么低效, 在某些环境条件或某些植物类群中, 其效率甚至高于虫媒传粉(Friedman & Barrett, 2009 ).目前, 被子植物中已经发现至少65次从动物媒向风媒的传粉转变(Linder, 1998 ; Friedman, 2011 ).传粉模式从虫媒转变为风媒, 相对普遍的原因可能在于植物只需要进化出相对简单的花部特征就可以适应风媒; 但从风媒转变为虫媒在自然界中却很少存在, 可能因为植物要重新获得与吸引、报酬和适应昆虫访花行为相关的复杂花部特征, 会经历更大的进化限制(Huang et al, 2013 ).因此, 通常认为从虫媒到风媒的传粉系统转变在某种程度上是不可逆的(Cox, 1991 ; Dodd et al, 1999 ).按照以上观点, 随着全球自然环境和动物传粉服务水平的双重变化(Hegland et al, 2009 ; Ollerton et al, 2014 ), 植物界中将极可能出现更高比例和多样化的风媒传粉现象. ...
中国沙棘(Hippophae rhamnoides L. ssp. sinensis Rousi)的开花特性及风媒传粉距离的检测
1
2008
... 相对于动物传粉研究, 目前我国风媒传粉研究所关注的植物种类仍十分有限.已发表的论文大多仅描述风媒植物的适应特征(如朱俊义等, 2014 ), 或使用野外实验方法研究风媒传粉的影响因素和生殖贡献(如鲁先文等, 2008 ; 崔大练等, 2008 ), 目前仍未见空气动力学方法在我国风媒传粉研究中的应用.此外, 仅有少量研究关注风媒与动物媒的并存现象及传粉转变的适应意义(Huang et al, 2013 ; Gong et al, 2016 ; Wang et al, 2016 ). ...
中国沙棘(Hippophae rhamnoides L. ssp. sinensis Rousi)的开花特性及风媒传粉距离的检测
1
2008
... 相对于动物传粉研究, 目前我国风媒传粉研究所关注的植物种类仍十分有限.已发表的论文大多仅描述风媒植物的适应特征(如朱俊义等, 2014 ), 或使用野外实验方法研究风媒传粉的影响因素和生殖贡献(如鲁先文等, 2008 ; 崔大练等, 2008 ), 目前仍未见空气动力学方法在我国风媒传粉研究中的应用.此外, 仅有少量研究关注风媒与动物媒的并存现象及传粉转变的适应意义(Huang et al, 2013 ; Gong et al, 2016 ; Wang et al, 2016 ). ...
Collection and washout of airborne pollens and spores by raindrops
1
1962
... 高降雨量带来的湿度增加可影响花粉的聚集程度, 风媒花粉在潮湿空气中极易变黏而导致U t 增加; 湿度还能影响花药的脱水程度, 从且影响花粉的释放(Niklas, 1985 ).降雨最直接的影响就是“雨滴清除”现象, 即适合大小的雨滴可在较短时间内清除空气中所有的花粉(McDonald, 1962 ).Gong等(2016)在晴雨两种天气条件于小叶买麻藤(Gnetum parvifolium )雌雄株附近悬挂黏性玻片, 发现晴天时, 雄雌株附近玻片均能接收到花粉, 但雨天时雌株附近玻片上却无花粉捕获. ...
Environmental factors affecting airborne pollen concentration in anemophilous species of Plantago
2001
The motion of windborne pollen grains around conifer ovulate cones: implications on wind pollination
1
1984
... 关于风洞实验, 其工作场的建立需满足以下3个条件: (1)配备变速风扇.Niklas和Buchmann (1987)使用变速范围为0.20-15.5 m/s的风扇; (2)工作场中气流的稳定性.风洞中放置充满氦气的中性漂浮球, 通过频闪照明观察球排出风洞的轨迹来判断(Niklas, 1984 ).因闪光频率已知, 该技术还可用来校准气流速度.实验中, 也可借助风速仪对气流速度进行周期性检测; (3)相对均匀的花粉浓度.实验中释放的花粉量可由体积判断, 释放点在风洞内部, 花粉浓度可通过连续监测穿过拍摄区的花粉数量判断.Niklas和Buchmann (1987)对2种麻黄(Ephedra trifurca 和E. nevadensis )的风洞实验结果显示, 某一特定的风速下, 植物在传粉过程中更易接受同种花粉.Cresswell等(2007)使用新鲜的花粉和雌球花材料, 在风速和花粉密度均稳定的风洞中, 对3种松科植物不同部位的花粉接受能力进行了比较, 结果发现3种植物的迎风面均能接受约70%的花粉, 背风面只能接受约2%的花粉, 而左右侧面接受的花粉比例在2种松属(Pinus )植物中无差异, 但在Cedrus libani 中却差异显著. ...
