|
|
||||||||||||||||||||||||||||||||||||
|
孑遗植物半日花叶际真菌群落的多样性与构建机制
生物多样性
2024, 32 (3):
23384-.
DOI: 10.17520/biods.2023384
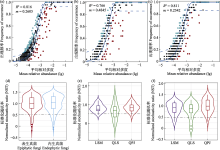
叶际微生物在维持植物健康生长方面具有重要的作用, 然而, 目前我们对珍稀孑遗植物半日花(Helianthemum songaricum)叶际真菌的多样性和群落结构等知之甚少。因此, 本研究利用Illumina高通量测序技术检测西鄂尔多斯自然保护区半日花叶片表生和内生真菌的多样性, 探究其网络结构特征以及群落构建的机制。结果显示: 棋盘井地区的叶际表生真菌丰富度指数(156.38 ± 8.42)显著高于内生真菌(111.13 ± 5.57), 棋盘井叶际表生真菌丰富度指数显著高于拉僧庙(125.57 ± 7.20)和千里山(114.75 ± 10.35), 而拉僧庙的内生真菌丰富度指数(155.71 ± 15.40)显著高于棋盘井。叶际真菌以子囊菌门、毛霉菌门和担子菌门为优势门, 叶际表生真菌和内生真菌分别在3个不同地点具有显著的指示类群, 叶片部位(即叶表和叶内)和地点显著影响叶片真菌的群落组成。共存网络分析表明, 叶际表生和内生真菌主要是协同作用, 拮抗作用较小。半日花叶际表生和内生真菌群落的构建主要由随机性过程驱动。综上所述, 半日花叶际表生和内生真菌的多样性和群落组成受到叶片部位(叶表和叶内)和地点的显著影响, 随机性过程主导叶际表生和内生真菌的群落构建。研究结果可为珍稀濒危植物的保护和合理利用提供一定的科学基础和实践指导。
表1
半日花叶际真菌网络拓扑参数
正文中引用本图/表的段落
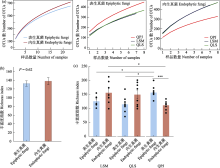
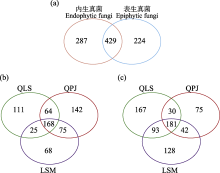
物种累积曲线显示, 无论是表生还是内生真菌的OTU数随着样品的增加均未达到平台期, 这意味着进一步采样会在一定程度上增加半日花叶际真菌的多样性(图1a)。此外, 单个样品的物种累积曲线分析表明, 不同样本的真菌多样性存在差异(附录1)。丰富度指数分析表明, 半日花的叶际内生真菌平均丰富度指数(平均值 ± 标准误: 137.96 ± 7.89)高于表生真菌(132.52 ± 6.08), 但无显著差异(P = 0.620, 图1b), 而棋盘井地区的叶际表生真菌(156.38 ± 8.42)显著高于内生真菌(111.13 ± 5.57, P < 0.001, 图1c)。在不同采样点之间叶际表生丰富度指数依次为棋盘井 > 拉僧庙 (125.57 ± 7.20) > 千里山(114.75 ± 10.35)。其中, 棋盘井地区的叶际表生真菌显著高于其他两个地区(P < 0.05, 图1c)。而叶际内生真菌的丰富度指数依次为拉僧庙(155.71 ± 15.40) > 千里山(149.25 ± 14.87) > 棋盘井。其中, 拉僧庙显著高于棋盘井地区(P = 0.021, 图1c)。并且, 基于OTU水平的Bray-Curtis距离进行NMDS排序分析表明, 叶际表生和内生真菌的群落组成显著不同(PerMANOVA, R2 = 0.13, P < 0.001, 图2a); PerMANOVA进一步统计分析发现, 不同地点的叶际表生真菌群落组成显著不同(R2 = 0.40, P < 0.001), 与此同时, 内生真菌在不同地点之间也具有显著差异(R2 = 0.22, P < 0.001, 图2b-c)。
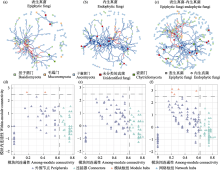
指示种分析表明, OTU_3 (线黑粉酵母属)、OTU_4 (Naganishia)、OTU_8 (放射毛霉属)等是千里山地区叶际表生真菌的指示类群; OTU_1 (放射毛霉属)、OTU_3、OTU_17 (Knufia)等是拉僧庙地区叶际表生真菌的指示类群; OTU_1、OTU_10 (Comoclathris)、OTU_13 (Bhatiellae)等是棋盘井地区叶际表生真菌的指示类群(图5a); OTU_8是千里山内生真菌指示类群; OTU_8、OTU_52 (Camarosporidiella)、OTU_123 (Comoclathris)是拉僧庙内生真菌指示类群; OTU_26 (Dimorpho- sporicola)、OTU_131 (Coniothyrium)、OTU_912 (葡萄座腔菌科Botryosphaeriaceae)是棋盘井内生真菌指示类群(图5b)。由此可见, 不同地区的半日花叶际表生真菌或内生真菌具有显著不同的指示类群, 且其指示类群的相对多度不同。例如, 叶际表生真菌指示类群OTU_1在拉僧庙和棋盘井的平均相对多度分别为35.56%、46.07% (图5a)。
利用共存网络(co-occurrence network)研究半日花叶际真菌群落的网络特征, 分别基于相对多度大于0.1%的表生和内生真菌构建了表生、内生和表生真菌-内生真菌共存网络(图6)。网络拓扑参数显示, 半日花叶际表生真菌网络由153个节点和282条边构成, 内生真菌网络是由214个节点和481条边构成, 表生真菌-内生真菌网络由357个节点和695条边构成, 且各网络的节点主要是子囊菌门、毛霉菌门和担子菌门组成(图6a-c)。网络的平均度和平均路径长度分析显示, 内生真菌网络的平均度高于表生真菌以及表生真菌-内生真菌网络; 而平均路径长度低于表生真菌和表生真菌-内生真菌网络, 其表生真菌-内生真菌网络的平均路径长度最高(表1)。这表明, 相比于叶际表生或表生真菌-内生真菌网络特征, 半日花内生真菌网络各成分互作程度和连接紧密度更高, 而表生真菌-内生真菌网络更加松散。表生和内生真菌网络的负相关边的比例分别为21.99%、4.78% (表1), 这显示半日花叶际表生真菌之间比内生真菌之间的竞争关系更加激烈。但网络中正相关边数均大于负相关边数, 表明叶际真菌群落之间的协同作用较大, 而拮抗作用较小。此外, 半日花叶际表生、内生和表生真菌-内生真菌网络的模块化指数分别为0.677、0.660和0.799 (表1), 具有高度模块化水平。
本文的其它图/表
|