
生物多样性 ›› 2014, Vol. 22 ›› Issue (4): 458-466. DOI: 10.3724/SP.J.1003.2014.13232 cstr: 32101.14.SP.J.1003.2014.13232
庄平*( )
)
收稿日期:2013-10-28
接受日期:2014-02-26
出版日期:2014-07-20
发布日期:2014-07-24
通讯作者:
庄平
基金资助:
Ping Zhuang*()
Received:2013-10-28
Accepted:2014-02-26
Online:2014-07-20
Published:2014-07-24
Contact:
Zhuang Ping
摘要:
目前有研究表明被子植物类群中单一的开花或展叶物候模式可能具有遗传性, 但两类物候事件的节律关系是否受遗传因素控制, 各种节律模式的形成与演化机制及其与生物系统演化的关系仍不清楚。作者对四川都江堰市迁地保育的2个海拔点的杜鹃属(Rhododendron)4亚属4组13亚组42种植物的开花-展叶节律模式进行了5年的观察研究。作者假定杜鹃属的开花-展叶节律模式为进化遗传特性, 包括先花后叶(F→L)、先花后叶部分重叠(F+L)、叶期包花期(L∈F)、先叶后花部分重叠(L+F)和先叶后花(L→F)等5种类型。同一地点不同年份以及不同地点同一物种的相关物候观察结果均显示, 尽管它们5年的物候发生时间随年度和观察点海拔不同而有波动, 但物候发生次序和开花-展叶节律模式均未见随年份而变化, 即每个物种只对应一种节律模式, 支持了上述假设。作者进一步提出了开花-展叶节律模式与物种的系统演化存在进化程度上的差异和序列关系的假设。不同节律模式的物种其区系地理以及不同海拔和分类群中的物种数量分布特征表明: (1)进化程度不同的节律模式分别对应于古老的中国-日本森林植物亚区和年轻的中国-喜玛拉雅森林植物亚区物种, 较原始的节律模式倾向于出现在中高山海拔区域, 并与杜鹃属可能起源山地的一定海拔区域构成关联, 而进化程度较高的节律模式倾向于出现在更高或更低的海拔地段; (2)节律模式的进化程度与有关亚属、组、亚组的系统位置和进化程度有一定关联; (3)节律模式的演化与生活型、叶性进化同步, 环境胁迫、生长期长度变化、光资源竞争等都可能是节律模式演化的外部动力。研究还表明, 在气候波动条件下物候事件间的发生次序比单一物候事件的时间属性更加稳定。
庄平 (2014) 四川都江堰迁地保育的42种杜鹃属植物开花-展叶物候节律. 生物多样性, 22, 458-466. DOI: 10.3724/SP.J.1003.2014.13232.
Ping Zhuang (2014) Analysis of the flowering-leafing phenorhythm of 42 Rhododendron species conserved ex situ in Dujiangyan, Sichuan Province, China. Biodiversity Science, 22, 458-466. DOI: 10.3724/SP.J.1003.2014.13232.
| 地理区系 Geographic flora | 观察种数 No. of species | F→L | F+L | L∈F | L+F | L→F | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | % | N | % | N | % | N | % | N | % | ||||||
| 中国-日本亚区 Sino-Japan subregion | 18 | 10 | 55.6 | 2 | 11.1 | 3 | 16.7 | 3 | 16.7 | - | - | ||||
| 常绿杜鹃亚属 Subgen. Hymenanthes | 11 | 7 | 63.6 | 2 | 18.2 | 2 | 18.2 | 1 | 9.1 | - | - | ||||
| 杜鹃亚属 Subgen. Rhododendron | 5 | 3 | 60.0 | - | - | 1 | 20.0 | 1 | 20.0 | - | - | ||||
| 中国-喜玛拉雅亚区 Sino-Himalaya subregion | 12 | 1 | 8.3 | 4 | 33.3 | 2 | 16.7 | 3 | 25.0 | 2 | 16.7 | ||||
| 常绿杜鹃亚属 Subgen. Hymenanthes | 4 | - | - | 2 | 50.0 | 1 | 25.0 | - | - | 1 | 25.0 | ||||
| 杜鹃亚属 Subgen. Rhododendron | 8 | 1 | 12.5 | 2 | 25.0 | 1 | 12.5 | 3 | 37.5 | 1 | 12.5 | ||||
| 两亚区共有 Common species between the two subregions | 12 | 4 | 33.3 | 1 | 8.3 | 3 | 25.0 | 2 | 16.7 | 2 | 16.7 | ||||
| 常绿杜鹃亚属 Subgen. Hymenanthes | 5 | 2 | 40.0 | - | - | 2 | 40.0 | 1 | 20.0 | - | - | ||||
| 杜鹃亚属 Subgen. Rhododendron | 6 | 2 | 33.3 | 1 | 16.7 | 1 | 16.7 | 1 | 16.7 | 1 | 16.7 | ||||
表1 42种杜鹃属植物开花-展叶节律模式与地理区系成分的关系
Table 1 Relationship between flowering-leafing phenorhythm type and the area-type of 42 Rhododendron species
| 地理区系 Geographic flora | 观察种数 No. of species | F→L | F+L | L∈F | L+F | L→F | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | % | N | % | N | % | N | % | N | % | ||||||
