|
|
||||||||||||||||||||||||||||||||||||||||||||||||||
|
基于3种线粒体标记探讨中日沿海角木叶鲽遗传多样性差异
生物多样性
2022, 30 (5):
21485-.
DOI: 10.17520/biods.2021485
角木叶鲽(Pleuronichthys cornutus)是东亚沿海重要的鲽形目经济鱼类, 为更好地保护和开发利用其种质资源, 有必要全面了解其遗传背景。本研究测定了中国和日本沿海7个群体200尾角木叶鲽线粒体控制区(CR) 5'端、细胞色素b (Cytb)和NADH脱氢酶第二亚基(ND2)基因序列, 比较不同标记在解析遗传多样性和种群结构上的可行性与有效性, 阐明中日沿海角木叶鲽群体间出现遗传分化的分子机制。CR序列分析发现中日沿海7个角木叶鲽群体遗传多样性表现出较高的单倍型多样性(Hd = 0.9699)和较低的核苷酸多样性(π = 0.0061); 各群体间无显著的遗传分化(FST = -0.0197-0.0184, P > 0.05); 单倍型网络未显示出明显的地理聚群和谱系结构; 分子方差分析(AMOVA)表明变异主要发生在群体内部(> 99.17%)。进一步通过Cytb和ND2基因分别与CR序列对比分析, 结果表明群体遗传多样性均表现为高Hd (0.9683-0.9829)低π (0.0050-0.0063)模式, 仅有ND2基因分析FST值(FST = 0.0302, P < 0.05)显示了中国碣石(GDJS)和日本明石(JAP)群体间显著的低水平遗传分化现象。CR、Cytb和ND2的单倍型网络图均无明显的地理聚类和谱系结构, AMOVA分析也显示变异主要来源于群体内(> 98.39%)。种群历史动态分析结果显示, 角木叶鲽可能在第四纪中更新世晚期经历了群体扩张事件, 扩张时间分别为31.93-9.58万年前(CR)、27.53-22.02万年前(Cytb)和26.99-18.75万年前(ND2)。综上所述, 中日沿海的角木叶鲽具有较高遗传多样性, GDJS和JAP群体间存在低度分化; ND2基因比CR和Cytb序列更适于分析角木叶鲽种群遗传结构, 选择多个遗传标记可有效弥补单一标记分析遗传多样性的局限性; 推测冰期两大独立避难所的形成及GDJS和JAP群体距离相隔较远是其发生遗传分化的主要原因。研究结果为中日沿海角木叶鲽渔业资源的种质保护与可持续利用提供了理论依据。
表5
基于CR、Cytb和ND2序列分析角木叶鲽群体的样本信息和遗传多样性
正文中引用本图/表的段落
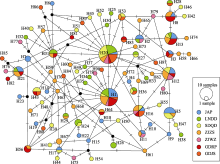
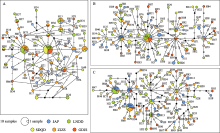
从中日海域中选择5个代表群体(样本信息见表5), 以分析3种分子标记应用于中日沿海角木叶鲽群体的遗传学差异。分析角木叶鲽CR序列的140个549 bp序列中, 存在2个碱基的插入或缺失。Cytb的140条序列全长为1,008 bp, ND2的140条序列长度为1,044 bp, 均不存在碱基的插入或缺失。三者碱基A+ T含量(53.8%-64.9%)高于碱基G + C含量, TS/TV均大于2.0, 符合mtDNA进化过程中转换频率高于颠换的规律, 适于进行遗传分析(Tarallo et al, 2016)。
如表5所示, 在不考虑插入、缺失的条件下角木叶鲽CR序列被检测到35个变异位点, 其中有22个简约信息位点。CR序列变异位点的数量定义了74个单倍型, 并经分析得出总体Hd为0.9683, π为0.0063。在Cytb和ND2基因序列中则分别检测到103个和112个变异位点, 分别定义了95种和94种单倍型, 其中简约信息位点分别为47个和51个。与CR基因相比, Cytb和ND2基因总体Hd值相近, 二者π值(0.0051, 0.0050)则略低于CR的π值, 即基于3种基因分析的5个角木叶鲽群体的遗传多样性整体分布模式均为高Hd低π, 表明中日沿海角木叶鲽群体遗传多样性较高。
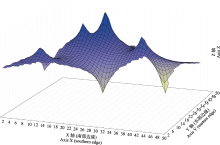
根据表5中的5个角木叶鲽群体的经纬度信息(表1)和ND2基因序列, 在AIS 1.0软件中进行Mantel Test检测角木叶鲽各群体间的地理距离与遗传分化值的相关程度, 并构建遗传距离空间分布图。Mantel Test结果显示遗传距离和地理距离不具有显著相关性(r = -0127, P > 0.05, 1,000个排列)。从遗传距离空间分布图(图3)可看出沿南部边缘从东到西, 存在一个较为平缓的曲面, 表明基因流动的障碍较低; 同时可见4处明显上升的尖峰, 但群体间变化幅度相对小, 其分布没有明确的空间模式。Mantel Test分析结果支持ND2基因的遗传距离空间分布图和成对FST值(表6), 即群体间遗传分化和地理距离之间没有显著相关性。结合AMOVA分析(表7)表明种群的变异主要发生在群体内, 因此中国碣石群体和日本明石群体间的遗传分化是微弱且不稳定的。
基于CR和Cytb基因分析5个角木叶鲽群体的遗传结构, 结果均显示无遗传分化、无地理聚群和谱系结构及变异主要发生在群体内部, 表明这两种基因应用于5个角木叶鲽遗传结构分析的结果差异较小。但基于ND2基因的分析揭示了角木叶鲽中国广东碣石群体和日本明石群体间低水平的遗传分化关系, 这为角木叶鲽的种群遗传结构补充了新的内容。但这种遗传分化水平较低且不稳定, 这可能是种群历史阶段所形成的群体间遗传分化被具有高扩散能力的庞大种群规模形成的基因频繁交流长期淡化所致。综上结果, 角木叶鲽的中国碣石群体和日本明石群体间存在显著的低水平遗传分化。在3种线粒体分子标记对比分析中国和日本沿海的角木叶鲽群体遗传多样性中, 角木叶鲽的ND2基因比CR、Cytb序列更适于进行该种群的遗传结构分析。
本文的其它图/表
|
||||||||||||||||||||||||||||||||||||||||||||||||||
备案号:京ICP备16067583号-7
Copyright © 2026 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn
![]()