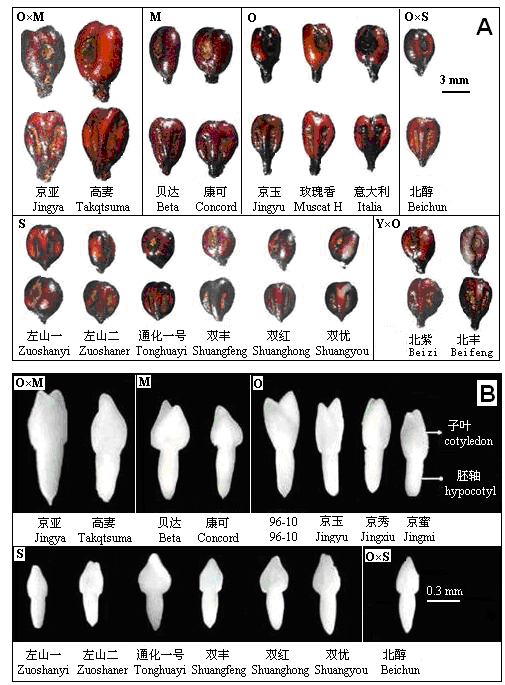
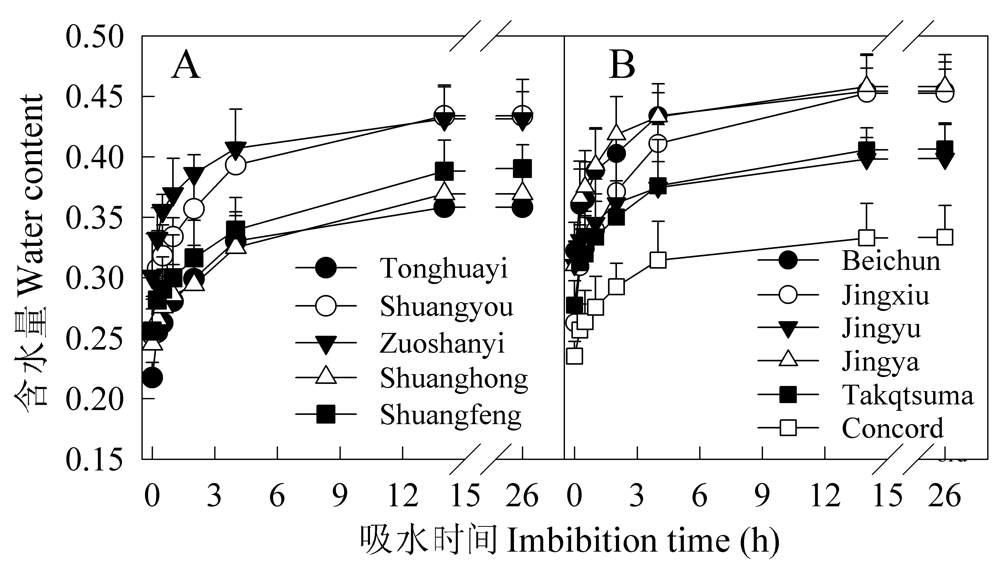
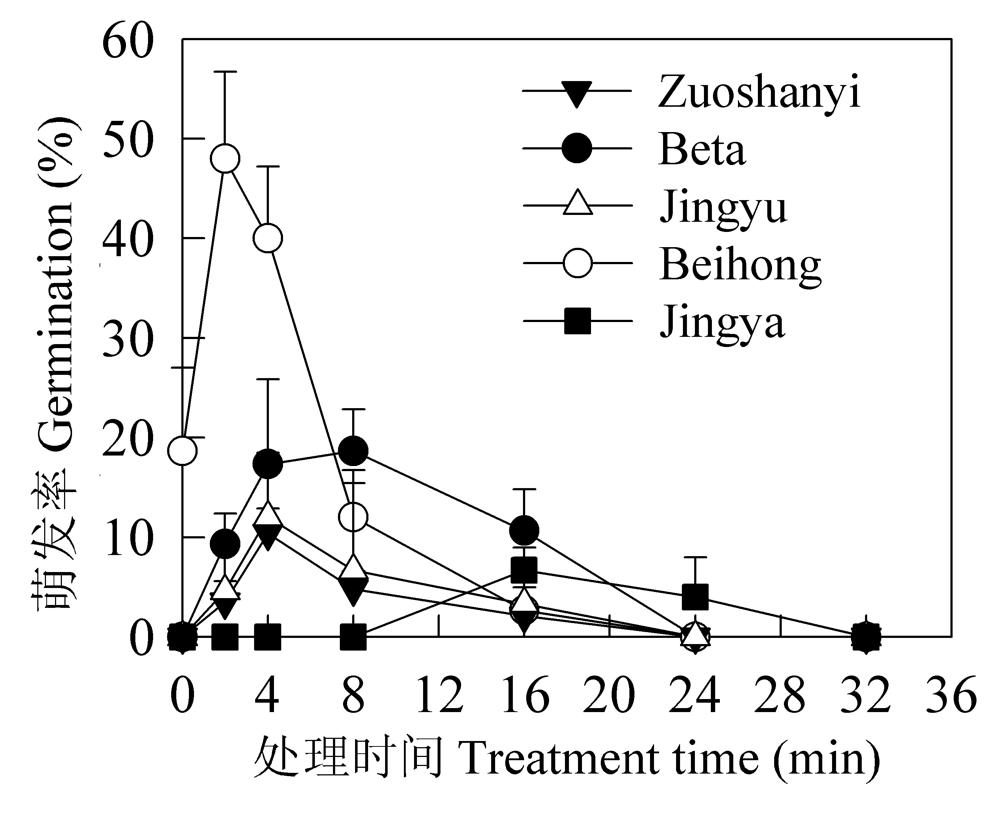
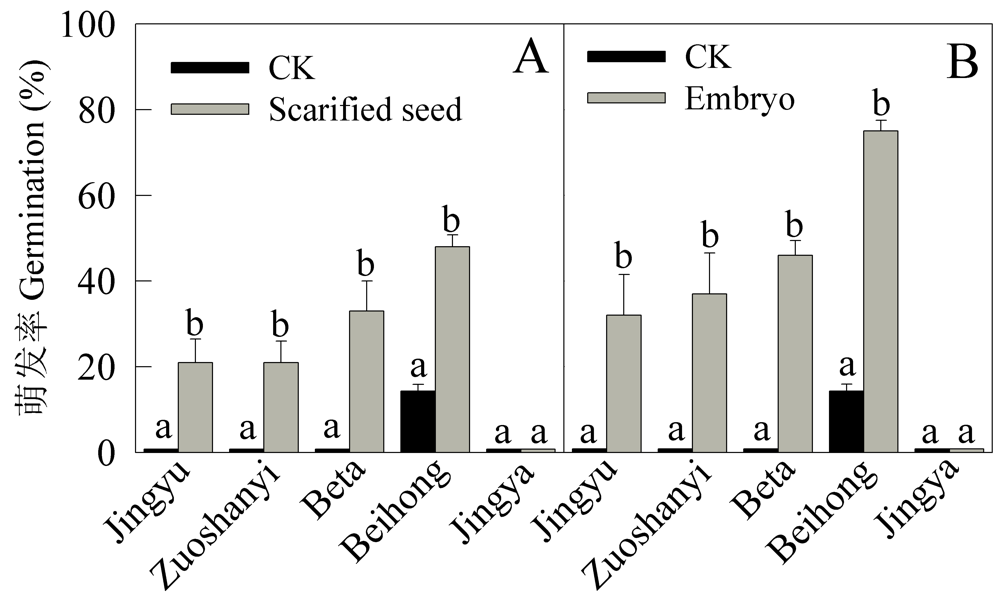
| [1] |
Alleweldt G, Speigel-Roy P, Reisch B (1990) Grapes (Vitis) genetic resources of temperate fruit and nut crops. Acta Horticulturae, 273,291-327.
|
| [2] |
Baskin JM, Baskin CC (2004) A classification system for seed dormancy. Seed Science Research, 14,1-16.