| 中国-日本亚区 Sino-Japan subregion | 18 | 10 | 55.6 | 2 | 11.1 | 3 | 16.7 | 3 | 16.7 | - | - | ||||
| 常绿杜鹃亚属 Subgen. Hymenanthes | 11 | 7 | 63.6 | 2 | 18.2 | 2 | 18.2 | 1 | 9.1 | - | - | ||||
| 杜鹃亚属 Subgen. Rhododendron | 5 | 3 | 60.0 | - | - | 1 | 20.0 | 1 | 20.0 | - | - | ||||
| 中国-喜玛拉雅亚区 Sino-Himalaya subregion | 12 | 1 | 8.3 | 4 | 33.3 | 2 | 16.7 | 3 | 25.0 | 2 | 16.7 | ||||
| 常绿杜鹃亚属 Subgen. Hymenanthes | 4 | - | - | 2 | 50.0 | 1 | 25.0 | - | - | 1 | 25.0 | ||||
| 杜鹃亚属 Subgen. Rhododendron | 8 | 1 | 12.5 | 2 | 25.0 | 1 | 12.5 | 3 | 37.5 | 1 | 12.5 | ||||
| 两亚区共有 Common species between the two subregions | 12 | 4 | 33.3 | 1 | 8.3 | 3 | 25.0 | 2 | 16.7 | 2 | 16.7 | ||||
| 常绿杜鹃亚属 Subgen. Hymenanthes | 5 | 2 | 40.0 | - | - | 2 | 40.0 | 1 | 20.0 | - | - | ||||
| 杜鹃亚属 Subgen. Rhododendron | 6 | 2 | 33.3 | 1 | 16.7 | 1 | 16.7 | 1 | 16.7 | 1 | 16.7 | ||||
| 矫正海拔区间(m) Rectified elevation range | 观察种数 No. of species | F→L | F+L | L∈F | L+F | L→F | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | % | N | % | N | % | N | % | N | % | ||||||
| <1,500 | 5 | 2 | 40.0 | - | - | 1 | 20.0 | 1 | 20.0 | 1 | 20.0 | ||||
| 1,500-2,000 | 8 | 4 | 50.0 | 1 | 12.5 | 2 | 25.0 | 1 | 12.5 | - | - | ||||
| 2,000-2,500 | 12 | 6 | 50.0 | 2 | 16.7 | 1 | 8.3 | 2 | 16.7 | 1 | 8.3 | ||||
| 2,500-3,000 | 12 | 3 | 25.0 | 3 | 25.0 | 2 | 16.7 | 3 | 25.0 | 1 | 8.3 | ||||
| >3,000 | 5 | - | - | 1 | 20.0 | 2 | 40.0 | 1 | 20.0 | 1 | 20.0 | ||||
表2 42种杜鹃属植物开花-展叶节律模式与海拔分布的关系
Table 2 Relationship between the flowering-leafing phenorhythm type and the elevation range of 42 Rhododendron species
| 矫正海拔区间(m) Rectified elevation range | 观察种数 No. of species | F→L | F+L | L∈F | L+F | L→F | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | % | N | % | N | % | N | % | N | % | ||||||
| <1,500 | 5 | 2 | 40.0 | - | - | 1 | 20.0 | 1 | 20.0 | 1 | 20.0 | ||||
| 1,500-2,000 | 8 | 4 | 50.0 | 1 | 12.5 | 2 | 25.0 | 1 | 12.5 | - | - | ||||
| 2,000-2,500 | 12 | 6 | 50.0 | 2 | 16.7 | 1 | 8.3 | 2 | 16.7 | 1 | 8.3 | ||||
| 2,500-3,000 | 12 | 3 | 25.0 | 3 | 25.0 | 2 | 16.7 | 3 | 25.0 | 1 | 8.3 | ||||
| >3,000 | 5 | - | - | 1 | 20.0 | 2 | 40.0 | 1 | 20.0 | 1 | 20.0 | ||||
| 类群 Group | 观察种数 No. of species | F→L | F+L | L∈F | L+F | L→F | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | % | N | % | N | % | N | % | N | % | ||||||
| 常绿杜鹃亚属 Subgen. Hymenanthes | 21 | 9 | 42.9 | 4 | 19.1 | 5 | 23.8 | 2 | 9.5 | 1 | 4.8 | ||||
| 云锦杜鹃亚组 Subsect. Fortunea | 6 | 4 | 66.7 | - | - | 1 | 17.5 | 1 | 17.5 | - | - | ||||
| 银叶杜鹃亚组 Subsect. Argyrophylla | 5 | 3 | 60.0 | 2 | 40.0 | - | - | - | - | - | - | ||||
| 麻花杜鹃亚组 Subsect. Maculifera | 3 | 1 | 33.3 | - | - | 1 | 33.3 | 1 | 33.3 | ||||||
| 杜鹃亚属 Subgen. Rhododendron | 18 | 6 | 33.3 | 3 | 16.7 | 3 | 16.7 | 5 | 27.8 | 1 | 5.6 | ||||
