|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
雌雄异株物种一把伞南星雌雄株空间分布及性别二态性
生物多样性
2024, 32 (6):
24084-.
DOI: 10.17520/biods.2024084
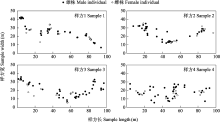
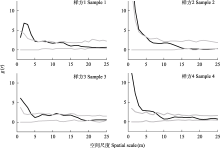
雌雄异株是有花植物雌雄性别功能在个体水平上由结合到分离的重要进化转变之一, 发生于被子植物的多个科属。雌雄异株物种的雌雄个体通常在分布、形态等方面存在差异。研究其雌雄个体的空间分布及性别二态性, 是理解雌雄异株物种繁殖策略及其适应与进化的重要前提。天南星属(Arisaema)是天南星科内已报道唯一存在雌雄异株且较为进化的类群, 一把伞南星(A. erubescens)是该属中不同性别个体存在差异的多年生草本植物。本文以湖南省八大公山国家级自然保护区内分布的一把伞南星为研究对象, 通过研究其雌雄个体的空间分布、花期物候、形态结构及访花者类群, 以期明确该物种雌雄植株的空间关联性以及花期物候、形态结构的性别二态性。结果表明: 一把伞南星群体内性比均呈极显著雄性偏倚, 雌雄个体在小尺度范围呈聚集分布, 但在较大尺度上呈随机分布; 雄性个体比雌性个体更早进入花期, 置换多元方差分析显示雌雄植株间形态特征存在显著差异, 其中假茎基径、佛焰苞管部直径、花序基径、花序柄直径等特征对性别二态性有重要贡献; 主要传粉者为白天活动的菌蚊科菌蚊属(Mycetophila)的蕈蚊, 其对雄株的访问频率极显著高于雌株。这些结果表明: 雌雄异株物种一把伞南星不同性别植株的空间关联与其传粉者蕈蚊的活动习性相匹配, 其开花物候、形态特征及传粉者访问等方面的性别二态性是对繁殖成功的保障。
表2
一把伞南星雌雄特征的比较(广义线性模型) (平均值 ± 标准误)。粗体表示具显著性差异(P < 0.05)特征中的较大值。
正文中引用本图/表的段落
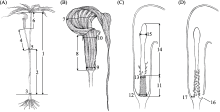
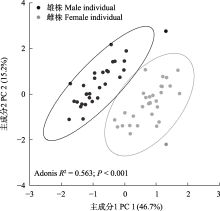
在本研究所测一把伞南星的20个形态性状中, 除佛焰苞檐宽、附属器直径和花序柄长外, 其余性状均表现出显著的性别二态性(表2)。尤其是雄株佛焰苞席卷边缘重叠处形成一个截面积约4.26 ± 0.36 mm2的水滴形小孔(exit hole), 且肉穗花序基部存在一个直径约2 mm的环形通道连接该孔, 而雌株佛焰苞花序基部无此结构(图1C)。除29%的雌株具2片叶外, 其余雌株为1片叶, 而雄株均为1片叶; 表现出二态性的特征中, 除花序长和小花直径外, 雌株的特征值均显著大于雄株(P < 0.05) (表2), 表明雌株较雄株高, 且具相对较大的茎干直径和佛焰苞花序。
雌雄异株常与攀援型木本植物相关, 且多为非生物传粉(Renner & Ricklefs, 1995)。对于生物传粉的雌雄异株物种, 雌雄繁殖特征的差异程度往往会受到一定限制: 如果传粉者偏好访问一种性别的植株, 或者两种性别的植株吸引不同的传粉者, 那么两性性状的差异将可能影响传粉成功(Delph & Ashman, 2006; Barrett & Hough, 2013)。因此它们的花通常不如两性花艳丽, 花朵较小, 常为白色、淡黄色或绿色(Vamosi et al, 2003)。对原产日本的6种天南星调查发现, 性别二态性主要体现在开花时间和繁殖结构上(Kinoshita, 1986), 本研究结果与之一致: 首先, 连续两年的观测发现一把伞南星雄株平均始花时间均极显著早于雌株, 雄株开花较早具有一定适应意义: 能够保证雌株开花时有花粉供其受精结实, 同时与其他雄性个体相比, 花期较早的雄性个体可能有更多机会使同种胚珠受精, 从而获得更高的繁殖适合度(Bullock & Bawa, 1981)。其次, 一把伞南星雌雄花序均为浅绿色佛焰苞包裹着肉穗花序, 但两种性别植株形态大小存在显著差异(表2, 图5), 主要体现在假茎基径、佛焰苞管部直径、花序基径、花序柄直径等方面(附录5), 总体来说雌株较雄株高, 且具相对较大的茎干直径和佛焰苞花序。由于雄性的繁殖阶段不需要结实及产生种子, 因此雌性将产生更高的资源消耗, 故需要更大的体型进行资源生产和积累(胡光万, 2008)。最后, 值得注意的是虽然一把伞南星雌雄个体的主要访花者均为菌蚊属蕈蚊, 但其对雄性个体的访问频率约是对雌性个体访问频率的2倍(图6A)。天南星属物种几乎不产花蜜(Vogel & Martens, 2000), 对蕈蚊的吸引主要依靠其附属器散发的挥发性化学气味(Suetsugu et al, 2022), 包括拟态蘑菇的形态、颜色和气味来吸引以蘑菇为食或作为交配地、产卵地的蕈蚊(Vogel & Martens, 2000); 亦或是释放类似雌性昆虫性信息素(如正十七烷)气味吸引雄性昆虫前来交配(Kostelc et al, 1980; Andreadis et al, 2015)。然而, 一把伞南星雌雄个体间传粉者访问频率的差异是否与其挥发性气味的性别二态性有关还有待于进一步探究。此外, 蕈蚊会食用花粉作为补充能量和繁殖的营养物质(Song et al, 2014), 一把伞南星的花部报酬仅为花粉, 这也可能是佛焰苞花序内含有花粉的雄性个体被访问更多的原因。
本文的其它图/表
|