The aerodynamics of wind pollination
4
1985
... 风媒传粉研究往往受诸多实际问题的限制, 如人力成本制约大量黏性玻片的花粉计数, 时间成本制约传粉者排除法的野外开展, 因为从套网处理到结实率数据获取往往需要等候太长时间.在这种情况下, 空气动力学则是一个很好的替代方法.研究者只需在野外快速收集研究物种的组织材料, 通过干燥或冷冻等方法保存后带回实验室, 开展后续实验即可.想在实验室内完全理解花粉在空气中的运动规律和实际效果是十分困难的, 因为太多的变量需要考虑(见Ackerman & Kevan, 2005 ).但就一般研究而言, 风媒传粉效率主要受花粉终端沉降速度(terminal settling velocity, U t )和周围气流加速场两个因素的影响(Niklas, 1985 ).U t 可用来推测植物风媒传粉的可能性, 风洞(wind tunnel)实验可以反映雌蕊周围气流加速场中花粉的真实运动情况, 而电脑模拟可以对其进行计算和评估(Niklas & Buchmann, 1987 ; Bolinder et al, 2015 ). ...
... 其中r 是花粉半径, g是重力加速度, ρ 和ρ 0 分别是花粉和介质的密度, μ是空气的动力粘度系数, U t 是终端沉降速度(Niklas, 1985 ); 而雌性生殖器官的形态和大小可根据实际情况使用数码拍照、MicroCT扫描仪或扫描电镜测定(Cresswell et al, 2007 ).雌性生殖器官周围花粉行为的模拟详见Niklas和Kerchner (1986)设计的程序.Niklas和Buchmann (1987)使用电脑模拟E. trifurca 和E. nevadensis 胚珠周围气流场中花粉的运动行为, 得到了与风洞实验相似的结果; 并且预测, 不同物种花粉大小和密度的差异, 导致各物种花粉具有特定的惯性特征, 结合不同物种胚珠所形成的特异空气动力微环境, 即产生了风媒传粉过程中更易接受同种花粉的现象.Bolinder等(2015)的电脑模拟发现E. foeminea 不具备任何风媒花粉捕捉的功能特征, 这也与其虫媒传粉的早期观点一致. ...
... 风速是影响风媒传粉的一个重要因素, 不但影响花粉的散播距离, 同时还影响柱头对花粉的捕捉(Whitehead, 1969 ).当风速大到可以克服花粉的U t , 但又不至于阻止柱头对花粉的接收时, 同种个体间的花粉传递效率最高(Niklas, 1985 ; Culley et al, 2002 ).鲁先文等(2008)使用风速计和黏性玻片法检验风速对沙棘(Hippophae rhamnoides )风媒传粉距离的影响, 发现风速较低时花粉传播距离一般不超过25 m; 但风速超过3.5 m/s时, 顺风花粉传播距离可高至85 m.Molina等(2001)使用Burkard定量空气自动取样器研究车前属(Plantago )植物花粉密度与环境因素的关系, 发现温度、湿度和风速对空气中的花粉密度均有显著影响. ...
... 高降雨量带来的湿度增加可影响花粉的聚集程度, 风媒花粉在潮湿空气中极易变黏而导致U t 增加; 湿度还能影响花药的脱水程度, 从且影响花粉的释放(Niklas, 1985 ).降雨最直接的影响就是“雨滴清除”现象, 即适合大小的雨滴可在较短时间内清除空气中所有的花粉(McDonald, 1962 ).Gong等(2016)在晴雨两种天气条件于小叶买麻藤(Gnetum parvifolium )雌雄株附近悬挂黏性玻片, 发现晴天时, 雄雌株附近玻片均能接收到花粉, 但雨天时雌株附近玻片上却无花粉捕获. ...