| 三花杜鹃亚组 Subsect. Triflora | 13 | 2 | 15.4 | 3 | 23.1 | 2 | 15.4 | 5 | 38.6 | 1 | 7.7 | ||||
| 映山红亚属 Subgen. Tsutsusi | 2 | - | - | - | - | - | - | 1 | 50.0 | 1 | 50.0 | ||||
| 羊踯躅亚属 Subgen. Pentanthera | 1 | - | - | - | - | - | - | - | - | 1 | 100.0 | ||||
| 合计 Total | 42 | 15 | 35.7 | 7 | 21.4 | 8 | 19.1 | 8 | 19.1 | 4 | 9.5 | ||||
表3 42种杜鹃属开花-展叶节律模式与系统发育的关系
Table 3 Relationship between the flowering-leafing phenorhythm type and phylogeny of 42 Rhododendron species
| 类群 Group | 观察种数 No. of species | F→L | F+L | L∈F | L+F | L→F | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | % | N | % | N | % | N | % | N | % | ||||||
| 常绿杜鹃亚属 Subgen. Hymenanthes | 21 | 9 | 42.9 | 4 | 19.1 | 5 | 23.8 | 2 | 9.5 | 1 | 4.8 | ||||
| 云锦杜鹃亚组 Subsect. Fortunea | 6 | 4 | 66.7 | - | - | 1 | 17.5 | 1 | 17.5 | - | - | ||||
| 银叶杜鹃亚组 Subsect. Argyrophylla | 5 | 3 | 60.0 | 2 | 40.0 | - | - | - | - | - | - | ||||
| 麻花杜鹃亚组 Subsect. Maculifera | 3 | 1 | 33.3 | - | - | 1 | 33.3 | 1 | 33.3 | ||||||
| 杜鹃亚属 Subgen. Rhododendron | 18 | 6 | 33.3 | 3 | 16.7 | 3 | 16.7 | 5 | 27.8 | 1 | 5.6 | ||||
| 三花杜鹃亚组 Subsect. Triflora | 13 | 2 | 15.4 | 3 | 23.1 | 2 | 15.4 | 5 | 38.6 | 1 | 7.7 | ||||
| 映山红亚属 Subgen. Tsutsusi | 2 | - | - | - | - | - | - | 1 | 50.0 | 1 | 50.0 | ||||
| 羊踯躅亚属 Subgen. Pentanthera | 1 | - | - | - | - | - | - | - | - | 1 | 100.0 | ||||
| 合计 Total | 42 | 15 | 35.7 | 7 | 21.4 | 8 | 19.1 | 8 | 19.1 | 4 | 9.5 | ||||
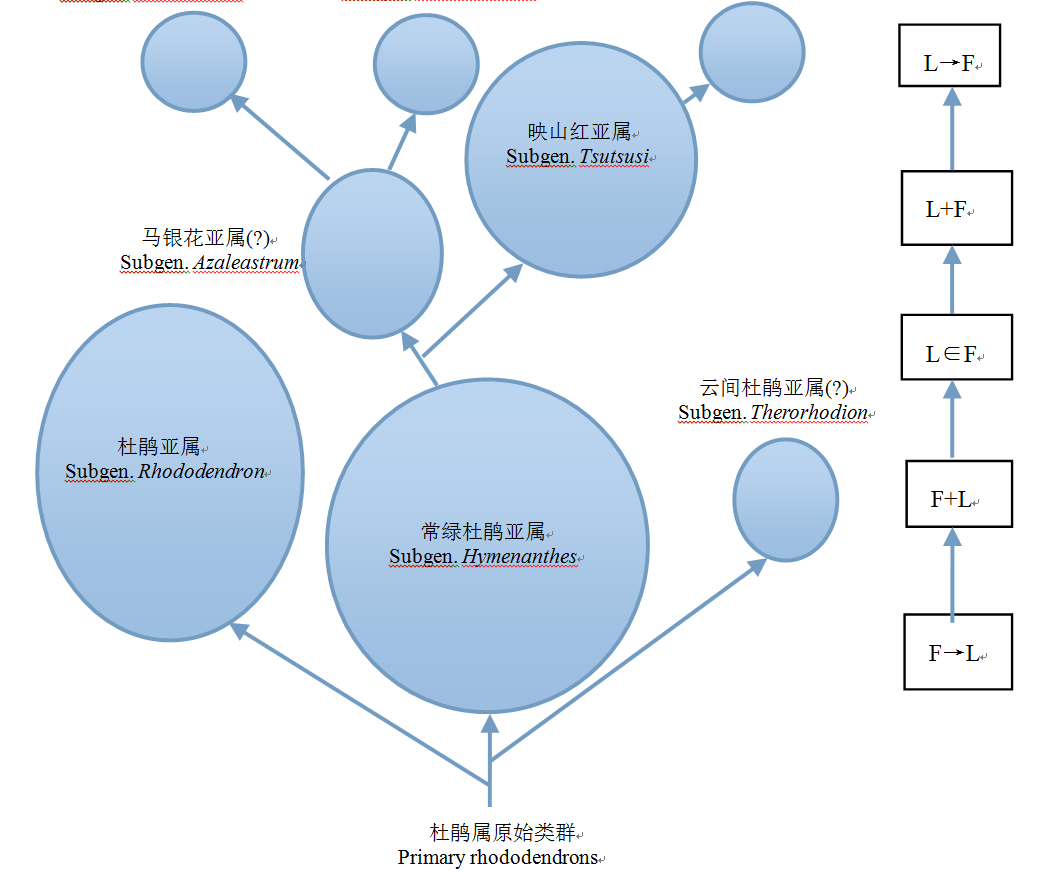
图1 杜鹃属植物亚属间关系及其花叶物候节律模式的演化方向。各节律模式的含义见表1。“?”表示相应亚属的花叶节律尚不清楚。
Fig. 1 Phylogenetic relationship of Rhododendron subgenera and evolutional sequence of their flowering-leafing phenorhythm type. The meaning of each phenorhythm type shows in Table 1. “?” shows the flowering-leafing phenorhythm type is not clear in the subgenera.
| 花叶节律模式 Phenorhythmotype | 种类 Species | 来源 Source | 海拔 Elevation (m) | 开花展叶物候时间 Phenologic timing of flowering and leafing① | ||||
|---|---|---|---|---|---|---|---|---|
| 3月 March | 4月Apr. | 5月May | 6月June | 7月July | ||||
| F→L: 先花后叶型Flowering before leafing | 美容杜鹃 R. calophytum | 龙池 Longchi | 2,000 | ○○○○ △△△△△△ | ||||
| 腺果杜鹃 R. davidii | 龙池 Longchi | 1,800 | ○○○○△△△△ | |||||
| 山光杜鹃 R. oreodoxa | 龙池 Longchi | 2,500 | ○○○○○ △△△ | |||||
| 团叶杜鹃 R. orbiculare | 汶川 Wenchuan | 2,500 | ○○○ △△△△△ | |||||
| 大王杜鹃 R. rex | 椅子丫口 Yiziyakou | 2,500 | ○○○○ △△△△△ | |||||
| 紫斑杜鹃 R. strigillosum var. monosematum | 峨眉山 Emeishan | 2,500 | ○○○○△△△△△ | |||||
| 岷江杜鹃 R. hunnewellianum | 龙池 Longchi | 1,400 | ○○○○○○△△△ | |||||
| 海绵杜鹃 R. pingianum | 峨眉山 Emeishan | 2,400 | ○○○△△△ | |||||
| 繁花杜鹃 R. floribundum | 龙肘山 Longzhoushan | 2,900 | ○○○ △△△△△△△△ | |||||
| 百合花杜鹃 R .liliiflorum | 庐山 Lushan | 1,300 | ○○○○△△△△△ | |||||
| 多鳞杜鹃 R. polylepis | 龙池 Longchi | 1,700 | ○○○○△△△△△ | |||||