DOI
URL
|
| [3] |
Ellis RH, Hong D, Roberts EH (1983) A note on the development of a practical procedure for promoting the germination of dormant seed of grape (Vitis spp.). Vitis, 22,211-219.
|
| [4] |
Fernandez GE, Clark JR, Moore JN (1991) Effect of seed coat manipulation on the germination of stenospermocarpic grape embryos cultured in ovule. HortScience, 26,1220.
DOI
URL
|
| [5] |
He PC (贺普超) (1999) Taxonomy and resources of grapes (Vitis). In: Vitis (葡萄学)(ed. He PC (贺普超)), pp.8-10. China Agriculture Press, Beijing. (in Chinese)
|
| [6] |
Jaillon O, Aury JM, Noel B, Policriti A, Clepet C, Casagrande A, Choisne N, Aubourg S, Vitulo N, Jubin C, Vezzi A, Legeai F, Hugueney P, Dasilva C, Horner D, Mica E, Jublot D, Poulain J, Bruyère C, Billault A, Segurens B, Gouyvenoux M, Ugarte E, Cattonaro F, Anthouard V, Vico V, Del Fabbro C, Alaux M, Di Gaspero G, Dumas V, Felice N, Paillard S, Juman I, Moroldo M, Scalabrin S, Canaguier A, Le Clainche I, Malacrida G, Durand E, Pesole G, Laucou V, Chatelet P, Merdinoglu D, Delledonne M, Pezzotti M, Lecharny A, Scarpelli C, Artiguenave F, Pè ME, Valle G, Morgante M, Caboche M, Adam-Blondon AF, Weissenbach J, Quétier F, Wincker P (2007) The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature, 449,463-467.
DOI
URL
PMID
|
| [7] |
Kong QS (孔庆山) (2004) Chinese Vitis (中国葡萄志), pp.16-18. China Agricultural Science and Technology Press, Beijing. (in Chinese)
|
| [8] |
Kucera B, Cohn MA, Leubner-Metzger G (2005) Plant hormone interaction during seed dormancy release and germination. Seed Science Research, 15,281-307.
|
| [9] |
Mao BR (毛伯韧), Wu ZS (吴兆苏) (1983) Study on the inheritance and mechanism of seed dormancy in wheat. Scientia Agricultura Sinica (中国农业科学), 16(6),53-60. (in Chinese with English abstract)
|
| [10] |
Nikolaeva MG (2004) On criteria to use in studies of seed evolution. Seed Science Research, 14,315-320.
|
| [11] |
Rajasekaran K, Vine J, Mullins MG (1982) Dormancy in somatic embryos and seeds of Vitis: changes in endogenous abscisic acid during embryogeny and germination. Planta, 154,139-144.
URL
PMID
|
| [12] |
Selim HH, Ibrahim FA, Fayek MA, Sari EI-Deen SA, Gama NM (1981) Effect of different treatments on germination of Romi red grape seeds. Vitis, 20,115-121.
|
| [13] |
Spiegel-Roy P, Shulman Y, Baron I, Ashbel E (1987) Effect of cyanamide in overcoming grape seed dormancy. HortScience, 22,208-210.
|
| [14] |
Thompson K, Ceriani RM, Bakker JP, Bekker RM (2003) Are seed dormancy and persistence in soil related? Seed Science Research, 13,97-100.
|
| [15] |
Zheng GH (郑光华) (2004) Seed Physiology Research (种子生理研究), pp.321-322. Science Press, Beijing. (in Chinese)
|

 )
)