Pollen capture and wind-induced movement of compact and diffuse grass panicles: implications for pollination efficiency
1
1987
... (3)问题和改进建议.为保证精确性, U t 的测定最好采用新鲜或者冻存保鲜的花粉(Karl Niklas, 私人联系).但对于一些野外采集后难以保鲜的花粉, 终端沉降速度只能在野外实时测定.例如买麻藤(Gnetum montanum )的花药一旦离体或稍用力碰触便会氧化发黑, 即使迅速-20℃冻存也无法避免(朱亚如, 个人观察).另外, 沉降塔设备在野外使用不便, Niklas和Buchmann (1987) 的黑箱子法更加便捷.但在实际使用过程中, 我们发现闪光灯的效果并不理想, 而强光手电的效果更佳.可预先在黑箱内壁制作一些刻度条背景, 便于记录花粉下落的距离.使用高清画质单反相机连续摄像, 记录花粉匀速下落的距离和时间, 即可求得U t . ...
A biophysical perspective on the pollination biology of Ephedra nevadensis and E. trifurca
1
2015
... 传粉生物学研究中对风媒贡献的忽略往往源自对研究方法理解和运用上的一些误区.如某些植物被观察到了昆虫访问, 抑或其具有某些适应虫媒传粉的特征, 如散发气味、具有鲜艳颜色等, 都被认为是虫媒传粉的; 但最新研究表明, 具以上花部特征的如买麻藤属(Gnetum )和乌头属(Aconitum )植物, 均存在风媒传粉模式(Gong et al, 2016 ; Wang et al, 2016 ).此外, 与虫媒传粉研究相似, 短时的观察和片面的实验并不能得到准确全面的结论(龚燕兵和黄双全, 2007 ); 如静风条件下的风媒传粉实验, 往往会错误评估植物的风媒传粉效率(Niklas, 2015 ).这些研究方法上的缺陷, 极可能导致对植物风媒传粉存在性的不正确描述, 从而严重低估自然界中风媒传粉的普遍性和多样化的风媒传粉策略(Owens et al, 1998 ).基于以上背景, 本文总结并评价了风媒传粉的研究方法, 以期能对开展相关植物繁殖生态和保护研究的学者提供方法学上的参考. ...
Aerodynamics of pollen capture in two sympatric Ephedra species
3
1987
... 风媒传粉研究往往受诸多实际问题的限制, 如人力成本制约大量黏性玻片的花粉计数, 时间成本制约传粉者排除法的野外开展, 因为从套网处理到结实率数据获取往往需要等候太长时间.在这种情况下, 空气动力学则是一个很好的替代方法.研究者只需在野外快速收集研究物种的组织材料, 通过干燥或冷冻等方法保存后带回实验室, 开展后续实验即可.想在实验室内完全理解花粉在空气中的运动规律和实际效果是十分困难的, 因为太多的变量需要考虑(见Ackerman & Kevan, 2005 ).但就一般研究而言, 风媒传粉效率主要受花粉终端沉降速度(terminal settling velocity, U t )和周围气流加速场两个因素的影响(Niklas, 1985 ).U t 可用来推测植物风媒传粉的可能性, 风洞(wind tunnel)实验可以反映雌蕊周围气流加速场中花粉的真实运动情况, 而电脑模拟可以对其进行计算和评估(Niklas & Buchmann, 1987 ; Bolinder et al, 2015 ). ...
... (1)频闪摄影法.包括黑箱子和玻璃圆筒法, 二者原理相似, 只是提供花粉在静止气流中下落的装置不同: 前者使用内壁涂有亚光材料以减少光散射的黑箱子(Niklas & Buchmann, 1987 ), 后者使用圆柱形的玻璃圆筒(Bolinder et al, 2015 ).测定技术均采用静止流体中花粉下落的频闪摄影技术.实验时需在黑暗的环境中放置好闪光灯和相机镜头, 当闪光灯和相机快门打开的同时释放花粉, 之后捕捉花粉的运动图像.使用匀速下落过程中两张连续图像的间距乘以闪光频率便可求得U t .Bolinder等(2015)使用玻璃圆筒法测得Ephedra foeminea 的U t 在4.7- 21.44 cm/s之间, 这种大的差异是由花粉的不同聚集程度引起的.Hirose和Osada (2016)的玻璃圆筒法也大致类似, 只不过在玻璃圆筒底端安装发射波长为635 nm的激光装置, 替代闪光灯照明以协助花粉运动过程的拍摄; 他们研究的5种植物的U t 平均值分布在1.03-3.2 cm/s之间, 花粉越大的物种, U t 值越大. ...