| 黄花杜鹃 R. lutescensx | 龙池 Longchi | 1,700 | ○○○△△△ | |||||
| 红棕杜鹃 R. rubiginosum | 木里 Muli | 3,200 | ○○○△△△△ | |||||
| 柔毛碎米花 R. mollicomum | 战河 Zhanhe | 3,200 | ○○○△△△△ | |||||
| 腋花杜鹃 R. racemosum | 战河 Zhanhe | 2,700 | ○○○△△△△ | |||||
| F+L: 先花后叶、部分重叠型 Flowering ahead and covering part time of leafing | 窄叶杜鹃 R. araiophyllum | 新主 Xinzhu | 2,600 | ○◎◎△ | ||||
| 银叶杜鹃 R. argyrophyllum | 峨眉山 Emeishan | 2,200 | ○○◎◎△ | |||||
| 峨嵋银叶杜鹃 R. argyrophylum subsp. omeiense | 龙池 Longchi | 1,800 | ○○◎◎△△ | |||||
| 巴郎杜鹃 R. balangense | 卧龙 Wolong | 2,700 | ○◎◎△ | |||||
| 基毛杜鹃 R. rigidum | 木里 Muli | 3,200 | ○◎◎△△ | |||||
| 紫花杜鹃 R. amesiae | 海螺沟 Hailuogou | 3,500 | ○◎◎△ | |||||
| 张口杜鹃 R. augustinii subsp. chasmanthum | 片马 Pianma | 3,100 | ○◎◎◎△△ | |||||
| L∈F: 叶期包花期型 Leafing covers flowering , | 喇叭杜鹃 R. discolor | 龙池 Longchi | 1,600 | △◎◎◎△ | ||||
| 绒毛杜鹃 R. pachytrichum | 海螺沟 Hailuogou | 3,200 | △◎◎◎△ | |||||
| 马缨杜鹃 R. delavayi | 大方 Dafang | 1,700 | △△△◎◎◎△ | |||||
| 皱皮杜鹃 R. wiltonii | 峨眉山 Emeishan | 2,400 | △◎◎◎△ | |||||
| 毛肋杜鹃 R. augustinii | 龙池 Longchi | 2,000 | △△◎◎◎△ | |||||
| 云南杜鹃 R. yunnanense | 玉龙雪山 Yulongxueshan | 3,600 | △◎◎◎△ | |||||
| 硬毛杜鹃 R. hirtipes | 多雄拉 Duoxiongla | 3,500 | △◎◎◎△△△△△△△△△△△△△△ | |||||
| 亮鳞杜鹃 R. heliolepis | 泸定 Luding | 3,100 | △△△◎◎◎△ | |||||
| L+F: 先叶后花、部分重叠型 Leafing ahead and covering part time of flowering | 大白杜鹃 R. decorum | 泸定 Luding | 1,800 | △◎◎○○ | ||||
| 硬叶杜鹃 R. tatsienense | 片马 Pianma | 3,200 | △△◎◎○ | |||||
| 长毛杜鹃 R. trichanthum | 川西 Chuanxi | 2,200 | △△◎○○ | |||||
| 问客杜鹃 R. ambiguum | 龙池 Longchi | 2,400 | △◎◎◎○ | |||||
| 山育杜鹃 R. oreotrephes | 泸定 Luding | 3,000 | △△◎◎○○ | |||||
| 三花杜鹃 R. triflorum | 多雄拉 Duoxiongla | 3,000 | △△△◎○○ | |||||
| 映山红 R. simsii | 大方 Dafang | 1,700 | △△△◎◎○ | |||||
| 长鳞杜鹃 R. longesquamatum | 龙池 Longchi | 2,700 | △△◎○○ | |||||
| L→F: 先叶后花型Leafing before flowering | 云雾杜鹃 R. chamaethomsonii | 多雄拉 Duoxiongla | 4,000 | △△△ ○○○ | ||||
| 白面杜鹃 R. zaleucum | 玉龙雪山 Yulongxueshan | 3,400 | △△△○○○ | |||||
| 亮毛杜鹃 R. microphyton | 无量山 Wuliangshan | 2,700 | △△△△○○○ | |||||
| 羊踯躅 R. moll | 庐山 Lushan | 1,300 | △△△△○○○○ | |||||
附表1 都江堰迁地保护的42种杜鹃属植物开花-展叶节律模式
Table S1 Flowering-leafing phenorhythm type of 42 Rhododendron species conserved ex situ in Dujiangyan, Sichuan Province
| 花叶节律模式 Phenorhythmotype | 种类 Species | 来源 Source | 海拔 Elevation (m) | 开花展叶物候时间 Phenologic timing of flowering and leafing① | ||||
|---|---|---|---|---|---|---|---|---|
| 3月 March | 4月Apr. | 5月May | 6月June | 7月July | ||||
| F→L: 先花后叶型Flowering before leafing | 美容杜鹃 R. calophytum | 龙池 Longchi | 2,000 | ○○○○ △△△△△△ | ||||
| 腺果杜鹃 R. davidii | 龙池 Longchi | 1,800 | ○○○○△△△△ | |||||
| 山光杜鹃 R. oreodoxa | 龙池 Longchi | 2,500 | ○○○○○ △△△ | |||||
| 团叶杜鹃 R. orbiculare | 汶川 Wenchuan | 2,500 | ○○○ △△△△△ | |||||
