|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
基于叶绿体基因组的江南牡丹草遗传多样性与遗传结构
生物多样性
2025, 33 (8):
25149-.
DOI: 10.17520/biods.2025149
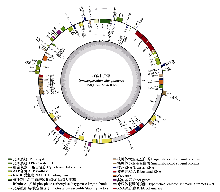
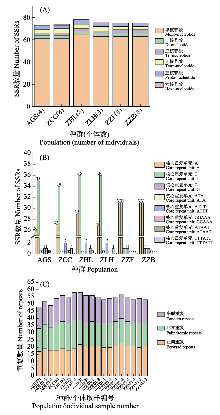
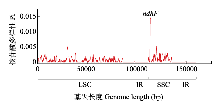
江南牡丹草(Gymnospermium kiangnanense)是分布于安徽和浙江的中国特有濒危植物, 已被纳入当地的珍稀濒危植物名录, 然而对其种群间遗传结构的相关研究较少。为了加强对江南牡丹草的保护, 本研究通过组装6个种群39个个体的叶绿体基因组序列并对其进行比较基因组学分析和种群遗传结构分析, 探讨了对江南牡丹草的保护策略。结果表明: (1)江南牡丹草叶绿体基因组在序列组成、基因结构和基因含量等方面都高度保守, 其中简单重复序列(simple sequence repeats, SSRs)类型表现出明显的种群特征; (2)叶绿体基因组中3个非编码区域: psbZ与trnG-GCC的间隔区(psbZ-trnG-GCC)、trnT-UGU与trnL-UAA的间隔区(trnT-UGU-trnL-UAA)以及ycf1与ndhF的间隔区(ycf1-ndhF), 均表现出较高的变异性, 同时ndhF具有较高的核苷酸多样性, 可作为潜在的分子标记; (3)叶绿体基因组具有较高的遗传多样性, 且种群间具有较高的遗传分化; (4) 6个种群39个个体的叶绿体基因组序列计算得出14个单倍型, 经过Network分析和Beast分析可以划分为3个支系; (5)江南牡丹草种群间的变异大, 且具有明显的谱系地理结构; (6)通过种群历史动态分析发现, 江南牡丹草种群未发生扩张, 一直处于平衡状态。江南牡丹草以异交的繁殖方式增加遗传变异、减少近交衰退, 再加上在中国东部存在的第四纪冰期避难所为其提供了稳定的生存环境, 因而其种群的遗传多样性较高。结实率低、种子扩散能力弱以及过度的人为活动可能是导致其濒危的主要原因, 因此通过对江南牡丹草叶绿体基因组的分析结合保护遗传学提出如下保护建议: (1)按支系划分3个保护单元, 重点保护浙江诸暨凤林下村种群, 设立保护小区; (2)在其花期进行人工放蜂, 增加传粉率和结实率; (3)降低群落的种群密度以改善光照, 促进幼苗生长; (4)在就地保护基础上, 于适宜植物园开展迁地保护与人工繁育; (5)加强科普宣传, 减少人为破坏; (6)合理利用其药用价值, 促进人工繁育与保护。
表1
江南牡丹草样品采集信息和遗传多样性
正文中引用本图/表的段落
江南牡丹草(Gymnospermium kiangnanense)是分布于安徽和浙江的中国特有濒危植物, 已被纳入当地的珍稀濒危植物名录, 然而对其种群间遗传结构的相关研究较少。为了加强对江南牡丹草的保护, 本研究通过组装6个种群39个个体的叶绿体基因组序列并对其进行比较基因组学分析和种群遗传结构分析, 探讨了对江南牡丹草的保护策略。结果表明: (1)江南牡丹草叶绿体基因组在序列组成、基因结构和基因含量等方面都高度保守, 其中简单重复序列(simple sequence repeats, SSRs)类型表现出明显的种群特征; (2)叶绿体基因组中3个非编码区域: psbZ与trnG-GCC的间隔区(psbZ-trnG-GCC)、trnT-UGU与trnL-UAA的间隔区(trnT-UGU-trnL-UAA)以及ycf1与ndhF的间隔区(ycf1-ndhF), 均表现出较高的变异性, 同时ndhF具有较高的核苷酸多样性, 可作为潜在的分子标记; (3)叶绿体基因组具有较高的遗传多样性, 且种群间具有较高的遗传分化; (4) 6个种群39个个体的叶绿体基因组序列计算得出14个单倍型, 经过Network分析和Beast分析可以划分为3个支系; (5)江南牡丹草种群间的变异大, 且具有明显的谱系地理结构; (6)通过种群历史动态分析发现, 江南牡丹草种群未发生扩张, 一直处于平衡状态。江南牡丹草以异交的繁殖方式增加遗传变异、减少近交衰退, 再加上在中国东部存在的第四纪冰期避难所为其提供了稳定的生存环境, 因而其种群的遗传多样性较高。结实率低、种子扩散能力弱以及过度的人为活动可能是导致其濒危的主要原因, 因此通过对江南牡丹草叶绿体基因组的分析结合保护遗传学提出如下保护建议: (1)按支系划分3个保护单元, 重点保护浙江诸暨凤林下村种群, 设立保护小区; (2)在其花期进行人工放蜂, 增加传粉率和结实率; (3)降低群落的种群密度以改善光照, 促进幼苗生长; (4)在就地保护基础上, 于适宜植物园开展迁地保护与人工繁育; (5)加强科普宣传, 减少人为破坏; (6)合理利用其药用价值, 促进人工繁育与保护。
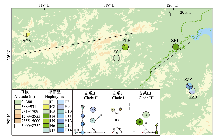
通过野外工作共采集浙江、安徽两个省份6个采样点的39份江南牡丹草实验材料, 基本覆盖该植物的所有分布区。为保证样本代表性, 相邻采集个体间隔不少于50 m, 每个个体采集3-5片新鲜健康的叶片。样品按区域分为AGS (安徽池州贵池石门村)、ZCC (浙江杭州淳安菖蒲村)、ZHL (浙江杭州西湖仙桥洞)、ZLH (浙江临安河桥陆平下村)、ZZB (浙江诸暨半丘村)和ZZF (浙江诸暨凤林下村) 6个组(表1)。
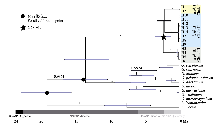
所分析的39个个体中, 对位排列后序列总长度为156,781 bp, 包含324个变异位点, 共检测到14个单倍型, 总单倍型多样性为0.9028, 浙江诸暨凤林下村种群的单倍型多样性和总核苷酸多态性都最高。所有的共享单倍型仅发生在同一地理种群不同个体之间, 不同种群之间不存在共享单倍型(表1)。通过Permut计算所得种群内平均遗传多样性为0.456, 总的遗传多样性值为1.000。
本文的其它图/表
|