
|
|
||
|
雌雄异株物种一把伞南星雌雄株空间分布及性别二态性
生物多样性
2024, 32 (6):
24084-.
DOI: 10.17520/biods.2024084
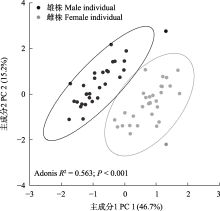
雌雄异株是有花植物雌雄性别功能在个体水平上由结合到分离的重要进化转变之一, 发生于被子植物的多个科属。雌雄异株物种的雌雄个体通常在分布、形态等方面存在差异。研究其雌雄个体的空间分布及性别二态性, 是理解雌雄异株物种繁殖策略及其适应与进化的重要前提。天南星属(Arisaema)是天南星科内已报道唯一存在雌雄异株且较为进化的类群, 一把伞南星(A. erubescens)是该属中不同性别个体存在差异的多年生草本植物。本文以湖南省八大公山国家级自然保护区内分布的一把伞南星为研究对象, 通过研究其雌雄个体的空间分布、花期物候、形态结构及访花者类群, 以期明确该物种雌雄植株的空间关联性以及花期物候、形态结构的性别二态性。结果表明: 一把伞南星群体内性比均呈极显著雄性偏倚, 雌雄个体在小尺度范围呈聚集分布, 但在较大尺度上呈随机分布; 雄性个体比雌性个体更早进入花期, 置换多元方差分析显示雌雄植株间形态特征存在显著差异, 其中假茎基径、佛焰苞管部直径、花序基径、花序柄直径等特征对性别二态性有重要贡献; 主要传粉者为白天活动的菌蚊科菌蚊属(Mycetophila)的蕈蚊, 其对雄株的访问频率极显著高于雌株。这些结果表明: 雌雄异株物种一把伞南星不同性别植株的空间关联与其传粉者蕈蚊的活动习性相匹配, 其开花物候、形态特征及传粉者访问等方面的性别二态性是对繁殖成功的保障。 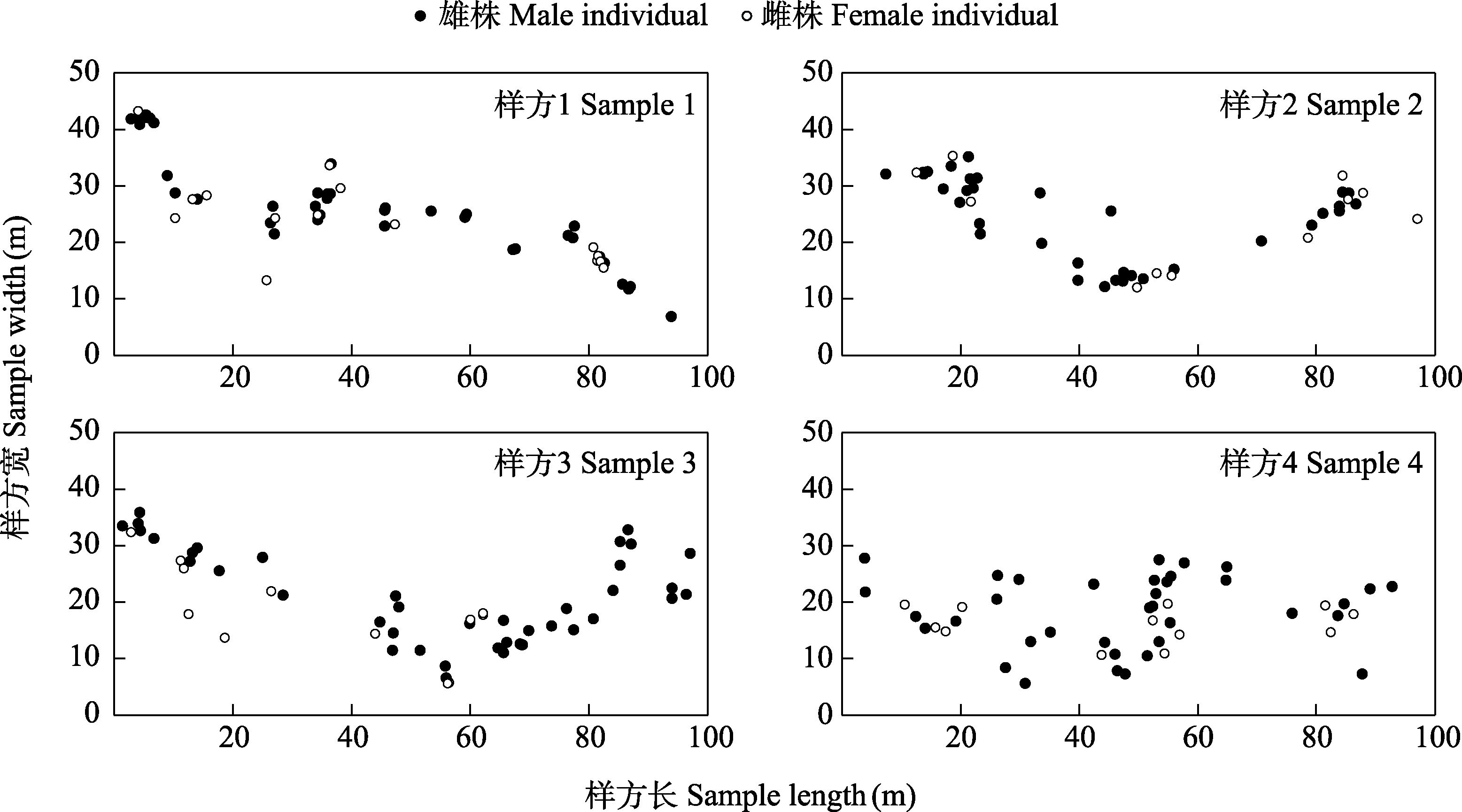 View image in article
图2
2022年八大公山国家级自然保护区一把伞南星在4个样方中的空间分布
正文中引用本图/表的段落
在2022年的花期结束后, 固定样方内共有有性植株78株(雄57株, 雌21株)。植株动态性比(雄 : 雌)随花期呈先小幅上升后下降的趋势, 种群进入花期后第10天性比最大(雄 : 雌 = 7.67), 后随雌株开放逐渐下降至最低0.33 (附录3)。4个样方(100 m × 50 m)中一把伞南星的最终性比均极显著偏离1 : 1, 平均性比(雄 : 雌)为3.01 ± 0.18, 表现出极显著的雄性性别偏倚(表1, 图2)。