| 大王杜鹃 R. rex | 椅子丫口 Yiziyakou | 2,500 | ○○○○ △△△△△ | |||||
| 紫斑杜鹃 R. strigillosum var. monosematum | 峨眉山 Emeishan | 2,500 | ○○○○△△△△△ | |||||
| 岷江杜鹃 R. hunnewellianum | 龙池 Longchi | 1,400 | ○○○○○○△△△ | |||||
| 海绵杜鹃 R. pingianum | 峨眉山 Emeishan | 2,400 | ○○○△△△ | |||||
| 繁花杜鹃 R. floribundum | 龙肘山 Longzhoushan | 2,900 | ○○○ △△△△△△△△ | |||||
| 百合花杜鹃 R .liliiflorum | 庐山 Lushan | 1,300 | ○○○○△△△△△ | |||||
| 多鳞杜鹃 R. polylepis | 龙池 Longchi | 1,700 | ○○○○△△△△△ | |||||
| 黄花杜鹃 R. lutescensx | 龙池 Longchi | 1,700 | ○○○△△△ | |||||
| 红棕杜鹃 R. rubiginosum | 木里 Muli | 3,200 | ○○○△△△△ | |||||
| 柔毛碎米花 R. mollicomum | 战河 Zhanhe | 3,200 | ○○○△△△△ | |||||
| 腋花杜鹃 R. racemosum | 战河 Zhanhe | 2,700 | ○○○△△△△ | |||||
| F+L: 先花后叶、部分重叠型 Flowering ahead and covering part time of leafing | 窄叶杜鹃 R. araiophyllum | 新主 Xinzhu | 2,600 | ○◎◎△ | ||||
| 银叶杜鹃 R. argyrophyllum | 峨眉山 Emeishan | 2,200 | ○○◎◎△ | |||||
| 峨嵋银叶杜鹃 R. argyrophylum subsp. omeiense | 龙池 Longchi | 1,800 | ○○◎◎△△ | |||||
| 巴郎杜鹃 R. balangense | 卧龙 Wolong | 2,700 | ○◎◎△ | |||||
| 基毛杜鹃 R. rigidum | 木里 Muli | 3,200 | ○◎◎△△ | |||||
| 紫花杜鹃 R. amesiae | 海螺沟 Hailuogou | 3,500 | ○◎◎△ | |||||
| 张口杜鹃 R. augustinii subsp. chasmanthum | 片马 Pianma | 3,100 | ○◎◎◎△△ | |||||
| L∈F: 叶期包花期型 Leafing covers flowering , | 喇叭杜鹃 R. discolor | 龙池 Longchi | 1,600 | △◎◎◎△ | ||||
| 绒毛杜鹃 R. pachytrichum | 海螺沟 Hailuogou | 3,200 | △◎◎◎△ | |||||
| 马缨杜鹃 R. delavayi | 大方 Dafang | 1,700 | △△△◎◎◎△ | |||||
| 皱皮杜鹃 R. wiltonii | 峨眉山 Emeishan | 2,400 | △◎◎◎△ | |||||
| 毛肋杜鹃 R. augustinii | 龙池 Longchi | 2,000 | △△◎◎◎△ | |||||
| 云南杜鹃 R. yunnanense | 玉龙雪山 Yulongxueshan | 3,600 | △◎◎◎△ | |||||
| 硬毛杜鹃 R. hirtipes | 多雄拉 Duoxiongla | 3,500 | △◎◎◎△△△△△△△△△△△△△△ | |||||
| 亮鳞杜鹃 R. heliolepis | 泸定 Luding | 3,100 | △△△◎◎◎△ | |||||
| L+F: 先叶后花、部分重叠型 Leafing ahead and covering part time of flowering | 大白杜鹃 R. decorum | 泸定 Luding | 1,800 | △◎◎○○ | ||||
| 硬叶杜鹃 R. tatsienense | 片马 Pianma | 3,200 | △△◎◎○ | |||||
| 长毛杜鹃 R. trichanthum | 川西 Chuanxi | 2,200 | △△◎○○ | |||||
| 问客杜鹃 R. ambiguum | 龙池 Longchi | 2,400 | △◎◎◎○ | |||||
| 山育杜鹃 R. oreotrephes | 泸定 Luding | 3,000 | △△◎◎○○ | |||||
| 三花杜鹃 R. triflorum | 多雄拉 Duoxiongla | 3,000 | △△△◎○○ | |||||
| 映山红 R. simsii | 大方 Dafang | 1,700 | △△△◎◎○ | |||||
| 长鳞杜鹃 R. longesquamatum | 龙池 Longchi | 2,700 | △△◎○○ | |||||
| L→F: 先叶后花型Leafing before flowering | 云雾杜鹃 R. chamaethomsonii | 多雄拉 Duoxiongla | 4,000 | △△△ ○○○ | ||||
| 白面杜鹃 R. zaleucum | 玉龙雪山 Yulongxueshan | 3,400 | △△△○○○ | |||||
| 亮毛杜鹃 R. microphyton | 无量山 Wuliangshan | 2,700 | △△△△○○○ | |||||
| 羊踯躅 R. moll | 庐山 Lushan | 1,300 | △△△△○○○○ | |||||
| [1] | Billings WD (1974) Adaptations and origins of alpine plants.Arctic and Alpine Research, 6, 129-142. |
| [2] | Billings WD, Mooney HA (1968) The ecology of arctic and alpine plant.Biological Reviews, 43, 481-529. |
