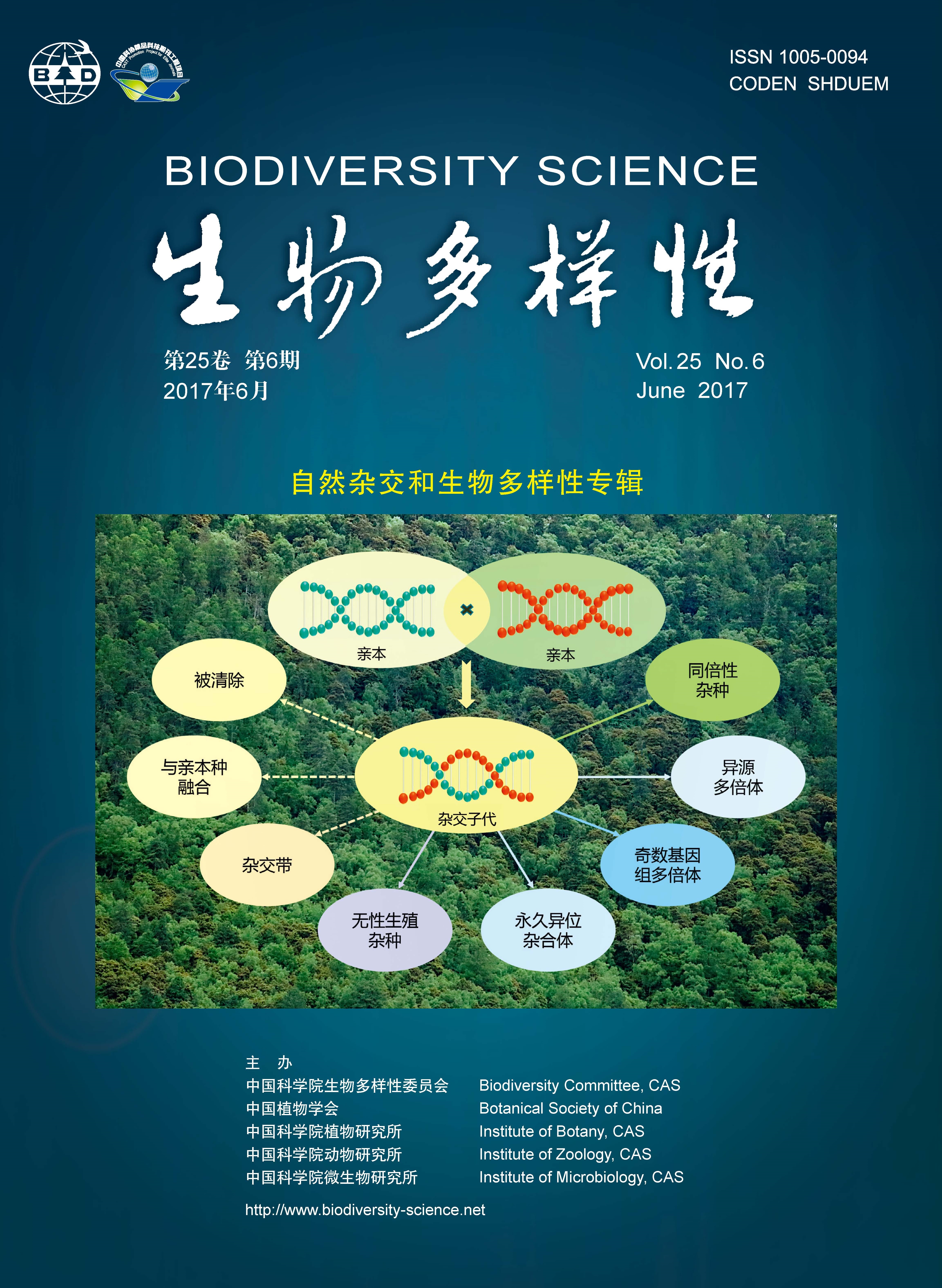
自然杂交在植物界中十分普遍, 一直是物种形成和进化研究的热点。秋海棠属(Begonia)为世界最大植物属之一, 其种类繁多, 但关于自然杂交的相关报道很少。系统调查秋海棠属的自然杂交不仅有利于理解该属植物的多样性, 也是弄清其资源和解决物种分类问题不可缺少的环节。本文通过对国产秋海棠属植物自然杂交现象的系统调查和分析, 发现总计29种(占国产已知200种秋海棠的约15%)参与了杂交, 产生了31个自然“杂种”约50个居群。其中, 掌叶秋海棠(B. hemsleyana)和粗喙秋海棠(B. longifolia)参与杂交程度最高, 分别与另外的8种和7种秋海棠发生了杂交; 而裂叶秋海棠(B. palmata)发生杂交的居群最多, 达16处。自然杂交以云南(尤其是南部)发生频率最高, 一共有20种参与, 产生了31处杂交居群; 台湾和广西次之。杂交多为单向发生, 个体以F1代个体为主, 尚未脱离亲本独立成种。野外传粉生物学初步观察发现: 秋海棠的主要访花昆虫为食蚜蝇类, 其次是蜜蜂科, 但各自传粉特点及效率尚待进一步研究。杂种及疑似亲本的鉴定可通过形态学、分子生物学和人工重复杂交等综合方法实现。杂种的基因组大小(C值)等于或接近两亲本的平均值, 误差通常小于10%, 有助于辅助鉴定杂种及亲本。我们总结后认为杂交发生及杂种形成必须满足5个条件: (1)重叠或邻近分布; (2)花期相遇; (3)有效传粉媒介; (4)杂交亲和; (5)适宜种子萌发幼苗生长的小生境及气候条件。据此, 我们预测了国产秋海棠属未来可能发生的约40个新杂交组合的可能亲本及地点。到目前为止, 尽管所有已知的秋海棠杂种尚未形成真正的新物种, 但是, 为了满足科学研究及实践交流的需要, 建议按照新分类群发表的严格要求, 将“杂种”逐一开展调查研究并整理发表, 并将其作为物种进化过程中的一个特殊分类单元对待。鉴于秋海棠属的杂种个体稀少且并未对亲本构成威胁, 反而在一定程度上增加了类群的多样性, 并为优良观赏品种的直接选育创造了机会, 因此, 国产秋海棠属种类丰富及杂种区应为就地保护的优先考虑范围之一。同时, 鉴于秋海棠属杂交亲和性高, 迁地保护栽培时应增大花期重叠种类的种植间距, 避免发生自然杂交。