... 基于风洞实验测定的不同物种花粉运动的差异, 可通过电脑模拟来评估导致这种空气动力学差异的物理基础(Niklas & Buchmann, 1987 ; Cresswell et al, 2007 ).电脑模拟前需获取的指标有: (1)花粉的密度、大小; (2)雌性生殖器官的大小、形态和普遍的朝向; (3)平均风速(Bolinder et al, 2015 ).花粉密度测定可使用密度梯度离心法(Forster & Flenley, 1993 ), 但如果U t 已知, 也可以采用以下公式: ...
Aerodynamics of Ephedra trifurca. II. Computer modelling of pollination efficiencies
1986
1
2012
... U t 就是在静止的气流中, 花粉垂直下落达到稳定状态, 即重力和阻力平衡后的运动速度(Niklas & Spatz, 2012 ), 风媒传粉植物的U t 一般为2-6 cm/s (郭友好, 1994 ).密度低、体积小的花粉往往U t 较小, 因而具有更大的传播距离(Cresswell et al, 2010 ).以下介绍测量U t 的两种常用方法及使用中的问题和改进建议: ...
Extinctions of aculeate pollinators in Britain and the role of large-scale agricultural changes
1
2014
... 种子植物的花粉传递依赖不同的传粉媒介, 虫媒传粉的多样性、普遍性和高效性使其受到大量关注(黄双全和郭友好, 2000 ), 而水媒和风媒传粉往往被认为效率较低而在自然生态系统中处于劣势地位(Darwin, 1876 ; Faegri & van der Pijl, 1979 ).然而, 新的研究表明, 风媒传粉并不如想象中那么低效, 在某些环境条件或某些植物类群中, 其效率甚至高于虫媒传粉(Friedman & Barrett, 2009 ).目前, 被子植物中已经发现至少65次从动物媒向风媒的传粉转变(Linder, 1998 ; Friedman, 2011 ).传粉模式从虫媒转变为风媒, 相对普遍的原因可能在于植物只需要进化出相对简单的花部特征就可以适应风媒; 但从风媒转变为虫媒在自然界中却很少存在, 可能因为植物要重新获得与吸引、报酬和适应昆虫访花行为相关的复杂花部特征, 会经历更大的进化限制(Huang et al, 2013 ).因此, 通常认为从虫媒到风媒的传粉系统转变在某种程度上是不可逆的(Cox, 1991 ; Dodd et al, 1999 ).按照以上观点, 随着全球自然环境和动物传粉服务水平的双重变化(Hegland et al, 2009 ; Ollerton et al, 2014 ), 植物界中将极可能出现更高比例和多样化的风媒传粉现象. ...
Pollination in conifers
2
1998
... 传粉生物学研究中对风媒贡献的忽略往往源自对研究方法理解和运用上的一些误区.如某些植物被观察到了昆虫访问, 抑或其具有某些适应虫媒传粉的特征, 如散发气味、具有鲜艳颜色等, 都被认为是虫媒传粉的; 但最新研究表明, 具以上花部特征的如买麻藤属(Gnetum )和乌头属(Aconitum )植物, 均存在风媒传粉模式(Gong et al, 2016 ; Wang et al, 2016 ).此外, 与虫媒传粉研究相似, 短时的观察和片面的实验并不能得到准确全面的结论(龚燕兵和黄双全, 2007 ); 如静风条件下的风媒传粉实验, 往往会错误评估植物的风媒传粉效率(Niklas, 2015 ).这些研究方法上的缺陷, 极可能导致对植物风媒传粉存在性的不正确描述, 从而严重低估自然界中风媒传粉的普遍性和多样化的风媒传粉策略(Owens et al, 1998 ).基于以上背景, 本文总结并评价了风媒传粉的研究方法, 以期能对开展相关植物繁殖生态和保护研究的学者提供方法学上的参考. ...
