
|
|
||
|
垂枝柏复合体的物种界定
生物多样性
2026, 34 (2):
25373-.
DOI: 10.17520/biods.2025373
物种是生物学研究和生物多样性保护与管理的基本单元。近年来, 整合物种概念逐渐广受青睐, 其强调近缘物种之间划分时应该满足多方面的证据。小果垂枝柏(Juniperus coxii)原为垂枝柏(J. recurva)的变种, 后虽被提升为独立物种, 但目前尚缺乏群体水平的证据支持。本文覆盖横断山-喜马拉雅东部区域, 采集垂枝柏复合体26个居群197个个体样品, 一方面基于叶绿体DNA数据划分谱系, 另一方面开展代表性居群形态学统计分析和生态位分化分析, 并整合本文的样本采集地点和已发布采集地点的环境气候因子开展生态位模拟, 旨在更准确地厘清两个分类群的界限。研究结果显示: 叶绿体单倍型网络分析将垂枝柏复合体分成东部和西部两个谱系, 地理分布上大致以错那市为界; 两个谱系在形态上存在一定程度的差异, 但重叠度较高; 东、西谱系生态位存在显著分化。以上证据支持将错那市以西的居群划分为垂枝柏, 而将错那市以东的居群划分为小果垂枝柏。本文初步厘清了垂枝柏和小果垂枝柏之间的物种界限, 为横断山-喜马拉雅区域的物种界定研究提供新案例, 对于该地区垂枝柏复合体的生物多样性保护与管理具有重要意义。 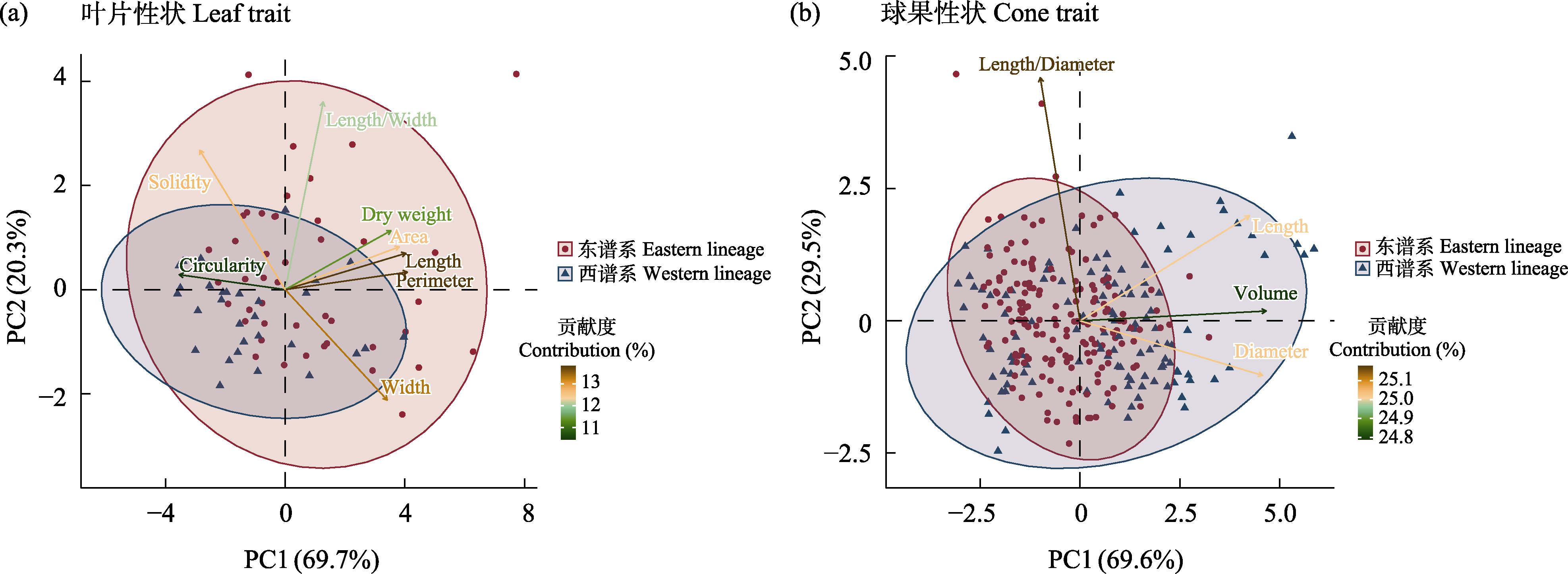 View image in article
图2
垂枝柏复合体形态性状分化。(a)叶片性状。Leaf area: 叶片面积; Length: 叶片长度; Width: 叶片宽度; Perimeter: 叶片周长; Circularity: 叶片圆度; Solidity: 叶片紧凑度; Dry weight: 叶片干重; Length/Width: 叶片长宽比。(b)球果性状。Length: 球果长度; Diameter: 球果直径; Volume: 球果体积。Length/diameter: 球果的长径比。
正文中引用本图/表的段落
东、西部两个谱系的8个叶片性状PCA中, PC1占总变异的69.7%, PC2占总变异的20.3%, PC1和PC2能够解释90.0%的变异; 4个球果性状PCA中, PC1占总变异的69.6%, PC2占总变异的29.5%, PC1和PC2能够解释99.1%的变异。无论是叶片性状, 还是球果性状, 两个谱系均存在一定程度的差异, 但因重叠较多, 未检测到统计间断(图2)。相比于西谱系, 东谱系的叶片具有更广泛的变异范围, 而球果的变异范围相对较小。
物种分化是一个持续且反复的过程, 自然界中多数物种仍处于分化路上(刘建全, 2016)。自然界中存在完全分化的近缘物种相对较少, 针对处于分化早中期的类群, 物种界定更为复杂, 需要整合系统发育、生态学和形态学等物种概念开展物种界定工作。以往仅依靠形态特征辨别近缘物种, 但种间无明显间断的形态特征给分类学家造成了困扰, 可能区分不出近缘物种或将同一物种拆分为多个(Duminil & Di Michele, 2009)。Ruiz-Sanchez & Sosa (2010)使用分子、形态和生态位数据对分布于新热带地区的墨西哥竹属(Otatea)物种重新划定, 将原先的3种扩充为7种。Li (2019)等通过群体水平的形态、分子数据和生态位模拟, 将青藏高原红景天属(Rhodiola)原来根据形态定义的7种重新划分为3种。本研究整合叶绿体SNP单倍型网络和系统发育关系、叶绿体基因组的系统发育关系、形态和生态位数据对垂枝柏复合体进行物种界定。单倍型网络分析揭示该复合体存在东部与西部两个谱系(图1a, 附录5)。基于SNP和叶绿体基因组的系统发育树均支持小果垂枝柏和垂枝柏各自构成单系群(图1c, 附录6)。野外调查发现, 二者无法通过形态直接区分。形态性状分析表明, 尽管叶片和球果性状存在一定差异, 但不存在统计间断(洪德元, 2016a; 图2)。因此, 仅基于形态性状无法区分小果垂枝柏和垂枝柏, 需对微观形态性状开展深入观察和研究。
本文的其它图/表
|
备案号:京ICP备16067583号-7
Copyright © 2026 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn
![]()

{kind=link}