| [3] | Brody AK (1997) Effects of pollinators, herbivores, and seed predators on flowering phenology.Ecology, 78, 1624-1631. |
| [4] | Chamberlain DF, Hyam R, Argent G (1996) The Genus Rhododendron: Its Classification and Synonymy. Royal Botanic Garden Edinburgh, Edinburgh. |
| [5] | Chen CD (陈昌笃), Zhuang P (庄平), Hu JC (胡锦矗) (2000) Biodiversity Research and Conservation in Dujiangyan,China (都江堰生物多样性研究与保护), pp. 11-27. Sichuan Science and Technology Press, Chengdu. (in Chinese) |
| [6] | Christian K (translated by Wu N (吴宁), Luo P (罗朋)) (2009) Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems (高山植物功能生态学), pp. 7-36, 87-96, 190-194, 223-236. Science Press, Beijing. (in Chinese) |
| [7] | Cullen J, Chamberlain DF (1978) A preliminary synopsis of the genus Rhododendron.Notes from the Royal Botanic Garden Edinburgh, 36, 105-126. |
| [8] | Dahl E (1986) Zonation in arctic and alpine tundra and fell-field ecobiomes. In: Ecosystem Theory Application (ed. Polunin N), pp. 35-62. Wiley, London. |
| [9] | Ding BY (丁炳扬), Jin XF (金孝锋) (2009) Taxonomic Study on Rhododendron Subgen. Tsutsusi Sleumer (Ericaceae) (杜鹃花属映山红亚属的分类研究), pp. 246-256. Science Press, Beijing. (in Chinese) |
| [10] | Escaravage N, Wagner J (2004) Pollination effectiveness and pollen dispersal in a Rhododendron ferrugineum (Ericaceae) population.Plant Biology, 6, 606-615. |
| [11] | Fang MY, Fang RZ (2005) Flora of China, Vol. 14. pp. 255-455. Science Press, Beijing. |
| [12] | Fang RZ (方瑞征), Min TL (闵天禄) (1981) The influence of uplift of Himalayas on the floristic formation of genus Rhododendron.Acta Botanica Yunnanica(云南植物研究), 3, 147-157. (in Chinese with English abstract) |
| [13] | Fang RZ (方瑞征), Min TL (闵天禄) (1995) The floristic study on the genus Rhododendron.Acta Botanica Yunnanica(云南植物研究), 17, 359-379. (in Chinese with English abstract) |
| [14] | Fennera M (1998) The phenology of growth and reproduction in plants. Perspectives in Plant Ecology,Evolution and Systematics, 1, 78-91. |
| [15] | Forrest J, Abraham JM (2010) Toward a synthetic understanding of the role of phenology in ecology and evolution.Philosophical Transactions of the Royal Society B: Biological Sciences, 365, 3101-3112. |
| [16] | Gaku K (1993) Relationships between flowering time and fruit set of the entomophilous alpine shrub, Rhododendron aureum (Ericaceae), inhabiting snow patches.American Journal of Botany, 80, 1300-1304. |
| [17] | Gaku K, Akira SH, Yuka K (2011) Pollination efficiency of bumblebee queens and workers in the alpine shrub Rhododendron aureum.International Journal of Plant Sciences, 172, 70-77. |
| [18] | Ge QS (葛全胜), Dai JH (戴君虎), Zheng JY (郑景云) (2010) The progress of phenology studies and challenges to modern phenology research in China.Disciplinary Development(科学发展), 25, 310-316. (in Chinese with English abstract) |