... 裸子植物被认为是风媒传粉起源的, 其中银杏和松柏类植物都是风媒传粉的(Owens et al, 1998 ; Jin et al, 2012 ); 苏铁类植物中虫媒和风媒传粉均能发挥作用(王乾等, 1997 ; Kono & Tobe, 2007 ); 买麻藤目中风媒是最主要的传粉方式, 但在各属中比例有所不同(Ickert-Bond & Renner, 2016 ).麻黄属(Ephedra )以风媒传粉为主(Bolinder et al, 2015 ), 而百岁兰(Welwitschia mirabilis )则主要为虫媒传粉(Wetschnig & Depisch, 1999 ), 早期研究认为买麻藤属是专性虫媒传粉的(Kato & Inoue, 1994 ), 但最新研究却揭示了其风媒传粉的存在(Gong et al, 2016 ). ...
高山植物繁殖策略的研究进展
1
2012
... 早期研究表明, 风媒传粉植物的比例往往随纬度和海拔的上升具有明显增加的趋势(Regal, 1982 ).温带地区植物居群相对聚集并且遮挡较少, 因而有利于花粉在空气中传播并成功传递到同种柱头.另外, 温带地区一般都具有严格的季节性, 相对于热带地区其植物生长季节较短, 因此很多树种通过早春开花以增加生长时间, 同时可避免与盛花期植物的传粉竞争.然而, 早春虫媒传粉服务相对缺乏且不稳定, 使得这些植物中很多依赖风媒传粉(Elzinga et al, 2007 ).正因如此, 温带的落叶森林、开阔草原、干旱荒漠及高山地区都易进化出风媒传粉物种(如彭德力等, 2012 ).Regal (1982)以北美木本植物为对象, 统计了不同生境中风媒传粉植物的比例, 发现不同群落间风媒传粉植物比例变化较大, 其中大部分温带森林中都有一半以上的种类是风媒传粉的. ...
高山植物繁殖策略的研究进展
1
2012
... 早期研究表明, 风媒传粉植物的比例往往随纬度和海拔的上升具有明显增加的趋势(Regal, 1982 ).温带地区植物居群相对聚集并且遮挡较少, 因而有利于花粉在空气中传播并成功传递到同种柱头.另外, 温带地区一般都具有严格的季节性, 相对于热带地区其植物生长季节较短, 因此很多树种通过早春开花以增加生长时间, 同时可避免与盛花期植物的传粉竞争.然而, 早春虫媒传粉服务相对缺乏且不稳定, 使得这些植物中很多依赖风媒传粉(Elzinga et al, 2007 ).正因如此, 温带的落叶森林、开阔草原、干旱荒漠及高山地区都易进化出风媒传粉物种(如彭德力等, 2012 ).Regal (1982)以北美木本植物为对象, 统计了不同生境中风媒传粉植物的比例, 发现不同群落间风媒传粉植物比例变化较大, 其中大部分温带森林中都有一半以上的种类是风媒传粉的. ...
Pollination by wind and animals: ecology of geographic patterns
2
1982
... 风媒传粉植物在地球上并不是随机分布的, 而是具有明显的地理和生态趋势(Whitehead, 1969 ; Regal, 1982 ).风媒花粉的成功传递不仅由风力决定, 许多生物与非生物因素都对风媒传粉有着重要影响, 如不同生境中植物群落结构的多样性、动物传粉服务的水平和稳定性、温湿度和降雨量等, 都能显著影响该生境中风媒传粉植物的比例(详见表1 和下文第3节). ...