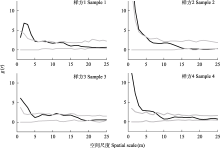
雌雄异株物种种群的性比通常偏离1 : 1的平衡预期(Sinclair et al, 2012)。本研究中一把伞南星2022年不同样方最终平均性比为3.01 ± 0.18, 均表现出极显著的雄性性别偏倚(表1, 图2), 这与天南星属其他物种一致(Richardson & Clay, 2001)。由于雌性的繁殖投入较高, 需通过减缓植株生长、推迟繁殖时间或降低开花频率等策略来权衡, 因此经过多次繁殖后性比呈现出雄性偏倚(Lloyd & Webb, 1977)。但存在放牧干扰的天南星属种群, 其性比则会受到严重影响。如在梅花鹿(Cervus nippon)密集食草的面积为0.36 ha的样地中, 尽管克隆繁殖的芦生天南星和无克隆繁殖的细齿南星并不可口, 但均未观察到两物种的雌性植株, 这可能与梅花鹿偏好啃食较大植株有关(Matsumoto et al, 2020)。此外, 上述两物种在放牧干扰下的空间聚集程度基本一致, 均呈现出小尺度内聚集分布(Matsumoto et al, 2020)。然而, 造成性比变化的同时也会导致雌雄异株植物种群中存在雌雄空间分隔(spatial segregation of the sexes), 据统计约20个科的30多个雌雄异株物种存在该现象(Barrett & Hough, 2013)。而干扰条件下样地内芦生天南星与细齿南星无雌株分布, 无法探讨雌雄植株间的空间关联性。一把伞南星具有与芦生天南星类似的无性繁殖能力(胡光万, 2008), 本研究4个样方中一把伞南星的雌雄植株空间关联虽有微小差别但总体趋势一致: 均在小尺度(5 m左右)呈聚集分布而在较大尺度(15 m以上)无相关, 呈随机分布(图3)。这可能与其传粉者为蕈蚊类群相适应, 虽然关于蕈蚊传粉距离没有明确的数据, 但Matsumoto等(2021)介绍: 细齿南星雌株和花粉供体间距离通常不超过44.3 m, 由此可见蕈蚊类传粉者携粉飞行距离较短, 因此一把伞南星较小尺度上的雌雄正相关有利于完成花粉传递促进繁殖成功。
本文的其它图/表
|


{kind=link}