| [19] | Gibbs D, Chamberlain D, Argent G (2011) The Red List of Rhododendrons, pp. 6-8Botanic Gardens Conservation International, Richmond, UK. |
| [20] | Hideyuki D, Mayumi T, Izumi K (2010) Genetic diversity increases regional variation in phenological dates in response to climate change.Global Change Biology, 16, 373-379. |
| [21] | Hirao AS, Kameyama Y, Ohara M, Isagi Y, Kudo G (2006) Seasonal changes in pollinator activities influence pollen dispersal and seed production of the alpine shrub Rhododendron aureum (Ericaceae).Molecular Ecology, 15, 1165-1173. |
| [22] | Hu WG (胡文光) (1990) A study on the genus Rhododendron L. Subsection Fortunea Sleumer in China.Acta Botanica Yunnanica(云南植物研究), 12, 367-374. (in Chinese with English abstract) |
| [23] | Kudo G, Ida TY, Tani T (2008) Linkages between phenology, pollination, photosynthesis, and reproduction in deciduous forest understory plants.Ecology, 89, 321-331. |
| [24] | Li YN (李亚男), Yang DM (杨冬梅), Sun SC (孙书存), Gao XM (高贤明) (2008) Effects of twig size on biomass allocation within twigs and on Lamina area supporting efficiency in Rhododendron: allometric scaling analyses.Journal of Plant Ecology(植物生态学报), 32, 1175-1183. (in Chinese with English abstract) |
| [25] | Makrodimos N, Blionis GJ, Krigas N, Vokou D (2007) Flower morphology, phenology and visitor patterns in an alpine community on Mt. Olympos, Greece. Flora, 203, 449-468. |
| [26] | Malciūtė A, Naujalis JR, Šaulienė I (2010) The seasonal development characteristic of different rhododendrons taxa and cultivars in Northern Lithuania. 1. Leafing peculiarities.Agriculture, 97, 107-114. |
| [27] | Miller-Rushing AJ, Inouye DW (2009) Variation in the impact of climate change on flowering phenology and abundance: an examination of two pairs of closely related wildflower species.American Journal of Botany, 96, 1821-1829. |
| [28] | Min TL (闵天禄), Fang RZ (方瑞征) (1979) On the origin and geographic distribution of genus Rhododendron L.Acta Botanica Yunnanica(云南植物研究), 1, 121-127. (in Chinese with English abstract) |
| [29] | Min TL (闵天禄), Fang RZ (方瑞征) (1990) The phylogeny and evolution of genus Rhododendron.Acta Botanica Yunnanica(云南植物研究), 12, 353-365. (in Chinese with English abstract) |
| [30] | Molau U (1993) Relationships between flowering phenology and life history strategies in tundra plants.Arctic and Alpine Research, 25, 391-402. |
| [31] | Nilsen ET (2003) Unique anatomical traits in leaves of Rhododendron section Vireya: a discussion of functional significance. In: Rhododendrons in Horticulture and Science (eds Argent G, McFarlane M), pp. 20-36. Edinburg, Scotland. |