... 早期研究表明, 风媒传粉植物的比例往往随纬度和海拔的上升具有明显增加的趋势(Regal, 1982 ).温带地区植物居群相对聚集并且遮挡较少, 因而有利于花粉在空气中传播并成功传递到同种柱头.另外, 温带地区一般都具有严格的季节性, 相对于热带地区其植物生长季节较短, 因此很多树种通过早春开花以增加生长时间, 同时可避免与盛花期植物的传粉竞争.然而, 早春虫媒传粉服务相对缺乏且不稳定, 使得这些植物中很多依赖风媒传粉(Elzinga et al, 2007 ).正因如此, 温带的落叶森林、开阔草原、干旱荒漠及高山地区都易进化出风媒传粉物种(如彭德力等, 2012 ).Regal (1982)以北美木本植物为对象, 统计了不同生境中风媒传粉植物的比例, 发现不同群落间风媒传粉植物比例变化较大, 其中大部分温带森林中都有一半以上的种类是风媒传粉的. ...
Pollination of the arroyo willow, Salix lasiolepis: role of insects and wind
1
1988
... 其一为网袋因素.一般的网袋都空间有限, 在野外使用过程中很易与植物繁殖器官接触而影响有性生殖过程.为避免这种不利影响, 需对套网处理进行“升级”.如若隔绝的传粉者为鸟类, 则仅需使用支撑较好的细铁丝网即可(如Fang et al, 2012 ); 如果使用尼龙网袋或纱袋隔绝传粉昆虫, 最好使用硬铁丝等制作支架支撑网袋, 并在封口处用绳线或细铁丝密封(见《生物多样性》2007年第15卷第6期封面).此外, 网袋的表面积较大可能富集空气中的花粉, 使得套网组接受比自然组更多的风媒花粉; 相反的情况, 尽管网袋孔径大于花粉直径, 网袋在某种程度上仍有可能阻挡风媒花粉的自由通行.因此, 为验证套袋对花粉的富集或阻碍, 需在套袋内外设置黏性玻片比较花粉密度, 以判断是否受到该类因素的影响(Sacchi & Price, 1988 ). ...
Pollen morphology and ultrastructure of several Gnetum species: an electron microscopic study
1
2016
... 花粉的大小、密度、形态和超微结构都能影响风媒传粉效率, 风媒花粉的大小一般在20-40 μm, 表面平滑干燥以减少花粉聚集(Ackerman & Kevan, 2005 ), 一些风媒物种的花粉壁内部存在大量空白区域, 有利于增加其浮力(Wodehouse, 1935 ).利用电镜技术就能清楚观察到花粉表面和内部结构差异(Tekleva, 2016 ).Bolinder等(2015)利用扫描和透射电镜技术对麻黄属植物的花粉进行研究, 发现花粉壁的超微结构与终端沉降速度是相关联的, 花粉壁顶盖下层空间内颗粒物质的相对比例和数量共同决定花粉密度, 并与整体的大小一起决定U t , 进而影响花粉散播能力. ...
Pollination decays in biodiversity hotspots
2006
Intensified wind pollination mediated by pollen dimorphism after range expansion in an ambophilous biennial Aconitum gymnandrum
2
2016
... 传粉生物学研究中对风媒贡献的忽略往往源自对研究方法理解和运用上的一些误区.如某些植物被观察到了昆虫访问, 抑或其具有某些适应虫媒传粉的特征, 如散发气味、具有鲜艳颜色等, 都被认为是虫媒传粉的; 但最新研究表明, 具以上花部特征的如买麻藤属(Gnetum )和乌头属(Aconitum )植物, 均存在风媒传粉模式(Gong et al, 2016 ; Wang et al, 2016 ).此外, 与虫媒传粉研究相似, 短时的观察和片面的实验并不能得到准确全面的结论(龚燕兵和黄双全, 2007 ); 如静风条件下的风媒传粉实验, 往往会错误评估植物的风媒传粉效率(Niklas, 2015 ).这些研究方法上的缺陷, 极可能导致对植物风媒传粉存在性的不正确描述, 从而严重低估自然界中风媒传粉的普遍性和多样化的风媒传粉策略(Owens et al, 1998 ).基于以上背景, 本文总结并评价了风媒传粉的研究方法, 以期能对开展相关植物繁殖生态和保护研究的学者提供方法学上的参考. ...
... 相对于动物传粉研究, 目前我国风媒传粉研究所关注的植物种类仍十分有限.已发表的论文大多仅描述风媒植物的适应特征(如朱俊义等, 2014 ), 或使用野外实验方法研究风媒传粉的影响因素和生殖贡献(如鲁先文等, 2008 ; 崔大练等, 2008 ), 目前仍未见空气动力学方法在我国风媒传粉研究中的应用.此外, 仅有少量研究关注风媒与动物媒的并存现象及传粉转变的适应意义(Huang et al, 2013 ; Gong et al, 2016 ; Wang et al, 2016 ). ...
攀枝花苏铁传粉生物学研究
1
1997
... 裸子植物被认为是风媒传粉起源的, 其中银杏和松柏类植物都是风媒传粉的(Owens et al, 1998 ; Jin et al, 2012 ); 苏铁类植物中虫媒和风媒传粉均能发挥作用(王乾等, 1997 ; Kono & Tobe, 2007 ); 买麻藤目中风媒是最主要的传粉方式, 但在各属中比例有所不同(Ickert-Bond & Renner, 2016 ).麻黄属(Ephedra )以风媒传粉为主(Bolinder et al, 2015 ), 而百岁兰(Welwitschia mirabilis )则主要为虫媒传粉(Wetschnig & Depisch, 1999 ), 早期研究认为买麻藤属是专性虫媒传粉的(Kato & Inoue, 1994 ), 但最新研究却揭示了其风媒传粉的存在(Gong et al, 2016 ). ...
攀枝花苏铁传粉生物学研究
1
1997
... 裸子植物被认为是风媒传粉起源的, 其中银杏和松柏类植物都是风媒传粉的(Owens et al, 1998 ; Jin et al, 2012 ); 苏铁类植物中虫媒和风媒传粉均能发挥作用(王乾等, 1997 ; Kono & Tobe, 2007 ); 买麻藤目中风媒是最主要的传粉方式, 但在各属中比例有所不同(Ickert-Bond & Renner, 2016 ).麻黄属(Ephedra )以风媒传粉为主(Bolinder et al, 2015 ), 而百岁兰(Welwitschia mirabilis )则主要为虫媒传粉(Wetschnig & Depisch, 1999 ), 早期研究认为买麻藤属是专性虫媒传粉的(Kato & Inoue, 1994 ), 但最新研究却揭示了其风媒传粉的存在(Gong et al, 2016 ). ...
Evolution and biogeography of gymnosperms
1
2014
... 种子植物在不同选择压力下进化出多样化的传粉模式, 其中虫媒传粉植物的普遍性已受到大量关注, 而风媒传粉植物的种类多样性往往被低估.现存五大类种子植物——银杏(Ginkgo biloba )、松柏类(conifers)、苏铁类(cycads)、买麻藤目和被子植物中(Wang & Ran, 2014 ), 都已先后报道了不同比例风媒传粉模式的存在(Ackerman, 2000 ; Cully et al, 2002 ). ...
Pollination biology of Welwitschia mirabilis HOOK. f. (Welwitschiaceae, Gnetopsida)
1
1999
... 裸子植物被认为是风媒传粉起源的, 其中银杏和松柏类植物都是风媒传粉的(Owens et al, 1998 ; Jin et al, 2012 ); 苏铁类植物中虫媒和风媒传粉均能发挥作用(王乾等, 1997 ; Kono & Tobe, 2007 ); 买麻藤目中风媒是最主要的传粉方式, 但在各属中比例有所不同(Ickert-Bond & Renner, 2016 ).麻黄属(Ephedra )以风媒传粉为主(Bolinder et al, 2015 ), 而百岁兰(Welwitschia mirabilis )则主要为虫媒传粉(Wetschnig & Depisch, 1999 ), 早期研究认为买麻藤属是专性虫媒传粉的(Kato & Inoue, 1994 ), 但最新研究却揭示了其风媒传粉的存在(Gong et al, 2016 ). ...
Wind pollination in the angiosperms: evolutionary and environmental considerations
2
1969
... 风媒传粉植物在地球上并不是随机分布的, 而是具有明显的地理和生态趋势(Whitehead, 1969 ; Regal, 1982 ).风媒花粉的成功传递不仅由风力决定, 许多生物与非生物因素都对风媒传粉有着重要影响, 如不同生境中植物群落结构的多样性、动物传粉服务的水平和稳定性、温湿度和降雨量等, 都能显著影响该生境中风媒传粉植物的比例(详见表1 和下文第3节). ...