| [32] | Sailesh R, Eike L, Krishna KS, Kaiyun G, Xu JC (2013) Flowering phenology of tree rhododendron along an elevation gradient in two sites in the Eastern Himalayas.International Journal of Biometeorology, 57, 225-240. |
| [33] | Sun H (孙航) (2002) Tethys retreat and Himalayas- Hengduanshan Mountains uplift and their significance on the origin and development of the Sino-Himalayan elements and alpine flora.Acta Botanica Yunnanica(云南植物研究), 24, 273-288. (in Chinese with English abstract) |
| [34] | Wan WM (宛渭敏), Liu XZ (刘秀珍) (1979) Observation Method for China’s Phenology (中国物候观测方法), pp. 1-40. Science Press, Beijing. (in Chinese) |
| [35] | Wu ZY (吴征镒) (1991) The areal-types of Chinese genera of seed plant.Acta Botanica Yunnanica(云南植物研究), 13(Suppl. IV), 1-139. (in Chinese with English abstract) |
| [36] | Wu ZY (吴征镒), Sun H (孙航), Zhou ZK (周浙昆), Li DZ (李德铢), Peng H (彭华) (2011) Floristics of Seed Plants of China (中国种子植物区系地理), pp. 52-120. Science Press, Beijing. (in Chinese) |
| [37] | Xiong ZX (熊子仙), Du Q (杜青), Wang QD (王启德) (2000) Taxon and anatomy leaves in Rhododendron from China.Guihaia(广西植物), 20, 335-338. (in Chinese with English abstract) |
| [38] | Ying TS (应俊生), Chen ML (陈梦玲) (2011) Plant Geography of China (中国植物地理), pp. 68-79. Shanghai Science and Technology Press, Shanghai. (in Chinese) |
| [39] | Zhuang P (庄平) (2012) Discussion on the Rhododendron geographical distribution types and their cause of formation in China.Guihaia(广西植物), 32, 150-156. (in Chinese with English abstract) |
| [40] | Zhuang P (庄平), Gao XM (高贤明) (2002) The concept of “The West China Rainy Zone” and its significance to the biodiversity conservation in China.Biodiversity Science(生物多样性), 10, 339-344. (in Chinese with English abstract) |
| [41] | Zhuang P (庄平), Zheng YR (郑元润), Shao HM (邵慧敏), Wang F (王飞) (2012) An assessment on the adaptability of Rhododendron plants under ex situ conservation.Biodiversity Science(生物多样性), 20, 665-675. (in Chinese with English abstract) |
| [42] | Zhuang P (庄平), Wang F (王飞), Shao HM (邵慧敏) (2013) Comparative study on Rhododendron and its distribution in W-Sichuan and SE-Tibet.Guihaia(广西植物), 33, 791-797. (in Chinese with English abstract) |
| [1] | 何花, 谭敦炎, 杨晓琛. 被子植物隐性雌雄异株性系统的多样性、系统演化及进化意义[J]. 生物多样性, 2024, 32(6): 24149-. |
| [2] | 葛美玲, 徐勤增, 范士亮, 王宗兴, 张学雷. 中国近海多毛纲底栖类群目与科水平的分类[J]. 生物多样性, 2018, 26(9): 998-1003. |
| [3] | 孙军, 薛冰. 全球气候变化下的海洋浮游植物多样性[J]. 生物多样性, 2016, 24(7): 739-747. |
| [4] | . 温带森林展叶物候学①[J]. 生物多样性, 2013, 21(1): 111-116. |
| [5] | 郑钰, 高博, 孙立夫, 邴艳红, 裴克全. 银叶杜鹃和繁花杜鹃根部真菌的多样性[J]. 生物多样性, 2010, 18(1): 76-82. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
备案号:京ICP备16067583号-7
Copyright © 2026 版权所有 《生物多样性》编辑部
地址: 北京香山南辛村20号, 邮编:100093
电话: 010-62836137, 62836665 E-mail: biodiversity@ibcas.ac.cn
![]()