... 风速是影响风媒传粉的一个重要因素, 不但影响花粉的散播距离, 同时还影响柱头对花粉的捕捉(Whitehead, 1969 ).当风速大到可以克服花粉的U t , 但又不至于阻止柱头对花粉的接收时, 同种个体间的花粉传递效率最高(Niklas, 1985 ; Culley et al, 2002 ).鲁先文等(2008)使用风速计和黏性玻片法检验风速对沙棘(Hippophae rhamnoides )风媒传粉距离的影响, 发现风速较低时花粉传播距离一般不超过25 m; 但风速超过3.5 m/s时, 顺风花粉传播距离可高至85 m.Molina等(2001)使用Burkard定量空气自动取样器研究车前属(Plantago )植物花粉密度与环境因素的关系, 发现温度、湿度和风速对空气中的花粉密度均有显著影响. ...
Pollen Grains: Their Structure, Identification and Significance in Science and Medicine
1
1935
... 花粉的大小、密度、形态和超微结构都能影响风媒传粉效率, 风媒花粉的大小一般在20-40 μm, 表面平滑干燥以减少花粉聚集(Ackerman & Kevan, 2005 ), 一些风媒物种的花粉壁内部存在大量空白区域, 有利于增加其浮力(Wodehouse, 1935 ).利用电镜技术就能清楚观察到花粉表面和内部结构差异(Tekleva, 2016 ).Bolinder等(2015)利用扫描和透射电镜技术对麻黄属植物的花粉进行研究, 发现花粉壁的超微结构与终端沉降速度是相关联的, 花粉壁顶盖下层空间内颗粒物质的相对比例和数量共同决定花粉密度, 并与整体的大小一起决定U t , 进而影响花粉散播能力. ...
1
2004
... 其三为植物因素.植物复杂的性系统(张大勇, 2004 )、交配系统(陈小勇, 2004 )和传粉系统(黄双全和郭友好, 2000 )可能增加实验结果解读的复杂性.研究者需在套网的基础上增加套袋(同时隔绝风媒和虫媒)和去雄处理并进行组合, 判断自动自交(autogamy)和无融合生殖(apomixis)在风媒传粉植物中发生的可能性(技术路线详见图1 ). ...
1
2004
... 其三为植物因素.植物复杂的性系统(张大勇, 2004 )、交配系统(陈小勇, 2004 )和传粉系统(黄双全和郭友好, 2000 )可能增加实验结果解读的复杂性.研究者需在套网的基础上增加套袋(同时隔绝风媒和虫媒)和去雄处理并进行组合, 判断自动自交(autogamy)和无融合生殖(apomixis)在风媒传粉植物中发生的可能性(技术路线详见图1 ). ...
桦木科植物花柱适应风媒传粉的特征
1
2014
... 相对于动物传粉研究, 目前我国风媒传粉研究所关注的植物种类仍十分有限.已发表的论文大多仅描述风媒植物的适应特征(如朱俊义等, 2014 ), 或使用野外实验方法研究风媒传粉的影响因素和生殖贡献(如鲁先文等, 2008 ; 崔大练等, 2008 ), 目前仍未见空气动力学方法在我国风媒传粉研究中的应用.此外, 仅有少量研究关注风媒与动物媒的并存现象及传粉转变的适应意义(Huang et al, 2013 ; Gong et al, 2016 ; Wang et al, 2016 ). ...
桦木科植物花柱适应风媒传粉的特征
1
2014
... 相对于动物传粉研究, 目前我国风媒传粉研究所关注的植物种类仍十分有限.已发表的论文大多仅描述风媒植物的适应特征(如朱俊义等, 2014 ), 或使用野外实验方法研究风媒传粉的影响因素和生殖贡献(如鲁先文等, 2008 ; 崔大练等, 2008 ), 目前仍未见空气动力学方法在我国风媒传粉研究中的应用.此外, 仅有少量研究关注风媒与动物媒的并存现象及传粉转变的适应意义(Huang et al, 2013 ; Gong et al, 2016 ; Wang et al, 2016 ). ...

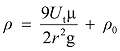


{kind=link}
{kind=link}